
ABSTRACT
Evidence suggests that activation of autophagy in neoplastic cells potentiates antitumor immunity through cross-presentation of tumor-associated antigens to T cells and release of immune mediators. The SQSTM1 (sequestosome 1, p62) protein is degraded by activated autophagy, and might enhance immune response to tumor cells. We hypothesized that tumor SQSTM1 expression level might be inversely associated with T-cell densities in colorectal carcinoma tissue. We evaluated tumor SQSTM1 expression by immunohistochemistry in 601 rectal and colon cancer cases within the Nurses' Health Study and Health Professionals Follow-up Study. Ordinal logistic regression analyses were conducted to assess the association of tumor SQSTM1 expression with CD3+, CD8+, CD45RO (PTPRC)+, or FOXP3+ cell density in tumor tissue, controlling for potential confounders, including tumor status of microsatellite instability, CpG island methylator phenotype, long interspersed nucleotide element-1 methylation level, and KRAS, BRAF, and PIK3CA mutations. Tumor SQSTM1 expression level was inversely associated with FOXP3+ cell density (ptrend = 0.006), but not with CD3+, CD8+, or CD45RO+ cell density (with the adjusted α level of 0.01 for multiple hypothesis testing). For a unit increase in quartile categories of FOXP3+ cell density, multivariable odds ratios were 0.66 [95% confidence interval (CI), 0.45–0.98] for intermediate-level SQSTM1 expression, and 0.55 (95% CI, 0.36–0.83) for high-level SQSTM1 expression, compared with low-level SQSTM1 expression. Tumor SQSTM1 expression is inversely associated with FOXP3+ cell density in colorectal cancer tissue, suggesting a possible role of SQSTM1-expressing carcinoma cells on regulatory T cells in the tumor microenvironment.
Abbreviations
| ATP | = | adenosine triphosphate |
| CI | = | confidence interval |
| CIMP | = | CpG island methylator phenotype |
| FFPE | = | formalin-fixed paraffin-embedded |
| LINE-1 | = | long interspersed nucleotide element-1 |
| MSI | = | microsatellite instability |
| MSS | = | microsatellite stable |
| OR | = | odds ratio |
| SD | = | standard deviation |
Introduction
Accumulating evidence attests a key role of T-cell-mediated adaptive immunity in inhibiting tumor evolution, and immunotherapy has emerged as a promising strategy to treat various cancers.Citation1-5 Autophagy is a homeostatic cellular recycling mechanism responsible for degrading cellular organelles and proteins. Autophagic activity in tumor cells may enhance extracellular release of immune mediators and cross-presentation of tumor-associated antigens to T cells, thereby potentiating immune response to tumor cells.Citation6-19 Emerging evidence attests to a key role of tumor autophagic activity in modulating functions of T cells such as CD8+ cytotoxic T cells and FOXP3+ regulatory T cells.Citation17,20-22 The SQSTM1 (sequestosome 1, p62) protein is a ubiquitin-binding scaffold molecule that plays a key role in autophagic degradation of ubiquitinated proteins,Citation23-29 and the degradation of SQSTM1 with tumor-related antigen may promote T-cell-mediated immunity.Citation30-32
Colorectal cancer represents a heterogeneous group of neoplasms resulting from genomic and epigenomic alterations, which influence and are influenced by tumor–host interactions.Citation33-36 A strong immune response to colorectal cancer manifested as high density of CD3+, CD8+, or CD45RO (PTPRC)+ T cells has been consistently associated with better clinical outcome.Citation37-43 An enhanced infiltration of T cells in colorectal cancer tissue has been associated with specific tumor molecular status, including high-level microsatellite instability (MSI-high).Citation41-45 Studies have shown that SQSTM1 is overexpressed in colorectal cancer,Citation46,47 but significance of SQSTM1 expression in colorectal cancer needs to be investigated. We hypothesized that low-level tumor SQSTM1 expression (indicating high autophagic activity) might be associated with high T-cell densities in colorectal cancer tissue. Because the complexity of tumor–host immune interactions in human cancers cannot be exactly recapitulated by any in vitro or non-human models, analyses of tumor characteristics and immune cells in human cancer tissue are valuable.
To test our hypothesis, we examined tumor SQSTM1 expression in relation to CD3+, CD8+, CD45RO+, or FOXP3+ cell densities in cancer tissue of more than 600 human colorectal cancer cases within the two US-nationwide prospective cohort studies. A better understanding of the relationship between autophagy and immune cells in the tumor microenvironment may open new opportunities to target autophagy and immunity for colorectal cancer prevention and therapy.
Results
Tumor SQSTM1 (p62) expression in colorectal cancer
We examined immunohistochemical expression levels of the SQSTM1 protein in 601 cases of colorectal carcinoma within the two US-nationwide prospective cohort studies. Among the 601 colorectal cancer cases, 131 (22%), 271 (45%), and 199 (33%) tumors showed low-level, intermediate-level, and high-level SQSTM1 expression, respectively.
Clinical, pathological, and molecular characteristics according to the tumor SQSTM1 expression levels in colorectal cancer are summarized in . Tumor SQSTM1 expression level was not significantly associated with any of the characteristics examined (p > 0.02; with the adjusted α level of 0.003 for multiple hypothesis testing) ().
Table 1. Clinical, pathological, and molecular features of colorectal cancers according to tumor SQSTM1 (p62) expression level.
Association of tumor SQSTM1 expression with T-cell density in colorectal cancer
shows the distribution of colorectal carcinoma cases according to the tumor SQSTM1 expression level and T-cell densities. Tumor SQSTM1 expression level was inversely correlated with FOXP3+ cell density (p = 0.001, by Spearman correlation test) with the adjusted α level of 0.01. In our primary hypothesis testing, we conducted ordinal logistic regression analyses to assess the associations of the tumor SQSTM1 expression level (an ordinal predictor variable) with the density of CD3+, CD8+, CD45RO+, or FOXP3+ cells (an ordinal quartile outcome variable) in colorectal cancer tissue ( and Table S1). Tumor SQSTM1 expression level was inversely associated with FOXP3+ cell density in ordinal logistic regression analyses (all ptrend ≤ 0.006; with the adjusted α level of 0.01). For a unit increase in quartile categories of FOXP3+ cell density, the multivariable ORs were 0.66 [95% confidence interval (CI), 0.45–0.98] for cases with intermediate-level SQSTM1 expression and 0.55 (95% CI, 0.36–0.83) for those with high-level SQSTM1 expression, compared with those with low-level SQSTM1 expression. The tumor SQSTM1 expression level was not significantly associated with CD3+, CD8+, or CD45RO+ cell density (all ptrend > 0.05; with the adjusted α level of 0.01).
Table 2. Distribution of colorectal cancer cases according to tumor SQSTM1 expression level and the density of T cells.
Table 3. Ordinal logistic regression analysis to assess the association of tumor SQSTM1 expression level (predictor) with the density of T cells (outcome).
Discussion
Using the database of the 601 colorectal cancer cases in the two US-nationwide prospective cohort studies, we found that higher tumor SQSTM1 expression was associated with lower density of FOXP3+ cells in human colorectal cancer tissue. The association persisted after controlling for potential confounders, including the tumor statuses of MSI, CIMP and LINE-1 methylation level, which have been correlated with the abundance of tumor infiltrating T cells in colorectal cancer.Citation41-45 Although a replication in independent data sets is needed, our human population-based data suggest a possible role of autophagic activity of tumor cells in regulating host immunity in colorectal cancer microenvironment.
Colorectal cancer development is not only driven by genomic and epigenomic alterations of tumor cells but also influenced by tumor–host interactions.Citation48-53 The importance of analyses of human tumor characteristics and host immunity has been increasing.Citation37,54 In fact, higher densities of CD3+, CD8+, and CD45RO+ cells in colorectal cancer tissue have been associated with better prognosis,Citation38-43,55 suggesting antitumor effects of these T cells in the tumor microenvironment. Therefore, there is a great need to identify potential molecular targets that can influence T-cell-mediated immune response to tumor.
FOXP3+ regulatory T cells have been considered as an immunosuppressive subset of T lymphocytes, and are functionally and phenotypically diverse with various functional profiles.Citation56 Accumulating evidence indicates that function of FOXP3+ regulatory T cells can be tailored for differing immune milieu and contexts, and that their roles for cancer progression (tumor-promoting or tumor-suppressive roles) appear to depend on tumor site and progression stage, probably reflecting alterations of the tumor microenvironment.Citation39,56 Although functional roles of FOXP3+ cells in various types of cancers remain unclear, high density of FOXP3+ cells has been generally associated with favorable outcome in colorectal cancer patients.Citation39-43,55 Particularly, abundant infiltration of T cells with low-intensity FOXP3 expression may be associated with better outcome.Citation57 Ladoire et al. have proposed that FOXP3+ regulatory cells have a crucial role in inhibiting tumor-promoting inflammatory responses to gut microbiota, which may explain favorable prognosis associated with abundant FOXP3+ cells in colorectal cancer.Citation58 Taken together, it seems to be plausible that FOXP3+ regulatory T cells may have a role in suppressing colorectal tumor progression through regulating tumor-promoting inflammation.
Autophagy is an evolutionarily conserved catabolic process by which cellular components are sequestered into a double-membrane vesicle (autophagosome) and delivered to the lysosome for terminal degradation and recycling.Citation16 Accumulating evidence suggests that autophagy plays a critical role in the regulation of immune response,Citation19,59 in particular, antitumor immunity that may influence response to immunotherapies.Citation16 Autophagy in neoplastic cells appears to increase the emission of potent chemotactic factors including adenosine triphosphate (ATP) and lysophosphatidylcholine, which in turn attracts immune cells to the tumor bed.Citation11-15 Emerging evidence provides that autophagic activity in tumor cells is essential for promoting cross-presentation of tumor-associated antigens to T cells, indicating a positive link between autophagy and adaptive immunity.Citation8-11 On the other hand, autophagy may has an immunosuppressive role,Citation60 while keeping inverse balance with FOXP3+ cells. These lines of evidence together with our current findings suggest that tumors may choose one or the other for immune evasion.
Accumulating evidence attests to a key role of tumor autophagic activity on not only CD8+ cytotoxic T cells but also FOXP3+ regulatory T cells in the tumor microenvironment although detailed mechanisms have not been discovered.Citation17,20-22 In this study, tumor SQSTM1 expression level was inversely associated with FOXP3+ cell density in tumor tissue, but not density of CD3+, CD8+, or CD45RO+ cells. Potential reasons for the difference may include difficulty in accurately measuring cellular autophagic activity, which might have caused false negative findings.
In our secondary analyses, we observed trends toward positive associations of tumor SQSTM1 expression with cecal tumor location, wild-type KRAS, and a higher level Fusobacterium nucleatum DNA in tumor tissue. One previous studyCitation46 has reported that tumor SQSTM1 expression is not associated with tumor location, but the sample size was much smaller (n = 178) than our current study. In an in vitro experimental study,Citation61 KRAS mutation in colorectal cancer appeared to attenuate tumor SQSTM1 expression level, and we observed the similar trend. In another in vitro experimental study,Citation62 F. nucleatum appeared to induce impairment of autophagic activity of host cells; however, our current data do not support such negative effects of F. nucleatum on autophagic activity of tumor cells. Further studies are needed to determine the associations of tumor SQSTM1 expression with clinical, pathological and molecular characteristics of colorectal cancer.
One limitation of our current study is its cross-sectional nature. Hence, we cannot exclude the possibility of reverse causation. It is possible that FOXP3+ cells might alter the tumor autophagy status. However, our specific hypothesis was based on several lines of experimental evidence indicating that autophagic activity in tumor cells promotes antitumor immune response.Citation8-16 Another limitation is that our study used SQSTM1 (but no other biomarkers) to quantify autophagic activity. Future studies should evaluate other autophagic markers including MAP1LC3 (LC3). Strengths of this study include the use of our molecular pathological epidemiologyCitation42, 63 database of more than 600 colorectal cancer cases in the two US-nationwide prospective cohort studies, which integrate epidemiologic exposures, clinicopathological features, tumor molecular features, and immune reaction status in colorectal cancer tissue. This population-based colorectal cancer database enabled us to rigorously examine the association of tumor SQSTM1 expression level with the T-cell density, controlling for potential confounders. In addition, our colorectal cancer specimens were derived from a large number of hospitals in diverse settings across the United States (but not based on a limited number of hospitals), which increases the generalizability of our findings. Last, we also used robust laboratory assays including tissue image analysis that could objectively quantify specific T-cell populations in tumor tissue.
In summary, tumor SQSTM1 expression level is inversely associated with FOXP3+ cell density in colorectal cancer tissue. Our population-based data suggest a possible effect of tumor SQSTM1 expression and autophagic activity on regulatory T cells in colorectal cancer microenvironment, and can promote further translational research on the associations of autophagy with host immunity in colorectal cancer.
Patients and methods
Study population
We used two independent US-nationwide prospective cohort studies: the Nurses' Health Study (121,701 women followed since 1976) and the Health Professionals Follow-up Study (51,529 men followed since 1986).Citation64-66 Every 2 y, study staff had sent follow-up questionnaires to the participants, to update information on diet and lifestyle factors and to identify newly diagnosed cancer and other diseases. In addition, we identified deaths of cohort participants in the National Death Index, to find fatal colorectal cancer cases that had not been reported. Study physicians reviewed medical records to gain information on tumor location and disease stage, and determined cause of deaths for deceased individuals. Formalin-fixed paraffin-embedded (FFPE) tissue blocks were collected from hospitals across the United States, where participants with colorectal cancer had undergone tumor resection. We included both colon and rectal carcinoma cases, considering the colorectal continuum model.Citation67 The study pathologist (S.O.) blinded to other data conducted centralized pathology review of all colorectal carcinoma cases, and recorded pathological features including tumor differentiation, and four patterns of histological lymphocytic reaction [Crohn's-like lymphoid reaction, peritumoral lymphocytic reaction, intratumoral periglandular reaction, and tumor-infiltrating lymphocytes (TILs)].Citation44,68 Tumor differentiation was categorized as well to moderate (> 50% glandular area) or poor (≤ 50% glandular area). Based on availability of data on tumor SQSTM1 expression and T-cell densities, a total of 601 colorectal cancer cases diagnosed up to 2008 were included in this study. Written informed consent was obtained from all study participants. Tissue collection and analyses were approved by the institutional review boards at the Harvard T.H. Chan School of Public Health and the Brigham and Women's Hospital (Boston, MA, USA).
Immunohistochemistry for CD3+, CD8+, CD45RO, FOXP3, and SQSMT1
We constructed tissue microarray from colorectal cancer blocks,Citation69 and conducted immunohistochemistry. Immunohistochemical analyses were performed for CD3+, CD8+, CD45RO (one isoform of the PTPRC protein), and FOXP3, as described previously.Citation43 We measured the densities of CD3+, CD8+, CD45RO+, and FOXP3+ cells in tumor tissue by using an automated scanning microscope and the Ariol image analysis system (Genetix). We evaluated up to four tissue microarray cores from each tumor, and calculated the average density (cells/mm2) of each T-cell population.Citation43
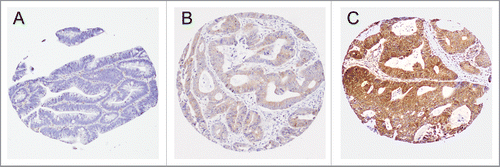
We performed immunohistochemistry for SQSTM1 (p62) as an autophagy-related marker. Since SQSTM1 is degraded by autophagy, autophagic activity has been inversely associated with SQSTM1 expression.Citation6,23-25,70 For immunohistochemistry, deparaffinized tissue sections were heated in a microwave using a pressure cooker for 17 min in Antigen Retrieval Citra Solution, pH 6 (BioGenex Laboratories). Tissue sections were incubated with a dual endogenous enzyme block (Dako) for 30 min and then serum-free protein block (Dako) for 10 min. Slides were incubated for 16 h at 4°C with a primary antibody against SQSTM1 (mouse monoclonal antibody, clone 2C11, Abnova; dilution, 1:1,500). Then, the EnVision HRP-labeled polymer (Dako) was applied to the sections for 30 min, followed by visualization with 3,3-diaminobenzidine and counterstaining with hematoxylin. Sections processed with the replacement of the primary antibody with Tris-buffered saline were used as negative controls. The cytoplasmic expression level (intensity) of SQSTM1 was recorded as low, intermediate, or high (). Tumor SQSTM1 expression was interpreted by a single pathologist (Y.M.) unaware of other data. A sample of 143 tumors was examined by a second pathologist (A.dS.). The weighted κ value for agreement between the two pathologists for SQSTM1 was 0.63, indicating reasonably good interobserver agreement (p < 0.0001 by Spearman's correlation test).
Figure 1. Tumor SQSTM1 (p62) expression in colorectal cancer. Tumor SQSTM1 expression was scored as low (A), intermediate (B), or high (C), according to cytoplasmic expression level of SQSTM1 in tumor cells.
Analyses of MSI, DNA methylation, and KRAS, BRAF, and PIK3CA mutations
DNA was extracted from archival colorectal cancer tissue blocks. MSI status was analyzed with the use of 10 microsatellite markers (BAT25, BAT26, BAT40, D2S123, D5S346, D17S250, D18S55, D18S56, D18S67, and D18S487), as described previously.Citation66,71 We defined MSI-high as the presence of instability in ≥ 30% of the markers, and MSI-low/microsatellite stability (MSS) as instability in <30% of the markers.Citation66,71 Methylation analyses of long interspersed nucleotide element-1 (LINE-1) and eight promoters specific to CpG island methylator phenotype (CIMP) (CACNA1G, CDKN2A, CRABP1, IGF2, MLH1, NEUROG1, RUNX3, and SOCS1) were performed.Citation66,72 CIMP-high was defined as ≥ 6/8 methylated promoters, while CIMP-low/negative as 0/8 to 5/8 methylated promoters, as described previously.Citation66,72 PCR reaction and pyrosequencing were performed for KRAS (codons 12, 13, 61, and 146),Citation73 BRAF (codon 600),Citation71 and PIK3CA (exons 9 and 20).Citation66,74
Analysis of the amount of F. nucleatum DNA
We extracted DNA from colorectal cancer FFPE tissue sections, and performed a quantitative PCR assay to measure the amount of tissue F. nucleatumDNA.Citation75 We categorized colorectal carcinoma cases with detectable F. nucleatum DNA as low or high in relation to the median of F. nucleatum DNA amounts among F. nucleatum detectable cases.Citation75,76
Statistical analysis
All statistical analyses were conducted using SAS (version 9.4, SAS Institute, Cary, NC, USA), and all p values were two-sided. Our primary hypothesis testing was assessment of the association of the tumor SQSTM1 expression level (an ordinal predictor variable) with the density of CD3+, CD8+, CD45RO+, or FOXP3+ cells in colorectal cancer tissue (an ordinal quartile outcome variable). Because we tested four primary outcome variables (CD3+ cells, CD8+ cells, CD45RO+ cells, and FOXP3+ cells), we adjusted a two-sided significance level to 0.01 ( = 0.05/4) based on the Bonferroni correction. All other analyses, including evaluation of individual odds ratio (OR) estimates, were secondary analyses.
To control for potential confounding, we performed multivariable ordinal logistic regression analysis where each T-cell density variable (CD3+, CD8+, CD45RO, and FOXP3) was used as an ordinal quartile outcome variable, and SQSTM1 expression level as the ordinal predictor variable of our primary interest. In the regression model, we initially included age (continuous), sex (female vs. male), year of diagnosis (continuous), family history of colorectal cancer in a first-degree relative (present vs. absent vs. missing), tumor location (proximal colon vs. distal colon vs. rectum vs. missing), MSI status (MSI-high vs. MSI-low/MSS vs. missing), CIMP status (high vs. low/negative vs. missing), KRAS mutation (mutant vs. wild-type vs. missing), BRAF mutation (mutant vs. wild-type vs. missing), PIK3CA mutation (mutant vs. wild-type vs. missing), and LINE-1 methylation level (continuous, with a missing indicator variable). Then, we performed a backward elimination with a threshold of p = 0.05 to select variables for the final model. In the final multivariable ordinal logistic regression model, for cases with missing information in any of the selected categorical variables, we included those cases in the majority category of a given covariate to limit the degrees of freedom and avoid overfitting of the model. We assessed the proportional odds assumption in the ordinal logistic regression model, which was generally satisfied (p > 0.05).
To assess the associations of SQSTM1 expression level with other categorical variables, the chi-square test was performed. To compare mean age and mean LINE-1 methylation levels, an analysis of variance assuming equal variances was performed. All of the cross-sectional univariable analyses for clinical, pathological, and molecular associations (with variables listed in ) were secondary exploratory analyses, and we adjusted two-sided sinificance level to 0.003 (= 0.05/18) by the Bonferroni correction for multiple hypothesis testing.
Disclosure of potential conflict of interest
A.T.C. previously served as a consultant for Bayer Healthcare, Millennium Pharmaceuticals, Pozen, Inc., and Pfizer, Inc. This study was not funded by Bayer Healthcare, Millennium Pharmaceuticals, Pozen, Inc., or Pfizer, Inc. The other authors declare that they have no conflicts of interest.
KONI_A_1284720_Supplementary_table_1.docx
Download MS Word (31.7 KB)Acknowledgments
We would like to thank the participants and staff of the Nurses' Health Study and the Health Professionals Follow-up Study for their valuable contributions as well as the following state cancer registries for their help: AL, AZ, AR, CA, CO, CT, DE, FL, GA, ID, IL, IN, IA, KY, LA, ME, MD, MA, MI, NE, NH, NJ, NY, NC, ND, OH, OK, OR, PA, RI, SC, TN, TX, VA, WA, WY. The authors assume full responsibility for analyses and interpretation of these data.
Funding
This work was supported by U.S. National Institutes of Health (NIH) grants [P01 CA87969 to M.J. Stampfer; UM1 CA186107 to M.J. Stampfer; P01 CA55075 to W.C. Willett; UM1 CA167552 to W.C. Willett; P50 CA127003 to C.S.F.; R01 CA137178 to A.T.C.; R01 CA151993 to S.O.; R35 CA197735 to S.O.; and K07 CA190673 to R.N.]; Nodal Award (to S.O.) from the Dana-Farber Harvard Cancer Center; and by grants from the Project P Fund, the Friends of the Dana-Farber Cancer Institute, Bennett Family Fund, and the Entertainment Industry Foundation through National Colorectal Cancer Research Alliance. J.Y is supported by National Natural Science Foundation of China (81200612); Tianjin City High School Science & Technology Fund Planning Project (20102217). K.K. is supported by a grant from Program for Advancing Strategic International Networks to Accelerate the Circulation of Talented Researchers from Japanese Society for the Promotion of Science. Y.M. is supported by a fellowship grant of the Keio Gijuku Fukuzawa Memorial Fund for the Advancement of Education and Research. T.H. was supported by a fellowship grant from the Uehara Memorial Foundation and by a grant from the Mochida Memorial Foundation for Medical and Pharmaceutical Research.
References
- Sharma P, Allison JP. The future of immune checkpoint therapy. Science 2015; 348:56-61; PMID:25838373; http://dx.doi.org/10.1126/science.aaa8172
- Le DT, Uram JN, Wang H, Bartlett BR, Kemberling H, Eyring AD, Skora AD, Luber BS, Azad NS, Laheru D, et al. PD-1 blockade in tumors with mismatch-repair deficiency. N Engl J Med 2015; 372:2509-20; PMID:26028255; http://dx.doi.org/10.1056/NEJMoa1500596
- Taube JM, Klein A, Brahmer JR, Xu H, Pan X, Kim JH, Chen L, Pardoll DM, Topalian SL, Anders RA. Association of PD-1, PD-1 ligands, and other features of the tumor immune microenvironment with response to anti-PD-1 therapy. Clin Cancer Res 2014; 20:5064-74; PMID:24714771; http://dx.doi.org/10.1158/1078-0432.CCR-13-3271
- Topalian SL, Drake CG, Pardoll DM. Immune checkpoint blockade: a common denominator approach to cancer therapy. Cancer Cell 2015; 27:450-61; PMID:25858804; http://dx.doi.org/10.1016/j.ccell.2015.03.001
- Postow MA, Callahan MK, Wolchok JD. Immune checkpoint blockade in cancer therapy. J Clin Oncol 2015; 33:1974-82; PMID:25605845; http://dx.doi.org/10.1200/JCO.2014.59.4358
- Kaur J, Debnath J. Autophagy at the crossroads of catabolism and anabolism. Nat Rev Mol Cell Biol 2015; 16:461-72; PMID:26177004; http://dx.doi.org/10.1038/nrm4024
- Umemura A, He F, Taniguchi K, Nakagawa H, Yamachika S, Font-Burgada J, Zhong Z, Subramaniam S, Raghunandan S, Duran A, et al. p62, Upregulated during preneoplasia, induces hepatocellular carcinogenesis by maintaining survival of stressed HCC-initiating cells. Cancer Cell 2016; 29:935-48; PMID:27211490; http://dx.doi.org/10.1016/j.ccell.2016.04.006
- Li Y, Wang LX, Yang G, Hao F, Urba WJ, Hu HM. Efficient cross-presentation depends on autophagy in tumor cells. Cancer Res 2008; 68:6889-95; PMID:18757401; http://dx.doi.org/10.1158/0008-5472.CAN-08-0161
- Li Y, Wang LX, Pang P, Cui Z, Aung S, Haley D, Fox BA, Urba WJ, Hu HM. Tumor-derived autophagosome vaccine: mechanism of cross-presentation and therapeutic efficacy. Clin Cancer Res 2011; 17:7047-57; PMID:22068657; http://dx.doi.org/10.1158/1078-0432.CCR-11-0951
- Li Y, Hahn T, Garrison K, Cui ZH, Thorburn A, Thorburn J, Hu HM, Akporiaye ET. The vitamin E analogue alpha-TEA stimulates tumor autophagy and enhances antigen cross-presentation. Cancer Res 2012; 72:3535-45; PMID:22745370; http://dx.doi.org/10.1158/0008-5472.CAN-11-3103
- Ma Y, Galluzzi L, Zitvogel L, Kroemer G. Autophagy and cellular immune responses. Immunity 2013; 39:211-27; PMID:23973220; http://dx.doi.org/10.1016/j.immuni.2013.07.017
- Elliott MR, Chekeni FB, Trampont PC, Lazarowski ER, Kadl A, Walk SF, Park D, Woodson RI, Ostankovich M, Sharma P, et al. Nucleotides released by apoptotic cells act as a find-me signal to promote phagocytic clearance. Nature 2009; 461:282-6; PMID:19741708; http://dx.doi.org/10.1038/nature08296
- Michaud M, Martins I, Sukkurwala AQ, Adjemian S, Ma Y, Pellegatti P, Shen S, Kepp O, Scoazec M, Mignot G, et al. Autophagy-dependent anticancer immune responses induced by chemotherapeutic agents in mice. Science 2011; 334:1573-7; PMID:22174255; http://dx.doi.org/10.1126/science.1208347
- Martins I, Michaud M, Sukkurwala AQ, Adjemian S, Ma Y, Shen S, Kepp O, Menger L, Vacchelli E, Galluzzi L, et al. Premortem autophagy determines the immunogenicity of chemotherapy-induced cancer cell death. Autophagy 2012; 8:413-5; PMID:22361584; http://dx.doi.org/10.4161/auto.19009
- Deretic V, Saitoh T, Akira S. Autophagy in infection, inflammation and immunity. Nat Rev Immunol 2013; 13:722-37; PMID:24064518; http://dx.doi.org/10.1038/nri3532
- Pan H, Chen L, Xu Y, Han W, Lou F, Fei W, Liu S, Jing Z, Sui X. Autophagy-associated immune responses and cancer immunotherapy. Oncotarget 2016; 7(16):21235-46; PMID:26788909; http://dx.doi.org/10.18632/oncotarget.6908
- Zhong Z, Sanchez-Lopez E, Karin M. Autophagy, inflammation, and immunity: a Troika governing cancer and its treatment. Cell 2016; 166:288-98; PMID:27419869; http://dx.doi.org/10.1016/j.cell.2016.05.051
- Pietrocola F, Pol J, Vacchelli E, Baracco EE, Levesque S, Castoldi F, Maiuri MC, Madeo F, Kroemer G. Autophagy induction for the treatment of cancer. Autophagy 2016; 12(10):1962-1964; PMID:27532519; http://dx.doi.org/10.1080/15548627.2016.1214778
- Cui J, Jin S, Wang RF. The BECN1-USP19 axis plays a role in the crosstalk between autophagy and antiviral immune responses. Autophagy 2016; 12(7):1-2; PMID:27096686; http://dx.doi.org/10.1080/15548627.2016.1173801
- Ladoire S, Enot D, Senovilla L, Chaix M, Zitvogel L, Kroemer G. Positive impact of autophagy in human breast cancer cells on local immunosurveillance. Oncoimmunology 2016; 5:e1174801; PMID:27471653; http://dx.doi.org/10.1080/2162402X.2016.1174801
- Ladoire S, Enot D, Senovilla L, Ghiringhelli F, Poirier-Colame V, Chaba K, Semeraro M, Chaix M, Penault-Llorca F, Arnould L, et al. The presence of LC3B puncta and HMGB1 expression in malignant cells correlate with the immune infiltrate in breast cancer. Autophagy 2016; 12:864-75; PMID:26979828; http://dx.doi.org/10.1080/15548627.2016.1154244
- Ladoire S, Senovilla L, Enot D, Ghiringhelli F, Poirier-Colame V, Chaba K, Erdag G, Schaefer JT, Deacon DH, Zitvogel L, et al. Biomarkers of immunogenic stress in metastases from melanoma patients: correlations with the immune infiltrate. Oncoimmunology 2016; 5:e1160193; PMID:27471635; http://dx.doi.org/10.1080/2162402X.2016.1160193
- Bjorkoy G, Lamark T, Pankiv S, Overvatn A, Brech A, Johansen T. Monitoring autophagic degradation of p62/SQSTM1. Methods Enzymol 2009; 452:181-97. PMID:19200883; http://dx.doi.org/10.1016/S0076-6879(08)03612-4
- Mathew R, Karp CM, Beaudoin B, Vuong N, Chen G, Chen HY, Bray K, Reddy A, Bhanot G, Gelinas C, et al. Autophagy suppresses tumorigenesis through elimination of p62. Cell 2009; 137:1062-75; PMID:19524509; http://dx.doi.org/10.1016/j.cell.2009.03.048
- Rogov V, Dotsch V, Johansen T, Kirkin V. Interactions between autophagy receptors and ubiquitin-like proteins form the molecular basis for selective autophagy. Mol Cell 2014; 53:167-78; PMID:24462201; http://dx.doi.org/10.1016/j.molcel.2013.12.014
- Bjorkoy G, Lamark T, Brech A, Outzen H, Perander M, Overvatn A, Stenmark H, Johansen T. p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. J Cell Biol 2005; 171:603-14; PMID:16286508; http://dx.doi.org/10.1083/jcb.200507002
- Pankiv S, Clausen TH, Lamark T, Brech A, Bruun JA, Outzen H, Øvervatn A, Bjørkøy G, Johansen T. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J Biol Chem 2007; 282:24131-45; PMID:17580304; http://dx.doi.org/10.1074/jbc.M702824200
- Katsuragi Y, Ichimura Y, Komatsu M. p62/SQSTM1 functions as a signaling hub and an autophagy adaptor. FEBS J 2015; 282:4672-8; PMID:26432171; http://dx.doi.org/10.1111/febs.13540
- Taniguchi K, Yamachika S, He F, Karin M. p62/SQSTM1—Dr. Jekyll and Mr. Hyde that prevents oxidative stress but promotes liver cancer. FEBS Lett 2016; 590(15):2375-97; PMID:27404485; http://dx.doi.org/10.1002/1873-3468.12301
- Kanayama M, Inoue M, Danzaki K, Hammer G, He YW, Shinohara ML. Autophagy enhances NFkappaB activity in specific tissue macrophages by sequestering A20 to boost antifungal immunity. Nat Commun 2015; 6:5779; PMID:25609235; http://dx.doi.org/10.1038/ncomms6779
- Lee Y, Sasai M, Ma JS, Sakaguchi N, Ohshima J, Bando H, Saitoh T, Akira S, Yamamoto M. p62 plays a specific role in interferon-gamma-induced presentation of a toxoplasma vacuolar antigen. Cell Rep 2015; 13:223-33; PMID:26440898; http://dx.doi.org/10.1016/j.celrep.2015.09.005
- Andersen AN, Landsverk OJ, Simonsen A, Bogen B, Corthay A, Oynebraten I. Coupling of HIV-1 antigen to the selective autophagy receptor SQSTM1/p62 promotes T-cell-mediated immunity. Front Immunol 2016; 7:167; PMID:27242780; http://dx.doi.org/10.3389/fimmu.2016.00167
- Kuipers EJ, Grady WM, Lieberman D, Seufferlein T, Sung JJ, Boelens PG, van de Velde CJ, Watanabe T. Colorectal cancer. Nat Rev Dis Primers 2015; 1:15065; PMID:27189416; http://dx.doi.org/10.1038/nrdp.2015.65
- Okugawa Y, Grady WM, Goel A. Epigenetic alterations in colorectal cancer: emerging biomarkers. Gastroenterology 2015; 149:1204-25.e12; PMID:26216839; http://dx.doi.org/10.1053/j.gastro.2015.07.011
- Di Caro G, Marchesi F, Laghi L, Grizzi F. Immune cells: plastic players along colorectal cancer progression. J Cell Mol Med 2013; 17:1088-95; PMID:24151976; http://dx.doi.org/10.1111/jcmm.12117
- Colussi D, Brandi G, Bazzoli F, Ricciardiello L. Molecular pathways involved in colorectal cancer: implications for disease behavior and prevention. Int J Mol Sci 2013; 14:16365-85; PMID:23965959; http://dx.doi.org/10.3390/ijms140816365
- Mlecnik B, Bindea G, Angell HK, Maby P, Angelova M, Tougeron D, Church SE, Lafontaine L, Fischer M, Fredriksen T, et al. Integrative analyses of colorectal cancer show immunoscore is a stronger predictor of patient survival than microsatellite instability. Immunity 2016; 44:698-711; PMID:26982367; http://dx.doi.org/10.1016/j.immuni.2016.02.025
- Galon J, Costes A, Sanchez-Cabo F, Kirilovsky A, Mlecnik B, Lagorce-Pages C, Tosolini M, Camus M, Berger A, Wind P, et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 2006; 313:1960-4; PMID:17008531; http://dx.doi.org/10.1126/science.1129139
- Shang B, Liu Y, Jiang SJ, Liu Y. Prognostic value of tumor-infiltrating FoxP3(+) regulatory T cells in cancers: a systematic review and meta-analysis. Sci Rep 2015; 5:15179; PMID:26462617; http://dx.doi.org/10.1038/srep15179
- Mei Z, Liu Y, Liu C, Cui A, Liang Z, Wang G, Peng H, Cui L, Li C. Tumour-infiltrating inflammation and prognosis in colorectal cancer: systematic review and meta-analysis. Br J Cancer 2014; 110:1595-605; PMID:24504370; http://dx.doi.org/10.1038/bjc.2014.46
- Palmqvist R, Wikberg M, Ling A, Edin S. The association of immune cell infiltration and prognosis in colorectal cancer. Curr Colorectal Cancer Rep 2013; 9:372-9; ; http://dx.doi.org/10.1007/s11888-013-0192-3
- Ogino S, Galon J, Fuchs CS, Dranoff G. Cancer immunology – analysis of host and tumor factors for personalized medicine. Nat Rev Clin Oncol 2011; 8:711-9; PMID:21826083; http://dx.doi.org/10.1038/nrclinonc.2011.122
- Nosho K, Baba Y, Tanaka N, Shima K, Hayashi M, Meyerhardt JA, Giovannucci E, Dranoff G, Fuchs CS, Ogino S. Tumour-infiltrating T-cell subsets, molecular changes in colorectal cancer, and prognosis: cohort study and literature review. J Pathol 2010; 222:350-66; PMID:20927778; http://dx.doi.org/10.1002/path.2774
- Ogino S, Nosho K, Irahara N, Meyerhardt JA, Baba Y, Shima K, Glickman JN, Ferrone CR, Mino-Kenudson M, Tanaka N, et al. Lymphocytic reaction to colorectal cancer is associated with longer survival, independent of lymph node count, microsatellite instability, and CpG island methylator phenotype. Clin Cancer Res 2009; 15:6412-20; PMID:19825961; http://dx.doi.org/10.1158/1078-0432.CCR-09-1438
- Llosa NJ, Cruise M, Tam A, Wicks EC, Hechenbleikner EM, Taube JM, Blosser RL, Fan H, Wang H, Luber BS, et al. The vigorous immune microenvironment of microsatellite instable colon cancer is balanced by multiple counter-inhibitory checkpoints. Cancer Discov 2015; 5:43-51; PMID:25358689; http://dx.doi.org/10.1158/2159-8290.CD-14-0863
- Park JM, Huang S, Wu TT, Foster NR, Sinicrope FA. Prognostic impact of Beclin 1, p62/sequestosome 1 and LC3 protein expression in colon carcinomas from patients receiving 5-fluorouracil as adjuvant chemotherapy. Cancer Biol Ther 2013; 14:100-7; PMID:23192274; http://dx.doi.org/10.4161/cbt.22954
- Ren F, Shu G, Liu G, Liu D, Zhou J, Yuan L, Zhou J. Knockdown of p62/sequestosome 1 attenuates autophagy and inhibits colorectal cancer cell growth. Mol Cell Biochem 2014; 385:95-102; PMID:24065390; http://dx.doi.org/10.1007/s11010-013-1818-0
- Kocarnik JM, Shiovitz S, Phipps AI. Molecular phenotypes of colorectal cancer and potential clinical applications. Gastroenterol Rep (Oxf) 2015; 3:269-76. PMID:26337942; http://dx.doi.org/10.1093/gastro/gov046
- Ogino S, Fuchs CS, Giovannucci E. How many molecular subtypes? Implications of the unique tumor principle in personalized medicine. Expert Rev Mol Diagn 2012; 12:621-8; PMID:22845482; http://dx.doi.org/10.1586/erm.12.46
- Martinez-Useros J, Garcia-Foncillas J. Obesity and colorectal cancer: molecular features of adipose tissue. J Transl Med 2016; 14:21; PMID:26801617; http://dx.doi.org/10.1186/s12967-016-0772-5
- Roy HK, Turzhitsky V, Wali R, Radosevich AJ, Jovanovic B, Della'Zanna G, Umar A, Rubin DT, Goldberg MJ, Bianchi L, et al. Spectral biomarkers for chemoprevention of colonic neoplasia: a placebo-controlled double-blinded trial with aspirin. Gut 2017; 66:285-92; PMID:26503631; http://dx.doi.org/10.1136/gutjnl-2015-309996
- Dienstmann R, Vermeulen L, Guinney J, Kopetz S, Tejpar S, Tabernero J. Consensus molecular subtypes and the evolution of precision medicine in colorectal cancer. Nat Rev Cancer 2017; 17(2):79-92; PMID:28050011; http://dx.doi.org/10.1038/nrc.2016.126
- Rescigno T, Micolucci L, Tecce MF, Capasso A. Bioactive nutrients and nutrigenomics in age-related diseases. Molecules 2017; 22:105; PMID:28075340; http://dx.doi.org/10.3390/molecules22010105
- Galon J, Mlecnik B, Bindea G, Angell HK, Berger A, Lagorce C, Lugli A, Zlobec I, Hartmann A, Bifulco C, et al. Towards the introduction of the ‘Immunoscore’ in the classification of malignant tumours. J Pathol 2014; 232:199-209; PMID:24122236; http://dx.doi.org/10.1002/path.4287
- Mlecnik B, Bindea G, Angell HK, Sasso MS, Obenauf AC, Fredriksen T, Lafontaine L, Bilocq AM, Kirilovsky A, Tosolini M, et al. Functional network pipeline reveals genetic determinants associated with in situ lymphocyte proliferation and survival of cancer patients. Sci Transl Med 2014; 6:228ra37; PMID:24648340; http://dx.doi.org/10.1126/scitranslmed.3007240
- Liston A, Gray DH. Homeostatic control of regulatory T cell diversity. Nat Rev Immunol 2014; 14:154-65; PMID:24481337; http://dx.doi.org/10.1038/nri3605
- Saito T, Nishikawa H, Wada H, Nagano Y, Sugiyama D, Atarashi K, Maeda Y, Hamaguchi M, Ohkura N, Sato E, et al. Two FOXP3+CD4+ T cell subpopulations distinctly control the prognosis of colorectal cancers. Nat Med 2016; 22(6):679-84; PMID:27111280; http://dx.doi.org/10.1038/nm.4086
- Ladoire S, Martin F, Ghiringhelli F. Prognostic role of FOXP3+ regulatory T cells infiltrating human carcinomas: the paradox of colorectal cancer. Cancer Immunol Immunother 2011; 60:909-18; PMID:21644034; http://dx.doi.org/10.1007/s00262-011-1046-y
- English L, Chemali M, Duron J, Rondeau C, Laplante A, Gingras D, Alexander D, Leib D, Norbury C, Lippé R, et al. Autophagy enhances the presentation of endogenous viral antigens on MHC class I molecules during HSV-1 infection. Nat Immunol 2009; 10:480-7; PMID:19305394; http://dx.doi.org/10.1038/ni.1720
- Tsai WT, Lo YC, Wu MS, Li CY, Kuo YP, Lai YH, Tsai Y, Chen KC, Chuang TH, Yao CH, et al. Mycotoxin patulin suppresses innate immune responses by mitochondrial dysfunction and p62/sequestosome-1-dependent mitophagy. J Biol Chem 2016; 291(37):19299-311; PMID:27458013; http://dx.doi.org/10.1074/jbc.M115.686683
- Goulielmaki M, Koustas E, Moysidou E, Vlassi M, Sasazuki T, Shirasawa S, Zografos G, Oikonomou E, Pintzas A. BRAF associated autophagy exploitation: BRAF and autophagy inhibitors synergise to efficiently overcome resistance of BRAF mutant colorectal cancer cells. Oncotarget 2016; 7:9188-221. PMID:26802026; http://dx.doi.org/10.18632/oncotarget.6942
- Tang B, Wang K, Jia YP, Zhu P, Fang Y, Zhang ZJ, Mao XH, Li Q, Zeng DZ. Fusobacterium nucleatum-induced impairment of autophagic flux enhances the expression of proinflammatory cytokines via ROS in Caco-2 cells. PLoS One 2016; 11:e0165701; PMID:27828984; http://dx.doi.org/10.1371/journal.pone.0165701
- Ogino S, Nishihara R, VanderWeele TJ, Wang M, Nishi A, Lochhead P, Qian ZR, Zhang X, Wu K, Nan H, et al. Review article: The role of molecular pathological epidemiology in the study of neoplastic and non-neoplastic diseases in the era of precision medicine. Epidemiology 2016; 27:602-11; PMID:26928707; http://dx.doi.org/10.1097/EDE.0000000000000471
- Nishihara R, Lochhead P, Kuchiba A, Jung S, Yamauchi M, Liao X, Imamura Y, Qian ZR, Morikawa T, Wang M. Aspirin use and risk of colorectal cancer according to BRAF mutation status. JAMA 2013; 309:2563-71; PMID:23800934; http://dx.doi.org/10.1001/jama.2013.6599
- Nishihara R, Wu K, Lochhead P, Morikawa T, Liao X, Qian ZR, Inamura K, Kim SA, Kuchiba A, Yamauchi M. Long-term colorectal-cancer incidence and mortality after lower endoscopy. N Engl J Med 2013; 369:1095-105; PMID:24047059; http://dx.doi.org/10.1056/NEJMoa1301969
- Liao X, Lochhead P, Nishihara R, Morikawa T, Kuchiba A, Yamauchi M, Imamura Y, Qian ZR, Baba Y, Shima K, et al. Aspirin use, tumor PIK3CA mutation, and colorectal-cancer survival. N Engl J Med 2012; 367:1596-606; PMID:23094721; http://dx.doi.org/10.1056/NEJMoa1207756
- Yamauchi M, Lochhead P, Morikawa T, Huttenhower C, Chan AT, Giovannucci E, Fuchs C, Ogino S. Colorectal cancer: a tale of two sides or a continuum? Gut 2012; 61:794-7; PMID:22490520; http://dx.doi.org/10.1136/gutjnl-2012-302014
- Cao Y, Nishihara R, Qian ZR, Song M, Mima K, Inamura K, et al. Regular Aspirin Use Associates With Lower Risk of Colorectal Cancers With Low Numbers of Tumor-Infiltrating Lymphocytes. Gastroenterology 2016; 151:879-92 e4; PMID:27475305; http://dx.doi.org/10.1053/j.gastro.2016.07.030
- Chan AT, Ogino S, Fuchs CS. Aspirin and the risk of colorectal cancer in relation to the expression of COX-2. N Engl J Med 2007; 356:2131-42; PMID:17522398; http://dx.doi.org/10.1056/NEJMoa067208
- Lamb CA, Yoshimori T, Tooze SA. The autophagosome: origins unknown, biogenesis complex. Nat Rev Mol Cell Biol 2013; 14:759-74; PMID:24201109; http://dx.doi.org/10.1038/nrm3696
- Ogino S, Nosho K, Kirkner GJ, Kawasaki T, Meyerhardt JA, Loda M, Giovannucci EL, Fuchs CS. CpG island methylator phenotype, microsatellite instability, BRAF mutation and clinical outcome in colon cancer. Gut 2009; 58:90-6; PMID:18832519; http://dx.doi.org/10.1136/gut.2008.155473
- Nosho K, Irahara N, Shima K, Kure S, Kirkner GJ, Schernhammer ES, Hazra A, Hunter DJ, Quackenbush J, Spiegelman D, et al. Comprehensive biostatistical analysis of CpG island methylator phenotype in colorectal cancer using a large population-based sample. PLoS One 2008; 3:e3698; PMID:19002263; http://dx.doi.org/10.1371/journal.pone.0003698
- Imamura Y, Lochhead P, Yamauchi M, Kuchiba A, Qian ZR, Liao X, Nishihara R, Jung S, Wu K, Nosho K, et al. Analyses of clinicopathological, molecular, and prognostic associations of KRAS codon 61 and codon 146 mutations in colorectal cancer: cohort study and literature review. Mol Cancer 2014; 13:135; PMID:24885062; http://dx.doi.org/10.1186/1476-4598-13-135
- Liao X, Morikawa T, Lochhead P, Imamura Y, Kuchiba A, Yamauchi M, Nosho K, Qian ZR, Nishihara R, Meyerhardt JA, et al. Prognostic role of PIK3CA mutation in colorectal cancer: cohort study and literature review. Clin Cancer Res 2012; 18:2257-68; PMID:22357840; http://dx.doi.org/10.1158/1078-0432.CCR-11-2410
- Mima K, Sukawa Y, Nishihara R, Qian ZR, Yamauchi M, Inamura K, Kim SA, Masuda A, Nowak JA, Nosho K, et al. Fusobacterium nucleatum and T Cells in Colorectal Carcinoma. JAMA Oncol 2015; 1:653-61; PMID:26181352; http://dx.doi.org/10.1001/jamaoncol.2015.1377
- Mima K, Nishihara R, Qian ZR, Cao Y, Sukawa Y, Nowak JA, et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016; 65:1973-80; PMID:26311717; http://dx.doi.org/10.1136/gutjnl-2015-310101