
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Immunogenicity testing in animals is a necessary preclinical assay for demonstration of vaccine efficacy the results of which are often the basis for the decision whether to proceed or withdraw the further development of the novel vaccine candidate. However, in vivo assays are rarely, if at all, optimized and validated. Here we clearly demonstrate the importance of in vivo assay (mumps virus immunogenicity testing in guinea pigs) optimization for gaining reliable results and the suitability of Fractional factorial design of experiments (DoE) for such a purpose. By the use of DoE with resolution IV (2IV(4-1)) we clearly revealed that the parameters significantly increasing assay sensitivity were interval between animal immunizations followed by the body weight of experimental animals. The quantity (0 versus 2%) of the stabilizer (fetal bovine serum, FBS) in the sample was shown as non-influencing parameter in DoE setup. However, the separate experiment investigating only the FBS influence, and performed under other parameters optimally set, showed that FBS also influences the results of immunogenicity assay. Such finding indicated that (a) factors with strong influence on the measured outcome can hide the effects of parameters with modest/low influence and (b) the matrix of mumps virus samples to be compared for immunogenicity must be identical for reliable virus immunogenicity comparison. Finally the 3 mumps vaccine strains widely used for decades in the licensed vaccines were for the first time compared in an animal model, and results obtained were in line with their reported immunogenicity in human population supporting the predictive power of the optimized in vivo assay.
Introduction
Preclinical testing is a necessary prerequisite for moving a candidate vaccine from the laboratory to the clinic and involves, among other biochemical and in vitro assays, also a proof of concept/immunogenicity studies in animals to evaluate and predict vaccine efficacy in humans.Citation1 In addition, in vivo immunogenicity testing could be invaluable tool for: (a) evaluation of vaccine production consistency; (b) monitoring the influence of changes in the production process on the vaccine quality;Citation1 (c) in the case of RNA viruses with extreme evolutionary capacities of wild-type strains, an estimation of vaccine efficacy against the circulating viruses.Citation2-5
Immunogenicity testing involving animals should be optimized not only (a) to comply with laws for the protection of laboratory animalsCitation1 that stress the importance of following the 3Rs (Replacement, Refinement and Reduction) philosophy, but also (b) to allow detection of subtle biological effects and to contribute significantly to quality assurance of vaccine productions. Strategies for optimization of assays to reach both these goals involve the use of Design of Experiments (DoE),Citation6-8 particularly Factorial Experimental Designs (FED).Citation9
Mumps, a common childhood disease, is caused by the mumps virus (MuV) which belongs to paramyxoviruses. Since 1970s the mumps has been controlled and prevented by vaccination with attenuated mumps vaccines, that resulted in significant reduction to nearly elimination of the disease from countries with high coverage rates of a 2-dose regimen.Citation10 However, resurgent mumps outbreaks among highly vaccinated population in recent yearsCitation11-18 in addition to relatively high rate of transient but unwanted neurological side effects by several vaccine strains Citation19 have raised many questions regarding the pathogenesis of MuV and vaccine efficacy, including considerations on the need to develop novel mumps vaccine candidates.
There has been no appropriate challenge/protection laboratory animal model for mumps, due to the fact that laboratory animals are not susceptible to natural mumps infection. The productive infection in animals was induced only by intracerebral inoculation of MuV strains.Citation19,20 It has been accepted that licensed MuV vaccines act through preventing viremia,Citation21 or MuV spreading by plasma from the site of infection and primary multiplication to different organs, including the central nervous system (the virus disseminates most likely in T-cells but also as free plasma virions).Citation10 Although the serologic correlate of protection induced by mumps vaccination has not been accepted yet, the neutralizing antibodies have been considered as the most probable candidate.Citation22-26 Regarding animal models, although guinea pigs, mice, cotton rats, rabbits and hamsters are not susceptible to MuV infection, they do generate neutralizing antibodies upon mumps administration.Citation27 In such a situation, we chose to establish and optimize immunogenicity assay in guinea pigs measuring quantity of functional virus-specific antibodies raised upon immunization of animals with MuV.
The focus of our research is optimization of the mumps virus immunogenicity assay in guinea pig by the use of FED with the goal to obtain the maximal resolution to detect subtle biological changes. The operating conditions (factors) which could have an influence on the assay performance were identified and their effects on the assay results were studied. FED, a systematic approach for studying the effects of multiple factors,Citation28 has been successfully used in the optimization and robustness testing of pharmaceutical analytical procedures.Citation29,30 However, it has not been widely used, although recommended, in optimization and validation studies of biological assays yet,Citation31 particularly in assays involving animals.
Finally, the optimized assay was used to compare the immunogenicity of 3 mumps virus vaccine strains (Jeryl Lynn 5, Urabe AM9 and L-Zagreb) that have been extensively used for years to vaccinate human populations. There has been no mumps virus immunogenicity preclinical in vivo study performed and reported by the optimized assay so far.
Results
Immunogenicity assay optimization
The influence of selected factors was investigated according to the experimental plan outlined in . LZG virus strain was used for assay optimization. Immunogenicity in 5 animals was assayed in each individual run. The quantity of functional virus-specific antibodies developed in their sera served as the measure of the response/outcome of the assay, expressed in titres. All experimental runs performed on the lower level of the factor X3 (virus quantity in the immunization dose), namely runs R1-R4 and R9-R12, resulted with undetectable virus-specific antibodies. These runs, together with the factor X3, had to be omitted from the data analysis and are denoted in gray in . Remaining 8 runs with quantifiable antibodies were recognized as design of the resolution IV, namely 2IV(4-1), that we found suitable for analysis (). Some experimental runs induced substantially lower (R5) or higher (R16) level of specific antibodies (). Factor X2 (FBS quantity in virus sample) had no influence on the assay outcome, while factor X1 (intervals between immunizations) had the most pronounced positive effect, followed by factor X4 (guinea pig body weight) (). The convenient graphical interpretation of the results by the Pareto diagram () demonstrated that factors X1 and X4 positively and significantly influenced the assay results when considering results of virus neutralization assay for antibody quantification. When HI assay was used, both factors were found to influence the assay results in positive direction, but only factor X1 significantly.
Figure 1. Schematic presentation of 2V(5–1) fractional factorial design of experiments for immunogenicity assay optimization. (A) Specification of operating factors and their levels. (B) Specification of experimental plan; experimental runs with invalid results (bellow the detection level of the assay) that were not included in the data analysis are denoted in gray.
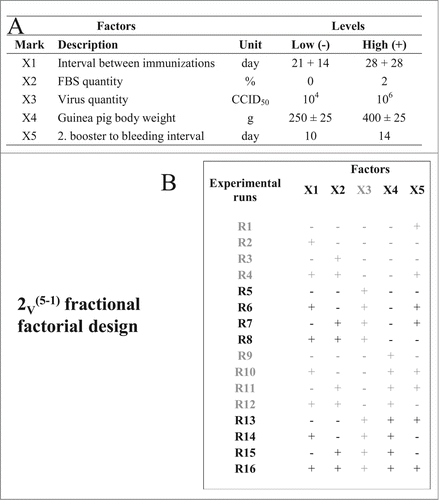
Figure 2. Specification of valid experiments recognized as 2IV(4–1) fractional factorial design (A) and immunogenicity assay results measured by virus neutralization assay (B) and HI assay (C). Results are expressed as mean (◊) and 95% confidence interval (CI) (⊥) of logarithmic values of individual results within each run. Full line denotes the mean and dashed lines denote 95% CI of the logarithmic values of total experimental plan, calculated from all individual values.
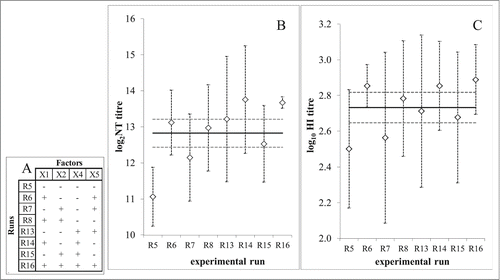
Figure 3. Data analysis of immunogenicity assay optimization according to 2IV(4-1) experimental design. (A) Mean value (◊) of immunogenicity assay outcome at higher (+1; ) and lower (−1;
) level of each experimental factor (X1 - intervals between immunizations; X2 – FBS quantity; X4 – guinea pig body weight; X5 – interval between 2. booster and bleeding) in comparison to mean value (full line) and 95% confidence interval (dashed lines) of the total experimental plan. Immunogenicity assay outcome was measured by virus neutralization assay (upper graphs) and HI assay (lower graphs). (B) Pareto plot of main effect estimates, presented by horizontal columns. The statistically significant effects (p = 0.05) are denoted by *. Immunogenicity assay outcome measured by virus neutralization assay (left) or HI assay (right).
Immunogenicity assay optimization considering immunization dose
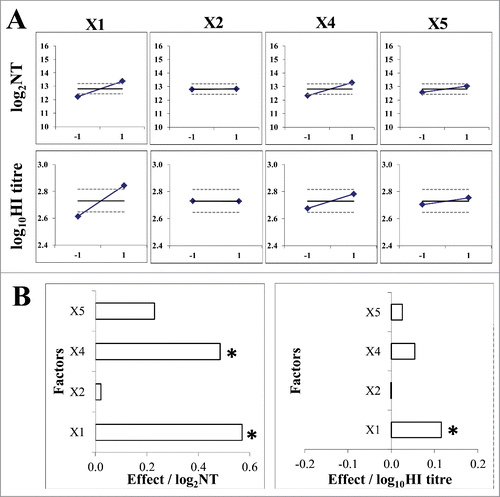
To find the range of virus quantities in immunization dose giving the linear dose-response relationship, guinea pigs were immunized under conditions so far proved to increase the sensitivity of this assay: animals of higher body weight (400 ± 50 g) were immunized 3 times in 28-day intervals, their blood taken for analysis on day 10 after the final immunization. Groups of 5 animals were immunized with decreasing quantities (in two fold steps) of LZG MuV, starting from 2,000,000 CCID50 per dose, namely the investigated interval was from 6.30 to 3.89 logCCID50 per dose. The logarithmic values of the resulting HI titres plotted against virus quantity per dose (in logCCID50) () demonstrated the excellent linear dose-response relation in the dose range from 5.1 to 6.3 logCCID50. Lower doses did not induce measurable antibody quantities in all (doses 3.89 and 4.19 logCCID50) or some (doses 4.5 and 4.8 logCCID50) immunized animals.
Figure 4. Virus quantity in immunization dose / immunogenicity assay outcome relationship. Mean values (◊) and 95% confidence interval (⊥) of the MuV-specific antibody titer (expressed in log10HI titer) obtained in each experimental group (n = 5) were plotted against the MuV quantities in the immunization dose. Linear relationship was obtained in the range 5.1 to 6.3 logCCID50 (125,000 to 1,000,000 CCID50) per dose.
Influence of FBS quantity in virus samples on immunogenicity assay outcome
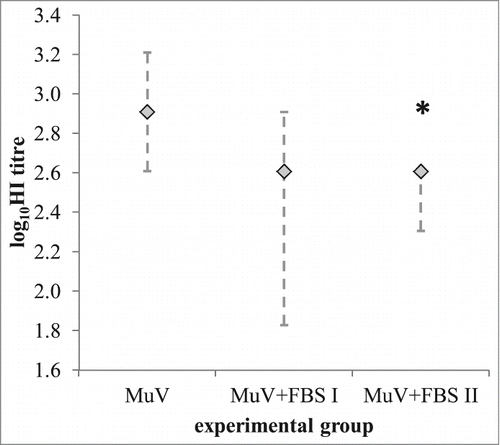
To study possible influence of FBS in the virus sample on virus specific immune response, 3 groups of animals, each of 5 females were immunized with 500,000 CCID50 LZG MuV per dose, either without FBS (MuV group) or with 2% FBS prepared in 2 ways: (1) by adding FBS before freezing of virus for storage (group MuV-FBS I) or by adding FBS before immunization in MuV sample previously stored without FBS (group MuV-FBS II). Animals immunized with pure MuV, without FBS, developed higher quantities of virus-specific functional antibodies in comparison to both groups containing FBS (). This difference between MuV and MuV-FBS II group was found statistically significant.
Figure 5. Influence of FBS in the immunization sample on the immunogenicity of the MuV. Groups of 5 animals were immunized with 500,000 CCID50 of LZG MuV per immunization dose, either without FBS in the sample (MuV) or with 2% FBS (MuV-FBS I and MuV-FBS II), prepared as follows. The unique sample of MuV was split in 2 aliquots, and one completed with 2% FBS (MuV-FBS sample), the other with equal volume of PBS (MuV sample) prior storage. MuV-FBS sample (virus titer 8.566 ± 0.226 log CCID50) was diluted with 2% FBS and suspension was used for immunization of a group named MuV-FBS I. MuV sample (virus titer 8.208 ± 0.373 logCCID50) was diluted either with pure PBS to obtain immunization suspension for the group MuV, or with 2% FBS for the immunization of the group MuV-FBS II. Medians (◊) with minimal and maximal values (⊥) of the MuV-specific antibody titer (expressed in log10HI titer) obtained in each experimental group (n = 5) are given; *p < 0.05 in comparison to MuV group.
Comparison of 3 MuV strains immunogenicity
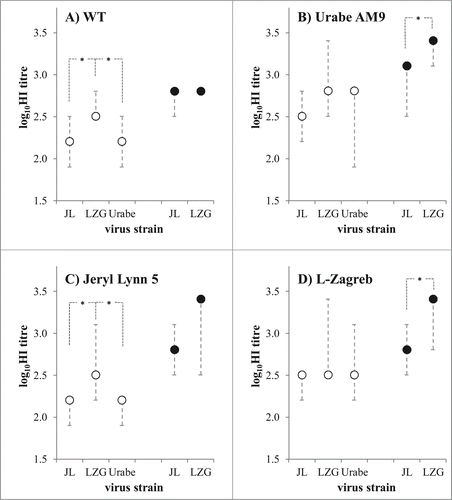
Immunogenicity of JL5, Urabe, and LZG MuV strains was investigated in optimized guinea pig animal model. Groups of 5 animals were immunized with 6.47 logCCID50 of respective MuV per dose. LZG and JL5 MuV were compared when immunization suspension contained only sorbitol (no FBS) in the sample (full circles in ), and all 3 viruses were compared with 1.4% FBS as a matrix in immunization suspension (blank circles in ). The induced titres were quantified by HI assay, with all 3 tested viruses – JL5 (), LZG () and Urabe () – as a hemagglutinating agent in the assay. Also, all sera were tested for their ability to inhibit hemagglutinating capacities of wild MuV virus strain MuVi/Split.CRO/05.11 (genotype G) (). All tested viruses were highly cross-reactive toward each other. Irrespective whether we tested the ability of animal antibodies to inhibit hemagglutinating activity of LZG, JL5, Urabe or WT, MuV specific antibody levels were highest in group immunized with LZG, followed by JL5 and Urabe immunized group. LZG induced significantly higher level of virus-specific antibodies in comparison to JL5 in both cases – when FBS was present (blank circles) or absent (full circles) in immunization suspensions. It is also obvious that animals immunized with both viruses without FBS in immunization suspension developed higher quantities of specific antibodies than when FBS was added (, comparison of blank to full circles in all graphs).
Figure 6. Comparison of 3 mumps vaccine strains in guinea pig immunogenicity assay. MuV specific antibody quantities were measured by 4 HI assays, differing in virus that was used as a hemagglutinating agent: (A) wild type virus MuVi/Split.CRO/05.11 strain (denoted as WT); (B) Urabe AM9; (C) Jeryl Lynn 5; and (D) L-Zagreb. Animals (n = 5 per group) were immunized with the same quantity (6.47 logCCID50 per dose) of each MuV strain (denoted on x-axis as LZG for L-Zagreb, Urabe for Urabe AM9 and JL5 for Jeryl Lynn 5). All three viruses were tested when FBS was included in immunization suspension (empty circles), and 2 of them were tested also without FBS in immunization suspension (full circles). Medians with minimal and maximal values of log10HI titre are given; *p < 0.05 between groups connected with dashed lines.
Discussion
The humoral immune response to the range of doses of antigen of interest gives sigmoidal dose-response curve.Citation32 Two low doses either fail to induce immune response or they induce it, however our techniques to measure it could be of too low sensitivity. On the other hand, with high antigen concentrations the plateau or maximal immune response is reached, that could not be increased by additional increase of antigen dose (it could be even reduced or suppressed). The optimal assay should be performed with doses giving linear relationship between dose and response.
In our original experimental plan we wanted to investigate, using DoE methodology, the influence of the selected operating factors and the range of their values on the outcome of the immunogenicity assay, because of the following reasons: (a) MuV dose of 10,000 CCID50 is in the range of virus quantity in a human dose of a final vaccine; Citation33 higher dose of 1,000,000 CCID50 was chosen, because we expected that human dose might be too low for animals known not to be susceptible for MuV replication; this has finally been proven; (b) the guinea pig body weights tested were 250 ± 25 g and 400 ± 25 g; the body weight is directly related to the animal age; the usage of animals of the lower body weight would reduce the costs of this assay; (c) 2 variants of intervals between immunizations were tested: 21 + 14 scheme, that would be favorable due to its shortness, and 28 + 28 scheme, that from our unpublished experience increases the immune response and is in line with observations that longer intervals positively contribute to antibody levels induced;Citation34 (d) the influence of the matrix was chosen to be tested due to our and others' experienceCitation35 that high concentration of the non-relevant protein reduces the immune response of the protein of interest due to immune competition; FBS is often used as a virus stabilizer (although not in final vaccines), and its protein concentration is much higher than that of virus in the sample, so the antigenic competition is highly probable.
Our results showed that 10,000 CCID50 of virus in immunization dose did not induce immune response measurable by our functional antibody quantification assays. DoE approach was proved very useful to select which levels of operating factors favor the sensitivity of the assay (give higher results). The assay should be performed with older (bigger animals) which is probably the consequence of the higher maturity of the animal immune system. Longer intervals between immunizations, and longer interval between final immunization and bleeding were also shown to increase the immunogenicity assay sensitivity. Separate experiment, conducted to determine the dose range giving linear dose/response relationship, showed that virus quantities per dose have to be higher than 125,000 CCID50 ().
It was surprising that FBS presence in the virus sample was shown not to affect the virus immunogenicity results (, factor X2). So we further investigated solely the contribution of FBS presence in the virus sample for immunization on the immunogenicity assay outcome. FBS in the sample significantly reduced the immunogenicity of the virus (). This reduction is induced by 2 mechanisms: (a) antigenic competition (, comparison of MuV and MuV-FBS II group; also , comparison of full and black circles); (b) due to two-fold difference in infective virus titer depending whether the same quantity of MuV was stored with (8.566 ± 0.226 logCCID50) or without (8.208 ± 0.373 logCCID50) FBS as a stabilizer (comparison of MuV and MuV-FBS I group). Since the titer is the regulatory measure of virus quantity in the sample,Citation33 it was used to adjust the virus quantity in the immunization dose. The MuV-FBS I sample was, consequently, diluted twice that much as the MuV to reach the equal immunization dose, while actually in both samples the concentration of mumps virus proteins was the same. Explanation of the fact that DoE methodology for operating factors screening on the immunogenicity assay outcome did not recognize FBS quantity as important factor (), in contrast to the separate experiment investigating isolated FBS contribution () might be that effect of FBS on immunogenicity is probably much lower in comparison to other factors' contribution, and was “masked” by the assay variations induced by other factors. However we have clearly shown that for optimal and reliable comparison of different MuV samples viruses have to be prepared in the same matrices.
Finally, the optimized assay was used to compare the immunogenicity of the 3 MuV strains that are among the most widely used for prophylaxis of mumps in human population. Our results on matrix impact presented above imply that the total antigen content (both infective and not infective virus particles that are inevitably present in any live virus suspension) influences the immunogenicity assay outcome. To minimize the possible discrepancies and differences between antigen content of the 3 virus samples to be compared expressed as CCID50 (infective virus quantity) and their total antigen content that was not monitored in this study, all 3 viruses were prepared (propagated) exactly in the same way and formulated equally. JL5 (genotype A), Urabe (genotype B) and LZG (genotype N) were highly cross-reactive to each other although belonging to different genotypesCitation36,37 and were also cross-reactive to the wild-type virus of genotype GCitation37 supporting the historically accepted fact that MuV exists as a single serotype, also verified in recent years.Citation38,39 Irrespective of which virus was used as a hemagglutinating agent, LZG showed significantly higher immunogenicity in comparison to JL5 strain. Urabe and JL5 were shown in our study of comparable immunogenicity. This is also in accordance with available epidemiological data from human studies that have so far not given the unambiguous answer on the these 2 vaccine strains immunogenicity. Ong et al.Citation40 reported on higher immunogenicity of Urabe strain in Singapore population, while others report on comparable immunogenicity.Citation41-43 However, one should be aware that higher immunogenicity of a vaccine strain does not immediately imply that the strain is better, and the safety side has to be considered simultaneously. JL5 strain has so far proved not to induce neurovirulence in rats,Citation44 and has been shown as the safest mumps vaccine strain in human population with the rare side effects reported.Citation19 In contrast, Urabe vaccine strain has already been removed from the market due to high incidence of adverse effects, while notably high incidence of side effects has also been reported due to vaccines based on LZG strain.Citation19
In conclusion we presented here the successful optimization of the animal in vivo assay using DoE methodological approach according to the animal welfare directions, and demonstrated the appropriateness and reliability of the optimized assay for comparison of the different MuV strains immunogenicity in small animal model. We stressed the importance on performing the studies under conditions giving the results in the linear part of dose/response curve, and also that different matrices around the viruses could have significant impact on the outcome of virus immunogenicity testing.
Materials and Methods
Reagents and chemicals
Minimum Essential Medium with Hank's salts (MEM-H(0); Applichem, Germany) was prepared according to the manufacturer's instructions, filter sterilized and stored in the dark at 2–8°C. For MEM-H(N) preparation, MEM-H(0) was supplemented with 50 μgmL−1 of neomycin (N) (Gibco-BRL, USA), before filter sterilization. For MEM-H(N)-FBS preparation, 10% fetal bovine serum (FBS; PAA Laboratories GmbH, Australia) was added (V/V) to MEM-H(N), before filter sterilization.
Experimental animals
Animal work was in accordance to the Croatian Law on Animal Welfare (2013) which strictly complies with EU Directive 2010/63/EU. Outbred female guinea pigs were of CRL:(H)BR strain, bred at the Institute of Immunology, Inc.
MuV samples
MuV strains were: L-Zagreb (LZG; Institute of Immunology Inc., Croatia), Urabe AM9 (Urabe; 1st International Reference Reagent for Mumps Vaccine, NIBSC), Jeryl Lynn 5 strain (JL5; a kind gift from B.K. Rima) and MuVi/Split.CRO/05.11 strain (WT, from “wild type;” isolated in our laboratory during the outbreak in 2011). All viruses were propagated in Vero cells (American Tissue and Culture Collection (ATCC)) for up to 2 passages. Vero cells were maintained in MEM-H(N)-FBS. Vero cell culture was infected with MuV (m.o.i. 0.001) and incubated in MEM-H(N) + 2% FBS at 35°C for 48 hours. The culture was washed twice with MEM-H(N) and incubated in fresh MEM-H(N) until cytophatic effect (CPE) appearance. The virus harvest was collected and clarified by centrifugation at 3,000 × g. Virus-containing supernatant was collected and virus further purified by ultracentrifugation at 141,000 × g for 2 hours at 4°C. Virus-containing pellet was resuspended in 5 mL of phosphate buffer saline (PBS) and stored in 200 µL aliquots until use at −60°C and bellow. Virus quantity was determined by titration in microplates on Vero cells as described.Citation45 Briefly, serial dilutions of virus suspension were added to a 96-well plate containing fully confluent Vero cells grown at a cell density of 1 × 106 cells/mL. After 10 d of incubation at 37°C and 5% CO2, the plate was examined under a microscope to enumerate the number of wells exhibiting cytopathic effect (CPE).
For comparative immunogenicity study of JL5, Urabe and LZG strains immunization suspensions were prepared in 2 different stabilizer containing matrices: MEM-H(0) containing 1.4% FBS, and MEM-H(0) containing 5% sorbitol. Animals in this study were immunized with 6.47 logCCID50 of virus per immunization dose.
MuV immunogenicity assay
Animals were immunized subcutaneously (s.c.) with 500 µL of immunization suspension, split at 2 sites (250 µL per site). Immunization protocol consisted of 3 immunizations (immunization and 2 boosters). Guinea pigs' blood was collected by heart puncture and sera prepared for further serological analyses by centrifugation at 1200 × g at 4°C for 30 min, decomplemented by heating at 56°C for 30 min, filter sterilized and stored until analysis at −20°C. Animal weight, interval between immunizations, interval between final immunization and bleeding, and virus quantity were the subject of here described investigation. The quantity of virus-specific antibody was determined by virus neutralization assay and/or hemagglutination inhibition assay.
Serological assays
Virus neutralization assay was performed in 96-well tissue culture microplates. Septuplicates of two-fold serial dilutions (100 µL/well) of guinea pig sera in MEM-H(0) + 2% FBS medium were preincubated with 100 CCID50 of the laboratory reference preparation of LZG vaccine strain (Institute of Immunology Inc.) (100 µL/well) at 37°C and 5% CO2 for 1.5 h. Donkey serum of known neutralizing capacity was used as positive control. After adding Vero cells (3.2 × 105/mL) (50 µL/well) the plates were incubated at 35°C and 5% CO2. Wells with pure cell suspension served as cell growth control. Wells with cell suspension and tested sera (in each dilution) served as control of sera toxicity. CPE was monitored on day 10 and neutralization titer (dilution inhibiting CPE in 50% of wells) evaluated by Spearman-Kärber method. Hemagglutination inhibition (HI) was performed according to Cross Citation46 with exception that 0.5% guinea pig erythrocyte suspension was used. The highest dilution of serum still capable to completely inhibit hemagglutination was recorded as HI titer.
Design of experiments (DoE)
Five operating factors (X1-X5), each at 2 levels (marked with minus (−) for the low, and plus (+) for the high level), were selected to explore their influence on outcome of the immunogenicity bioassay ():
X1 – Interval between immunizations: 21 + 14 scheme (21-day interval between immunization and the first booster followed by 14 d interval between the first and the second booster); 28 + 28 scheme (28-day interval between each immunization);
X2 – Virus stabilizer quantity: FBS was used as a stabilizer, and its influence was investigated at 0 and 2%;
X3 – Virus quantity per immunization dose: 10,000 and 1,000,000 CCID50;
X4 – Guinea pig weight: 250 ± 25 and 400 ± 25 g;
X5 – Interval between final immunization and bleeding: 10 or 14 days;
The fractional FED with resolution V was planned (2V(5-1) - number of factors: 5; number of levels for each factor: 2; total number of experimental runs: 24 = 16). Experimental runs were marked R1–R16, each consisting of 5 animals immunized according to equal immunization procedure in which investigated factors were at levels indicated in .
Influence of FBS quantity in virus samples on immunogenicity assay outcome
To study possible influence of FBS in the virus sample on virus specific immune response we pelleted LZG MuV by ultracentrifugation, resuspended in PBS and then split in 2 equal aliquots: in one appropriate quantity of FBS was added to reach 2% FBS concentration, in another the equal quantity of PBS was added. This way 2 virus samples were prepared containing the equal amount of virus material, only they differed in matrix – the first was in 2% FBS (MuV-FBS sample) and another was without stabilizer (MuV sample). Both were aliquoted à 200 µL and stored at −60°C or bellow, until titration (virus quantity determination) or their usage for animal immunization. The determined virus titer of MuV-FBS sample was 8.566 ± 0.226 and for MuV sample was 8.208 ± 0.373 logCCID50. MuV-FBS sample was diluted with 2% FBS and suspension was used for immunization of a group named MuV-FBS I, MuV sample was diluted either with pure PBS to obtain immunization suspension for the group MuV, or with 2% FBS for the immunization of the group MuV-FBS II.
Data analysis
Results of virus neutralization and HI assay, expressed as titres, were first linearized by calculating logarithmic values and then used for all calculations and statistical analysis. For each experimental run mean value was calculated and 95% confidence interval as an indicator of measurement uncertainty. The main effect of each factor was calculated according to the formula:
(1)
(1) where index X represents factors 1 to 5, n is the total number of experimental runs (16),
are the log (titres) at (+) level obtained for each factor and
are the log(titres) at (−) level obtained for the same factor. The significance of the each factor effects was calculated and presented graphically by Pareto plot using computer program, SigmaPlot Ver. 11″. Differences between groups were statistically evaluated using Kruskal-Wallis ANOVA, followed by multiple Mann-Whitney U-nonparametric tests; p-values less than 0.05 were considered significant. Analyses were performed using Statistica 6.0 for Windows, StatSoft Inc.
Disclosure of Potential Conflicts of Interest
Institute of Immunology Inc. had previously been a producer of a mumps vaccine based on L-Zagreb MuV strain.
Acknowledgments
The authors wish to thank Mrs Maja Šantak, PhD, for providing viruses for this study, for sharing her knowledge on work with viruses and for critical reading of the manuscript. The authors are also grateful to Mrs Maja Jagušić for constructive critics during the manuscript preparation.
Funding
The work was financially supported by the Croatian Ministry of Science, Education and Sports (grant 021-0212432-2033 to BH), by the Croatian Science Foundation (grants 8193 to MB and 6255 to DF) and by Institute of Immunology, Inc.
References
- WHO guidelines on nonclinical evaluation of vaccines. WHO Technical Report Series, No. 927, 2005
- Domingo E, Holland JJ. RNA virus mutations and fitness for survival. Annu Rev Microbiol 1997; 51:51-78; PMID:9343347; http://dx.doi.org/10.1146/annurev.micro.51.1.151
- Drake JW. Rates of spontaneous mutation among RNA viruses. Proc Natl Acad Sci U S A 1993; 90:4171-5; PMID:8387212; http://dx.doi.org/10.1073/pnas.90.9.4171
- Drake JW, Holland JJ. Mutation rates among RNA viruses. Proc Natl Acad Sci U S A 1999; 96:13910-3; PMID:10570172; http://dx.doi.org/10.1073/pnas.96.24.13910
- Steinhauer DA, Holland JJ. Rapid evolution of RNA viruses. Annu Rev Microbiol 1987; 41:409-33; PMID:3318675; http://dx.doi.org/10.1146/annurev.mi.41.100187.002205
- Madden JC, Hewitt M, Przybylak K, Vandbriel RJ, Piersma AH, Cronin MTD. Strategies for the optimisation for in vivo experiments in accordance with the 3Rs philosophy. Regul Toxicol Pharmacol 2012; 63:140-54; PMID:22446816; http://dx.doi.org/10.1016/j.yrtph.2012.03.010
- Das RG, Fry D, Preziosi R, Hudson M. Planning for reduction. ATLA 2009; 37:27-32; PMID:19292573
- Kilkenny C, Parsons N, Kadiszewsky E, Festing MFW, Cuthill IC, Fry D, Hutton J, Altman DG. Survey of the quality of experimental design, statistical analysis and reporting of research using animals. PloS One 2009; 4, e7824; PMID:19956596; http://dx.doi.org/10.1371/journal.pone.0007824
- Shaw R, Festing MFW, Peers I, Furlong L. Use of factorial designs to optimize animal experiments and reduce animal use. ILAR J 2002; 43:223-32; PMID:12391398; http://dx.doi.org/10.1093/ilar.43.4.223
- Carbone KM, Rubin S. Mumps virus. In: Knipe DM, Howley PM, editors. Fields Virology, 5th ed. Lippincott Williams & Wilkins, 2007, p. 1528-50.
- Watson-Creed G, Saunders A, Scott J, Lowe L, Pettipas J, Hatchette TF. Two successive outbreaks of mumps in Nova Scotia among vaccinated adolescents and young adults. CMAJ 2006; 175:483-8; PMID:16940266; http://dx.doi.org/10.1503/cmaj.060660
- Dayan GH, Quinlisk MP, Parker AA, Barskey AE, Harris ML, Schwartz JM, Hunt K, Finley CG, Leschinsky DP, O'Keefe AL, et al. Recent resurgence of mumps in the United States. New Engl J Med 2008; 358:1580-9; PMID:18403766; http://dx.doi.org/10.1056/NEJMoa0706589
- Rota JS, Turner JC, Yost-Daljev MK, Freeman M, Toney DM, Meisel E, Williams N, Sowers SB, Lowe L, Rota PA, et al. Investigation of a mumps outbreak among university students with two measles-mumps rubella (MMR) vaccinations, Virginia, September-December 2006. J Med Virol 2009; 81:1819-25; PMID:19697404; http://dx.doi.org/10.1002/jmv.21557
- Stein-Zamir C, Shoob H, Abramson N, Tallen-Gozani E, Sokolov I, Zentner G. Mumps outbreak in Jerusalem affecting mainly male adolescents. Eurosurveillance 2009; 14:19940
- Carr MJ, Moss E, Waters A, Dean J, Jin L, Coughlan S, Connell J, Hall WW, Hassan J. Molecular epidemiological evaluation of the recent resurgence in mumps virus infections in Ireland. J Clin Microbiol 2010; 48:3288-94; PMID:20660212; http://dx.doi.org/10.1128/JCM.00434-10
- Echevarría JE, Castellanos A, Sanz JC, Pérez C, Palacios G, Martínez de Aragón MV, Pena Rey I, Mosquera M, de Ory F, Royuela E. Circulation of mumps virus genotypes in Spain from 1996 to 2007. J Clin Microbiol 2010; 48:1245-54; PMID:20107099; http://dx.doi.org/10.1128/JCM.02386-09
- Otto W, Mankertz A, Santibanez S, Saygili H, Wenzel J, Jilg W, Wieland WF, Borgmann S. Ongoing outbreak of mumps affecting adolescents and young adults in Bavaria, Germany, August to October 2010. Eurosurveillance 2010; 15:19748; PMID:21172171
- Whelan J, van Binnendijk R, Greenland K, Fanoy E, Khargi M, Yap K, Boot H, Veltman N, Swaan C, van der Bij A, de Melker H, Hahné S. Ongoing mumps outbreak in a student population with high vaccination coverage, Netherlands, 2010. Eurosurveillance 2010; 15:19554; PMID:20460086
- Rubin SA, Afzal MA. Neurovirulence safety testing of mumps vaccines - historical perspective and current status. Vaccine 2011; 29:2850-5; PMID:21334386; http://dx.doi.org/10.1016/j.vaccine.2011.02.005
- Houard D, Varsanyi TM, Milican F, Norrby E, Bollen A. Protection of hamsters against experimental mumps virus (MuV) infection by antibodies raised against the MuV surface glycoproteins expressed from recombinant vaccinia virus vectors. J Gen Virol 1995; 76:421-3; PMID:7844562; http://dx.doi.org/10.1099/0022-1317-76-2-421
- Plotkin SA. Correlates of vaccine-induced immunity. Clin Infect Dis 2008; 47:401-9; PMID:18558875; http://dx.doi.org/10.1086/589862
- Plotkin SA. Correlates of protection induced by vaccination. Clin Vaccine Immunol 2010; 17:1055-65; PMID:20463105; http://dx.doi.org/10.1128/CVI.00131-10
- Latner DR, McGrew M, Williams NJ, Sowers SB, Bellini WJ, Hickman CJ. Estimates of mumps seroprevalence may be influenced by antibody specificity and serologic method. Clin Vaccine Immunol 2014; 21:286-97; PMID:24371258; http://dx.doi.org/10.1128/CVI.00621-13
- Poethko-Mueller C, Mankertz A. Seroprevalence of measles-, mumps- and rubella-specific IgG antibodies in German children and adolescents and predictors for seronegativity. PLoS One 2012; 7(8):e42867; PMID:22880124; http://dx.doi.org/10.1371/journal.pone.0042867
- Usonis V, Bakasenas V, Denis M. Neutralization activity and persistence of antibodies induced in response to vaccination with a novel mumps strain, RIT 4385. Infection 2001; 29:159-62; PMID:11440387; http://dx.doi.org/10.1007/s15010-001-1098-7
- Nöjd J, Tecle T, Samuelsson A, Orvell C. Mumps virus neutralizing antibodies do not protect against reinfection with a heterologous mumps genotype. Vaccine 2001; 19:1727-31; PMID:11166897; http://dx.doi.org/10.1016/S0264-410X(00)00392-3
- Flynn M, Mahon BP. Cell mediated and humoral immune responses to mumps virus: recent developments. Rec Res Dev Virol 2003; 5:97-115
- Eriksson L, Johansson E, Kettaneh-Wold N, Wikström C, Wold S. Design of Experiments, Principles and Applications, Stockholm: Umetrics AB, 2000.
- Araujo P. Key aspects of analytical method validation and linearity evaluation. J Chromatogr B 2009; 877:2224-34; PMID:18929516; http://dx.doi.org/10.1016/j.jchromb.2008.09.030
- Dejaegher B, Vander Heyden Y. Ruggedness and robustness testing. J Chromatogr A 2007; 1158:138-57; PMID:17379230; http://dx.doi.org/10.1016/j.chroma.2007.02.086
- Krzywinski M, Altman N. Two-factor designs. Nat Methods 2014; 11:699-700
- Young CR, Ebringer A, Archer JR. Antigen dose and strain variation as factors in the genetic control of the immune response to sperm whale myoglobin. Immunology 1978; 34:571-9; PMID:417022
- Mumps Vaccine (live). 01/2008:0538. In: European Pharmacopoeia 8th Edition, Strasbourg: Council of Europe, 2013.
- Hanly WC, Artwohl JE, Bennett BT. Review of polyclonal antibody production procedures in mammals and poultry. ILAR J 1995; 37:93-118; PMID:11528030; http://dx.doi.org/10.1093/ilar.37.3.93
- Hunt JD, Brown LE, Jackson DC. Antigenic competition. eLS 2001
- Ivancic J, Kosutic Gulija T, Forcic D, Baricevic M, Jug R, Mesko-Prejac M, Mazuran R. Genetic characterization of L-Zagreb mumps vaccine strain. Virus Res 2005; 109:95-105; PMID:15826917
- Ivancic-Jelecki J, Santak M, Forcic D. Variability of hemagglutinin-neuraminidase and nucleocapsid protein of vaccine and wild-type mumps virus strains. InfectGenet Evol 2008; 8:603-13; PMID:18508415
- Šantak M, Lang-Balija M, Inavcic-Jelecki J, Košutić-Gulija T, Ljubin-Sternak S, Forcic D. Antigenic differences between vaccine and circulating wild-type mumps viruses decreases neutralization capacity of vaccine-induced antibodies. Epidemiol Infect 2013; 141:1298-309
- Rubin S, Mauldin J, Chumakov K, Vanderzanden J, Iskow R, Carbone K. Serological and phylogenetic evidence of monotypic immune responses to different mumps virus strains. Vaccine 2006; 24:2662-8; PMID:16309801
- Ong G, Goh KT, Ma S, Chew SK. Comparative efficacy of Rubini, Jeryl-Lynn and Urabe mumps vaccine in an Asian population. J Infect 2005; 51: 294-8; PMID:16291282
- Toscani L, Batou M, Bouvier P, Schlaepfer A. Comparison of the efficacy of various strains of mumps vaccine: a study in schools. Soz Praventiv Med 1996; 41:341-7
- Chamot E, Toscani L, Egger P, Germann D, Bourquin C. [Estimation of the efficacy of three strains of mumps vaccines during an epidemic of mumps in the Geneva canton (Switzerland)]. Rev Epidemiol Sante Publique 1998; 46:100-7; PMID:9592852
- Schloegel M, Osterwalder JJ, Galeazzi RL, Vernazza PL. Comparative efficacy of three mumps vaccines during disease outbreak in eastern Switzerland: cohort study. BMJ 1999; 519:352-3
- Rubin SA, Afzal MA, Powell CL, Bentley ML, Auda GR, Taffs RE, Carbone KM. The rat-based neurovirulence safety test for the assessment of mumps virus neurovirulence in humans: an international collaborative study. J Infect Dis 2005; 191:1123-8; PMID:15747248
- Forcic D, Kosutić-Gulija T, Santak M, Jug R, Ivancic-Jelecki J, Markusic M, Mazuran R. Comparisons of mumps virus potency estimates obtained by 50% cell culture infective dose assay and plaque assay. Vaccine 2010; 28:1887-92; PMID:19961964
- Cross G. Hemagglutination inhibition assay. Semin Avian Exot Pet 2002; 11:15-8