
ABSTRACT
Malaria remains one of the most significant infectious diseases worldwide. Concordant with scaled intervention efforts and the emphasis of elimination and eradication on the agenda of many malaria control programs, the development of a malaria vaccine that reduces transmission of the parasite from human host to mosquito vector has been incorporated as an important new strategic goal. Transmission of malaria from man to mosquito relies on gametocytes, highly specialized sexual-stage parasites, that once mature, circulate in the peripheral vasculature of the human host. The complex interplay between mature gametocytes, their uptake in the mosquito bloodmeal and forward maturation/fertilization events provide unique opportunities for intervention. Although recent advances have yielded greater understanding into the mechanisms that mediate sequestration of immature gametocytes in the human host, the spatial dynamics of circulating mature gametocytes in the cutaneous microvaculature remains far less defined, which is the focus of this review.
Introduction
Despite an unprecedented increase in implementation of malaria control policies over the last decade, malaria remains a considerable public health burden.Citation1 Global risk assessment analysis indicates that currently an estimated 3.3 billion people in 97 countries and territories are at risk of malaria with 1.2 billion at high risk.Citation2 Sub-Saharan African children under the age of five disproportionately bear the brunt of this disease, accounting for approximately 90% of the estimated 627,000 deaths due to malaria that occur each year.Citation1-3 Recent intensification of control efforts to levels not seen since the World Health Organization's Global Malaria Eradication Program (GMEP) in the late 1950s have escalated to calls for malaria elimination and eradication.Citation4,5 This recent paradigm shift Citation6 has engendered an important new strategic goal that includes the development of a safe and effective malaria vaccine that prevents transmission and the onward occurrence of new cases of infection, an aim that was corroborated in the 2013 update to the Malaria Vaccine Technology Roadmap.Citation7
Although progress toward the development of a deployable transmission blocking vaccine has been slow, the dynamic interplay between parasite and mosquito vector allow for attack from a multitude of angles. In particular, the interface between the mature infective circulating Plasmodium falciparum gametocyte and the arthropod vector in the dermis, the subject of this review, will be considered. Furthermore, novel approaches to assess the same dermal component sampled by the mosquito in field studies will be addressed along with the implications of gametocyte spatial dynamics on the development of anti-gametocyte transmission blocking vaccines.
Gametocyte development in the human host
Malaria in humans is caused by five species of Plasmodium (P. falciparum, P. vivax, P. ovale, P. malariae, and P. knowlesi), vector-borne pathogens that principally cause pathology to the human host by infecting and lysing erythrocytes. P. falciparum and P. vivax, which together account for the majority of morbidity and mortality due to malaria, differ in endemicity with P. falciparum being transmitted most intensely in sub-Saharan Africa,Citation3 while P. vivax has a much larger area of endemicity that includes Central and Southeast Asia, the Western Pacific, and South America.Citation8 Although the notion of P. vivax as a causative agent of benign disease has been dispelled, parasites in this review will herein refer to P. falciparum for which the transmissible stages have been best characterized.
Beginning with the parasitized female Anopheline mosquito while ingesting a blood meal, the vertebrate host is inoculated with sporozoites, which rapidly traffic to the liver and undergo a 7–10 day period of intrahepatocytic development. Upon maturation into schizonts, the infected hepatocytes are lysed, releasing a large population of merozoites, that invade the bloodstream and initiate repetitive cycles of intra-erythrocytic asexual multiplication. The ensuing rounds of erythrocyte lysis account for many of the clinical manifestations associated with this disease.
To perpetuate transmission from human host back to the female Anopheline mosquito, a small fraction of parasites differentiate into sexual erythrocytic stages (gametocytes). Gametocytes remain in a state of arrested cell development until ingested by a feeding mosquito, at which point, male micro-gametes are released in the mosquito's luminal midgut and fertilize activated macro-gametes, generating zygotes.Citation9 Subsequent parasite maturational events in the mosquito ultimately generate sporozoites that migrate to the mosquito salivary gland, rendering the mosquito infectious to humans. The trigger for asexual blood stage parasite development into intraerythrocytic gametocytes, previously thought to be determined by different kinds of environmental stress, including parasite density, anemia, reticulocytosis, host immune responses and drug treatment,Citation10-17 has now been shown by forward and reverse genetic approaches to be an epigenetic phenomenon, in large part mediated by the transcription factor AP2-G.Citation18,19 Although gametocytes are not thought to contribute to any pathology, they play a critical role in ensuring the long term survival of the parasite.
Intraerthrocytic gametocyte maturation is a protracted process of 10–14 days Citation20,21 that can be divided into five morphologically distinct stages (I–V) as shown in .Citation21 Approximately 40 hours after invasion of a red cell by a sexually committed merozoite, stage I gametocytes can be discerned with some difficulty from young trophozoites in giemsa stained blood films, although definitive classification usually requires antibody based immunofluorescence assays with early gametocyte markers such as Pfs16.Citation12 Successive gametocyte enlargement and distinct morphologic features define stage II-V gametocytes, which can be readily discerned by light microscopy. Commencement of slight erythrocyte distension in stage III culminates in the classic “sausage or crescent shape” of mature stage V gametocytes, which is in large part mediated by the subpellicular microtubule-based cytoskeleton and surrounding double membrane.
Figure 1. The five morphologically distinct stages (I – V) of P. falciparum gametocytes in giemsa stained, methanol fixed blood films. Early subpellicular cytoskeleton formation in stage II results in D-shaped parasites that enlarge in stage III to slightly distend the erythrocyte. Dramatic elongation and pointed ends characterize stage IV gametocytes that ultimately mature into stage V gametocytes. Characteristic morphology of this final stage includes an elongated parasite with rounded ends and a length to width ratio of ∼3:1. Dark pigmented hemozoin crystals are present in multiple gametocyte developmental stages, and are generated from the biocrystallization of hematin, a toxic intermediate derived from the parasite digestion of hemoglobin.
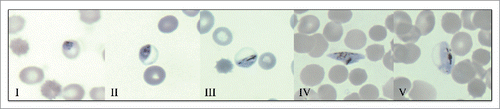
Developing immature gametocytes (stages I-IV) are thought to sequester in the spleen and extravascular space of the haematopoietic system of infected individuals,Citation22-25 whereas mature gametocytes (stage V) are released out into the peripheral circulation. Recent studies of P. falciparum induced molecular and cellular remodeling of the host erythrocyte during gametocyte maturation, have led to the hypothesis that the enrichment of immature gametocytes in niches such as the haematopoietic system of human bone marrow Citation24,26-28 and subsequent release of mature stage V gametocytes into the circulation is governed not by receptor-ligand interactions as long presumed, but instead by mechanical properties. It is also plausible that infective mature gametocytes sequester in subdermal and or dermal sites to enhance transmission potential, although this hypothesis lacks a critical body of supporting evidence.
Rheologic properties of gametocytes
Erythrocytes have an extraordinary capacity to undergo extensive deformation without cell fragmentation, an essential attribute to allow it to effectively perform its function of oxygen delivery. The erythrocyte plasma membrane is composed of amphiphilic surfactant molecules anchored to a network of skeletal proteins via tethering sites (transmembrane proteins) in the bilayer.Citation29 Erythrocytes infected with asexual P. falciparum parasites exhibit dramatically decreased membrane deformability along with enhanced cytoadhesive properties that allow them to sequester in the microvasculature,Citation30 thereby avoiding splenic clearance. These erythrocyte modifications are mediated by a subset of parasite proteins, many of which carry a pentameric host-targeting signal sequence (Plasmodium export element [PEXEL]),Citation31-33 that are transported across the parasitophorous vacuole membrane (PVM) into the erythrocyte membrane, where they complex with components of the erythrocyte cytoskeleton. The major adhesive ligand expressed on the surface of infected erythrocytes is P. falciparum erythrocyte membrane 1 (PfEMP1), which is present at raised platforms termed knobs and can mediate interactions with a number of receptors expressed on vascular endothelial cells, such as CD36.Citation34 Additional proteins such as knob-associated histidine-rich protein (KAHRP), mature parasite-infected erythrocyte surface antigen (MESA), PfEMP3, and Pf332, have also been demonstrated to alter mechanical properties of the infected erythrocyte either by anchoring PfEMP1 into the erythrocyte membrane or facilitating its transport to the surface.Citation35-38
Although the surface expression of PfEMP1 has been demonstrated in young gametocytes (stages I and IIa),Citation39 the failure to detect PfEMP1 expression at later gametocyte stages,Citation39-41 and the absence of knobs or appreciable adherence of gametocytes to endothelial cell lines,Citation42 have prompted investigators to explore alternative mechanisms to explain immature gametocyte sequestration. Using a combination of ektacytometry and microsphiltration methods, Tibúrcio et al demonstrated that mature gametocyte-infected erythrocytes are more deformable than immature gametocyte-infected erythrocytes and that this switch in cellular deformability is accompanied by the deassociation of parasite derived STEVOR proteins from the infected erythrocyte membrane.Citation27 In a different approach that utilized a transgenic line for 3D live imaging, in vitro capillary assays and 3D finite element whole cell modeling, Aingarain et al also observed an overall decrease of erythrocyte deformability of immature gametocytes followed by a rapid restoration in mature gametocytes.Citation28 Intriguingly, based on the known dimensions of the splenic endothelial slits (∼2 µm), these modeling efforts also suggested that stage V gametocytes could successfully transit the endothelial slits under physiological pressure (∼10 Pa). Deformability of mature gametocyte-infected erythrocytes has also been linked to reduced cAMP-signaling and increased phosphodiesterase expression in stage V gametocytes, whereas cAMP-dependent parasite kinase activity may contribute to the decreased deformability of immature gametocytes.Citation43
These observations have led to the hypothesis that immature gametocyte enrichment in the extravascular space of the bone marrow and spleen and subsequent release of mature gametocyte into the peripheral circulation is largely mediated by mechanical properties. Furthermore, these studies have established that the persistence of stage V gametocytes in the circulation is ensured by its increased deformability and ability to transit the splenic endothelial slits, thereby escaping splenic entrapment and removal [reviewed in Citation44]. Triggers and mechanisms responsible for the shift in deformability remain largely unknown, however plausible mechanisms to account for the latter, include dismantling of the parasite peripheral cytoskeleton Citation45 and/or interaction of exported parasite proteins with the erythrocyte cytoskeleton.Citation27
Architecture of the cutaneous microvasculature
The microcirculation is defined as the blood flow through arterioles, capillaries and venules which are the smallest vessels in the vasculature and are embedded within organs and tissues. The cutaneous microcirculation is organized as two horizontal plexuses, an upper horizontal network in the papillary dermis that gives rise to the nutritive capillary loops of the dermal papillae, and a lower horizontal plexus in the reticular dermis that is located at the dermal-subcutaneous interface ().Citation46-48 Perforating vessels from the underlying muscles and subcutaneous fat form the lower plexus, giving rise to arterioles and venules that directly connect with the upper horizontal plexus, providing additional lateral tributaries that supply the hair bulbs and sweat glands.
Figure 2. Illustration of the architecture of the cutaneous microcirculation and the fascicle of a biting anopheline mosquito. The vessel diameters of the arterioles, collecting venules, terminal arterioles, and capillary loops are denoted along with the fascicle length and width.
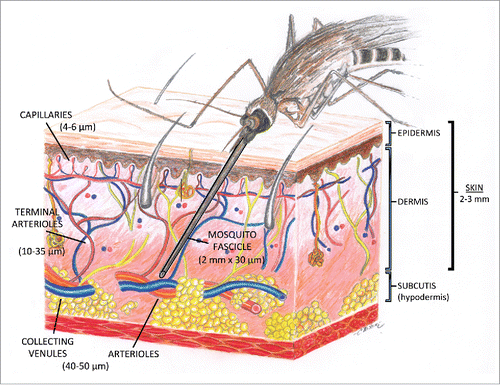
The microvessels in the papillary dermis vary in diameter from 10 to 35 µm, whereas in the reticular dermis, the vessels are twice as large (40 to 50 µm) and represent artertioles and collecting venules.Citation47 The capillary loop arises from a terminal arteriole in the horizontal papillary plexus and is composed of an ascending limb, an intrapapillary loop having a hairpin turn, and a descending limb that connects with a postcapillary venule. The arterioles in the papillary dermis vary from 17 to 26 µm in diameter,Citation49 and shrink in the next contiguous microvascular segment, the arterial capillary, to an outside diameter of 10–12 µm and an internal endothelial tube diameter of 4–6 µm. The venous capillary connects with a postcapillary venule, a vessel whose external diameter increases from 12 to 35 µm and whose endothelial tube diameter enlarges from 8 to 26 µm. There are no ultrastructural differences between the capillary loops at different sites in the skin, including the lower leg; only their density varies.Citation49 Skin has an average thickness of 2–3 mm, varying from 0.5 mm on the eyelids to 4.0 mm on the heels of the feet,Citation50 and contains approximately 20 blood vessels per square inch.
Mechanics of a mosquito bite
Female Anopheline mosquitoes are the vectors responsible for the transmission of malaria. The fascicle, the part of the mosquito used for penetrating the epidermis and drawing blood, is approximately 2 mm in length by 30 µm in diameter. The approximate dimensions of the fasciscle and its comparative size in relationship to the vessels in the cutaneous microvasculature is depicted in . Initial observations of mosquito blood feeding, focused on Aedes aegypti mosquitoes feeding on the leg of a frog or the ear of a mouse.Citation51,52 The Aedes fascicle is much too large to enter the capillaries in the papillary dermis, but capable of penetrating the larger arterioles and collecting venules in the reticular dermis.Citation53
The fascicle is comprised of 6 different stylets which are the labrum, the paired mandibles, the hypopharynx, and the paired maxillae.Citation54 The penetration of the fascicle is achieved by movements of the maxillae, with the maxillary teeth serving as grappling hooks. The fascicle is then pushed deeper into the skin by the action of the right and left protractor and retractors (which are anchored to the skin by its teeth) pulling the mosquito cranium toward the skin surface. Astonishingly, once penetration has been achieved, the tip of the fascicle has been observed to bend through almost a right angle. In a study conducted by Choumet et al that utilized intravital videomicroscopy to analyze the feeding behavior of Anopheles gambiae on anesthetized mice,Citation55 remarkable flexibility of the tip of the labrum during the probing phase of feeding was also observed. This ostensibly allows the mosquito to probe a large area under the skin, without the mosquito having to withdraw the proboscis or change the point of entry. Probing is accompanied by saliva deposition to counteract physiological responses to the arthropod such as hemostasis and inflammation. Contact with a blood vessel is then followed by an engorgement step, which continues until repletion of the mosquito is achieved. Choumet et al, also observed a diversity of blood vessels sizes that were selected for feeding that ranged from blood vessels twice the size of the fascicle to even larger vessels.Citation55 They also observed that mosquitoes infected with the NK65 strain of Plasmodium berghei (rodent malaria) did not differ significantly from uninfected mosquitoes in the time lag until the mosquito began probing or the duration of probing, but that infected mosquitoes were more willing to probe and took longer to achieve repletion than uninfected mosquitoes.
Evidence for the subdermal/dermal sequestration of mature gametocytes
Limited evidence is available to support the hypothesis that mature gametocytes preferentially sequester in the dermal vasculature to facilitate parasite transmission. Considering the critical role of vector parasite interaction at the dermal interface however, lends credence to the hypothesis that they in fact do. Landau et al, have demonstrated preferential localization of infectious P. chabaudi (rodent-malaria) gametocytes in subdermal capillaries in animal models,Citation56,57 whereas trophozoites were found in higher concentrations in larger vessels. Evidence for subdermal/dermal sequestration in humans is supported by a unique study conducted in 1952 that examined 200 study participants for different stages of malaria parasites by skin scarification.Citation58 In this study, a 3-fold higher prevalence of gametocytes was observed in skin samples compared to thick smears prepared from finger-prick samples (which sample deeper vascular beds) from the same area, a finding that was recapitulated in a different setting by the same research group. Evidence for gametocyte clustering in the blood meal may also implicate mechanisms for aggregation of gametocytes in the human host.Citation59 Skin-punch biopsies, not yet attempted to the author's knowledge due to significant ethical and logistic obstacles, produce a cylindrical core of tissue extending through the epidermis, dermis and down into the subcutaneous fat, represent ideal tissue sections that could provide profound insight into the sequestration patterns of gametocytes in the cutaneous microvasculature.
Gametocytes may also respond to circadian rhythms to synch maximal sub-dermal/dermal gametocyte densities with peak hours of mosquito biting. A study of five Gambian infants,Citation21 and more recently, a study of eight Tanzanian children,Citation60 suggest that gametocyte maturation in the peripheral blood may be linked to daily temperature fluctuations in the host. Although peak biting time of the local mosquito vector (00:30–03:30) did not overlap with the highest peripheral density of gametocytes (15:30–19:30), these results do suggest that gametocyte density in the peripheral circulation is a dynamic process that may be governed by host signals. Clearly, additional epidemiological studies are warranted to further investigate these preliminary observations.
Concluding remarks
The fascinating complex interplay between parasite and mosquito vector, and the development of targeted interventions to disrupt parasite transmission is a topic of great interest in the current era of malaria elimination and eradication. Given the constraints of mosquito fascicle length and width, the most likely scenario is that mature infective gametocytes are taken up from 40–50 um arterioles and venules and larger vessels in the retiular dermis at the dermal-subcutaneous junction. Given that subcutaneous vessels are located on average more than 2 mm from the epidermal surface and that the fascicle is ∼2mm in length, it is therefore unlikely that mosquitoes are able to probe into this deeper region. Sequestration of mature gametocytes due to mechanical retention in smaller cutaneous capillaries, their subsequent release into draining larger venules and uptake in a blood meal due to the intense suction exerted by the mosquito pharyngeal pump is a tenable hypothesis that could be probed with further intravital imaging studies. The postulated ability of mature stage V gametocytes to transit ∼2 um endothelial slits in the spleen,Citation28 argues against mechanical retention in 4 um diameter capillaries, and rather for putative adhesin-ligand interactions that could retain mature gametocytes in these larger arterioles and collecting venules.
Skin-punch biopsies, as previously mentioned, could provide insight into the spatial dynamics of gametocyte distribution. An alternative and more palatable approach to punch biopsies includes sampling the dermis with painless microneedles, engineered to mimic a mosquito's proboscis. Microneedles (reviewed in Citation61) have an external diameter of only 60 um (as compared to 900 um for conventional syringes) with a depth of penetration of 3 mm below the skin. These measurements closely mirror the mosquito fascicle and its depth of penetration, and could have utility in painlessly analyzing dermal gametocyte spatial dynamics in epidemiologic studies. Additionally, field studies analyzing the relationship between inherited erythrocyte cytoskeletal disorders characterized by decreased deformability such as Southeast Asian ovalocytosis Citation62,63 and gametocyte carriage rates and/or parasite transmission may yield insight into the mechanisms that mediate the spatial distribution of mature gametocytes in humans. As morphologic analysis indicates that the gametocytes of certain species of murine and rodent plasmodia (P. yoelii & P. chabaudi) also sequester in peripheral capillaries,Citation56,57,64 skin sampling techniques employed in animal models may also prove invaluable in addressing these questions.
Identification of the mechanisms mediating mature gametocyte sequestration and/or their temporal periodicity provides a rational basis for targeted mechanistic studies and the development of new transmission blocking interventions tailored to interrupt contact between the parasite and mosquito vector. A novel class of transmission blocking vaccines that target putative parasite encoded molecules expressed on the gametocyte-infected erythrocyte surface has been put forth by Sutherland.Citation65 Acknowledging the lack of robust experimental data, he hypothesized that this list of putative antigens could include adhesins associated with sequestration and signal receptors responsive to positional and temporal indicators from the host. This approach differs from the leading transmission blocking vaccine candidates, which are based on antibodies generated in the human host that block gamete fertilization/ookinete invasion events in the mosquito luminal mid-gut. Target antigens in this class include Pfs230 (reviewed in Citation66) and Pfs48/45 Citation67-69 in male and female gametocytes and Pfs25 Citation70-71 expressed in zygotes and ookinetes. Pfs25 has undergone extensive evaluation in pre-clinical and phase I clinical trials Citation72 and represents one of the leading target antigens for the development of a transmission blocking vaccine.
Perhaps the answer to gametocyte sequestration in the cutaneous microcirculation lies right before our eyes. The characteristic elliptical banana-shaped morphology of mature gametocytes, one of the most striking features of P. falciparum gametocytes, may passively dictate their impaction in cutaneous capillaries by orienting the gametocytes perpendicularly to the axis of blood flow.Citation73 Whatever the case may be, longitudinal epidemiologic studies, gene profiling, proteomic analysis, and further mechanistic studies are sure to uncover exciting new targetable antigens to assist in the development of an efficacious and deployable malaria transmission blocking vaccine.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
The author gratefully acknowledges critical review of the manuscript by colleagues at the Center for International Health Research, Providence, RI (particularly Christina E. Nixon, PhD).
Funding
CPN is funded by NIGMS Exploratory Grant # 1P20GM104317-01s.
References
- Murray CJ, Rosenfeld LC, Lim SS, Andrew KG, Foreman KJ, Haring D, Fullman N, Naghavi M, Lozano R, Lopez AD. Global malaria mortality between 1980 and 2010: a systematic analysis. Lancet 2012; 379:413-31; PMID:22305225; http://dx.doi.org/10.1016/S0140-6736(12)60034-8
- World Health Organization. World Malaria Report 2014.
- Hay SI, Okiro EA, Gething PW, Patil AP, Tatem AJ, Guerra CA, Snow RW. Estimating the global clinical burden of Plasmodium falciparum malaria in 2007. PLoS Med 2010; 7:e1000290; PMID:20563310; http://dx.doi.org/10.1371/journal.pmed.1000290
- Alonso PL, Brown G, Arevalo-Herrera M, Binka F, Chitnis C, Collins F, Doumbo OK, Greenwood B, Hall BF, Levine MM, et al. A research agenda to underpin malaria eradication. PLoS Med 2011; 8:e1000406; PMID:21311579; http://dx.doi.org/10.1371/journal.pmed.1000406
- Nunes JK, Woods C, Carter T, Raphael T, Morin MJ, Diallo D, Leboulleux D, Jain S, Loucq C, Kaslow DC, et al. Development of a transmission-blocking malaria vaccine: progress, challenges, and the path forward. Vaccine 2014; 32:5531-9; PMID:25077422; http://dx.doi.org/10.1016/j.vaccine.2014.07.030
- Roberts L, Enserink M. Malaria. Did they really say eradication? Science 2007; 318:1544-5; PMID:18063766; http://dx.doi.org/10.1126/science.318.5856.1544
- Malaria Vaccine Funders Group. Malaria Vaccine Technology Roadmap; 2013. Available at: http://www.who.int/immunization/topics/malaria/vaccine_roadmap/TRM_update_nov13.pdf?ua=1
- Gething PW, Elyazar IR, Moyes CL, Smith DL, Battle KE, Guerra CA, Patil AP, Tatem AJ, Howes RE, Myers MF, et al. A long neglected world malaria map: Plasmodium vivax endemicity in 2010. PloS Negl Trop Dis 2012; 6:e1814; PMID:22970336; http://dx.doi.org/10.1371/journal.pntd.0001814
- Baton LA, Ranford-Cartwright LC. Spreading the seeds of million-murdering death: metamorphoses of malaria in the mosquito. Trends Parasitol 2005; 21:573-80; PMID:16236552; http://dx.doi.org/10.1016/j.pt.2005.09.012
- Motard A, Landau I, Nussler A, Grau G, Baccam D, Mazier D, Targett GA. The role of reactive nitrogen intermediates in modulation of gametocyte infectivity of rodent malaria parasites. Parasite Immunol 1993; 15:21-6; PMID:8433851; http://dx.doi.org/10.1111/j.1365-3024.1993.tb00568.x
- Puta C, Manyando C. Enhanced gametocyte production in Fansidar-treated Plasmodium falciparum malaria patients: implications for malaria transmission control programmes. Trop Med Int Health 1997; 2:227-9; PMID:9491100; http://dx.doi.org/10.1046/j.1365-3156.1997.d01-267.x
- Alano P. Plasmodium falciparum gametocytes: still many secrets of a hidden life. Mol Microbiol 2007; 66:291-302; PMID:17784927; http://dx.doi.org/10.1111/j.1365-2958.2007.05904.x
- Smalley ME, Brown J. Plasmodium falciparum gametocytogenesis stimulated by lymphocytes and serum from infected Gambian children. Trans R Soc Trop Med Hyg 1981; 75:316-7; PMID:7029805; http://dx.doi.org/10.1016/0035-9203(81)90348-5
- Babiker HA, Schneider P, Reece SE. Gametocytes: insights gained during a decade of molecular monitoring. Trends Parasitol 2008; 24:525-30; PMID:18801702; http://dx.doi.org/10.1016/j.pt.2008.08.001
- Schneweis S, Maier WA, Seitz HM. Haemolysis of infected erythrocytes-a trigger for formation of Plasmodium falciparum gametocytes? Parasitol Res 1991; 77:458-60; PMID:1891456; http://dx.doi.org/10.1007/BF00931646
- Sokha CS, Trape JF, Robert V. Gametocytemia in Senegalese children with uncomplicated falciparum malaria treated with chloroquine, amodiaquine or sulfadoxine + pyrimethamine. Parasite 2001; 8:243-50; PMID:11584755; http://dx.doi.org/10.1051/parasite/2001083243
- Buckling A, Ranford-Cartwright LC, Miles A, Read AF. Chloroquine increases Plasmodium falciparum gametocytogenesis in vitro. Parasitology 1999; 118:339-46; PMID:10340323; http://dx.doi.org/10.1017/S0031182099003960
- Kafsack BF, Rovira-Graells N, Clark TG, Bancells C, Crowley VM, Campino SG, Williams AE, Drought LG, Kwiatkowski DP, Baker DA, et al. A transcriptional switch underlies commitment to sexual development in malaria parasites. Nature 2014; 131:248-52; http://dx.doi.org/10.1038/nature12920
- Sinha A, Hughes KR, Modrzynska KK, Otto TD, Pfander C, Dickens NJ, Religa AA, Bushell E, Graham AL, Cameron R, et al. A cascade of DNA-binding proteins for sexual commitment and development in Plasmodium. Nature 2014; 13:253-7; http://dx.doi.org/10.1038/nature12970
- Bruce MC, Alano P, Duthrie S, Carter R. Commitment of the malaria parasite Plasmodium falciparum to sexual and asexual development. Parasitology 1990; 100:191-200; PMID:2189114; http://dx.doi.org/10.1017/S0031182000061199
- Hawking F, Wilson ME, Gammage K. Evidence for cyclic development and short-lived maturity in the gametocytes of Plasmodium falciparum. Trans R Soc Trop Med Hyg 1971; 65:549-59; PMID:5003557; http://dx.doi.org/10.1016/0035-9203(71)90036-8
- Thomson JG, Robertson A. The structure and development of Plasmodium falciparum gametocytes in the internal organs and peripheral circulation. Trans R Soc Trop Med Hyg 1935; 29:31-40; http://dx.doi.org/10.1016/S0035-9203(35)90015-3
- Farfour E, Charlotte F, Settegrana C, Miyara M, Buffet P. The extravascular compartment of the bone marrow: A niche for Plasmodium falciparum gametocyte maturation? Malar J 2012; 11:285; PMID:22905863; http://dx.doi.org/10.1186/1475-2875-11-285
- Joice R, Nilsson SK, Montgomery J, Dankwa S, Egan E, Morahan B, Seydel KB, Bertuccini L, Alano P, Williamson KC, et al. Plasmodium falciparum transmission stages accumulate in the human bone marrow. Sci Transl Med 2014; 6:244re5; PMID:25009232; http://dx.doi.org/10.1126/scitranslmed.3008882
- Smalley ME, Abdalla S, Brown J. The distribution of Plasmodium falciparum in the peripheral blood and bone marrow of Gambian children. Trans R Soc Trop Med Hyg 1981; 75:103-5; PMID:7022784; http://dx.doi.org/10.1016/0035-9203(81)90019-5
- Tibúrcio M, Sauerwein R, Lavazec C, Alano P. Erythrocyte remodeling by Plasmodium falciparum gametocytes in the human host interplay. Trends Parasitol 2015; 31:270-8; PMID:25824624; http://dx.doi.org/10.1016/j.pt.2015.02.006
- Tibúrico M, Niang M, Deplaine G, Perrot S, Bischoff E, Ndour PA, Silvestrini F, Khattab A, Milon G, David PH, et al. A switch in infected erythrocyte deformability at the maturation and blood circulation of Plasmodium falciparum transmission stages. Blood 2012; 119:e172-80; PMID:22517905; http://dx.doi.org/10.1182/blood-2012-03-414557
- Aingaran M, Zhang R, Law SK, Peng Z, Undisz A, Meyer E, Diez-Silva M, Burke TA, Spielmann T, Lim CT, et al. Host cell deformability is linked to transmission in the human malaria parasite Plasmodium falciparum. Cell Microbiol 2012; 14:983-93; PMID:22417683; http://dx.doi.org/10.1111/j.1462-5822.2012.01786.x
- Mohandas N, Chasis JA. Red blood cell deformability, membrane material properties and shape: regulation by transmembrane, skeletal and cytosolic proteins and lipids. Semin Hematol 1993; 30:171-92; PMID:8211222
- Beeson JG, Reeder JC, Rogerson SJ, Brown GV. Parasite adhesion and immune evasion in placental malaria. Trends Parasitol 2001; 17:331-7; PMID:11423376; http://dx.doi.org/10.1016/S1471-4922(01)01917-1
- Bhattacharjee S, van Ooij C, Balu B, Adams JH, Haldar K. Maurer's clefts of Plasmodium falciparum are secretory organelles that concentrate virulence protein reporters for delivery to the host erythrocyte. Blood 2008; 111:2418-26; PMID:18057226; http://dx.doi.org/10.1182/blood-2007-09-115279
- Marti M, Good RT, Rug M, Knuepfer E, Cowman AF. Targeting malaria virulence and remodeling proteins to the host erythrocyte. Science 2004; 306:1930-3; PMID:15591202; http://dx.doi.org/10.1126/science.1102452
- Hiller NL, Bhattacharjee S, van Ooij C, et al. A host-targeting signal in virulence proteins reveals a secretome in malarial infection. Science 2004; 306:1934-7; PMID:15591203; http://dx.doi.org/10.1126/science.1102737
- Cooke BM, Mohandas N, Coppel RL. Malaria and the red blood cell membrane. Semin Hematol 2004; 41:173-88; PMID:15071793; http://dx.doi.org/10.1053/j.seminhematol.2004.01.004
- Glenister FK, Coppel RL, Cowman AL, Mohandas N, Cooke BM. Contribution of parasite proteins to altered mechanical properties of malaria-infected red blood cells. Blood 2002; 99:1060-3; PMID:11807013; http://dx.doi.org/10.1182/blood.V99.3.1060
- Crabb BS, Cooke BM, Reeder JC, Waller RF, Caruana SR, Davern KM, Wickham ME, Brown GV, Coppel RL, Cowman AF. Targeted gene disruption shows that knobs enable malaria-infected red cells to cytoadhere under physiological shear stress. Cell 1997; 89:287-96; PMID:9108483; http://dx.doi.org/10.1016/S0092-8674(00)80207-X
- Waller KL, Nunomura W, An X, Cooke BM, Mohandas N, Coppel RL. Mature parasite-infected erythrocyte surface antigen (MESA) of Plasmodium falciparum binds to the 30-kDa domain of protein 4.1 in malaria-infected red blood cells. Blood 2003; 102:1911-4; PMID:12730097; http://dx.doi.org/10.1182/blood-2002-11-3513
- Glenister RK, Fernandez KM, Kats LM, Hanssen E, Mohandas N, Coppel RL, Cooke BM. Functional alteration of red blood cells by a megadalton protein of Plasmodium falciparum. Blood 2009; 113:919-28; PMID:18832660; http://dx.doi.org/10.1182/blood-2008-05-157735
- Hayward RE, Tiwari B, Piper KP, Baruch DI, Day KP. Virulence and transmission success of the malarial parasite Plasmodium falciparum. Proc Natl Acad Sci USA 1999; 96:4563-8; PMID:10200302; http://dx.doi.org/10.1073/pnas.96.8.4563
- Day KP, Hayward RE, Smith D, Culvenor JG. CD36-dependent adhesion and knob expression of the transmission stages of Plasmodium falciparum is stage specific. Mol Biochem Parasitol 1998; 93:167-77; PMID:9662702; http://dx.doi.org/10.1016/S0166-6851(98)00040-1
- Rogers NJ, Daramola O, Targett GA, Hall BS. CD36 and intercellular adhesion molecule 1 mediate adhesion of developing Plasmodium falciparum gametocytes. Infect Immun 1996; 64:1480-3; PMID:8606124
- Silvestrini F, Tibúrcio M, Bertuccini L, Alano P. Differential adhesive properties of sequestered asexual and sexual stages of Plasmodium falciparum on human endothelial cells are tissue independent. PloS One 2012; 7:e31567; PMID:22363675; http://dx.doi.org/10.1371/journal.pone.0031567
- Ramdani G, Naissant B, Thompson E, Breil F, Lorthiois A, Dupuy F, Cummings R, Duffier Y, Corbett Y, Mercereau-Puijalon O, et al. cAMP-signalling regulates gametocyte-infected erythrocyte deformability required for malaria parasite transmission. PloS Pathog 2015; 11:e1004815; PMID:25951195; http://dx.doi.org/10.1371/journal.ppat.1004815
- Dixon MW, Dearnley MK, Hanssen E, Gilberger T, Tilley L. Shape-shifting gametocytes: how and why does P. falciparum go banana-shaped? Trends Parasitol 2012; 28:471-8; PMID:22939181; http://dx.doi.org/10.1016/j.pt.2012.07.007
- Sinden RE. Gametocytogenesis of Plasmodium falciparum in vitro: ultrastructural observations on the lethal action of chloroquine. Ann Trop Med Parasitol 1982; 76:15-23; PMID:7044323; http://dx.doi.org/10.1080/00034983.1982.11687500
- Braverman IM. Ultrastructure and organization of the cutaneous microvasculature in normal and pathologic states. J Invest Dermatol 1989; 93:2S-9S; PMID:2666519; http://dx.doi.org/10.1111/1523-1747.ep12580893
- Braverman IM, Keh-Yen A. Ultrastructure of the human dermal microcirculation: III. The vessels in the mid- and lower dermis and subcutaneous fat. J Invest Dermatol 1981; 77:297-304; PMID:7264363; http://dx.doi.org/10.1111/1523-1747.ep12482470
- Braverman IM, Yen A. Ultrastructure of the human dermal microcirculation: II. The capillary loops of the dermal papillae. J Invest Dermatol 1977; 68:44-52; PMID:830769; http://dx.doi.org/10.1111/1523-1747.ep12485165
- Yen A, Braverman IM. Ultrastructure of the human dermal microcirculation: the horizontal plexus of the papillary dermis. J Invest Dermatol 1976; 66:131-42; PMID:1249441; http://dx.doi.org/10.1111/1523-1747.ep12481678
- Histology Guide, Faculty of Biological Sciences, University of Leeds. Available at http://www.histology.leeds.ac.uk/skin/skin_layers.php
- Gordon RM, Lumsden WHR. A study of the behavior of the mouth-parts of mosquitoes when taking up blood from living tissue; together with some observations on the ingestion of microfilariae. Ann Trop Med Parasit 1939; 33:259-78; http://dx.doi.org/10.1080/00034983.1939.11685071
- Robinson GG. The mouthparts and their function in the female mosquito, Anopheles maculipennis. Parasitology 1939; 31:212-42; http://dx.doi.org/10.1017/S0031182000012774
- Swaminathan VS. Mechanics of a Mosquito Bite (Thesis submitted to the North Carolina State University, Raleigh NC). 2006.
- Clements AN. “The biology of mosquitoes” Volume 1. Chapman and Hall.
- Choumet V, Attout T, Chartier L, Khun J, et al. Visualizing non infectious and infectious Anopheles gambiae blood feedings in naïve and saliva-immunized mice. PLoS One 2012; 7:e50464; PMID:23272060; http://dx.doi.org/10.1371/journal.pone.0050464
- Gautret P, Miltgen F, Gantier JC, Chabaud AG, Landau I. Enhanced gametocyte formation by Plasmodium chabaudi in immature erythrocytes: pattern of production, sequestration, and infectivity to mosquitoes. J Parasitol 1996; 82:900-6; PMID:8973397; http://dx.doi.org/10.2307/3284196
- Landau I, Chabaud A. Parasitic pattern of rodent Plasmodium in blood from mouse tail and from Anopheles blood meal. Parassitologia 2002; 44:111-5; PMID:12404818
- Chardome M, Janssen PJ. Enquête sur l'incidence malarienne par la method dermique dans la regionde Lubilash (Congo Belge). Ann Soc Belge Med Trop 1952; 32:209-11; PMID:12976890
- Pichon G, Awono-Ambene HP, Robert V. High heterogeneity in the number of Plasmodium falciparum gametocytes in the bloodmeal of mosquitoes fed on the same host. Parasitology 2000; 121:115-20; PMID:11085230; http://dx.doi.org/10.1017/S0031182099006277
- Magesa SM, Mdira YK, Akida JA, Bygbjerg IC, Jakobsen PH. Observations on the periodicity of Plasmodium falciparum gametocytes in natural human infections. Acta Trop 2000; 76:239-46; PMID:10974164; http://dx.doi.org/10.1016/S0001-706X(00)00110-8
- Prausnitz MR. Microneedles for transdermal drug delivery. Adv Drug Deliv Rev 2004; 56:581-7; PMID:15019747; http://dx.doi.org/10.1016/j.addr.2003.10.023
- Jarolim P, Palek J, Amato D, Hassan K, Sapak P, Nurse GT, Rubin HL, Zhai S, Sahr KE, Liu SC. Deletion in erythrocyte band 3 gene in malaria-resistant Southeast Asian ovalocytosis. Proc Natl Acad Sci USA 1991; 88:11022-6; PMID:1722314; http://dx.doi.org/10.1073/pnas.88.24.11022
- Mohandas N, Lie-Injo LE, Friedman M, Mak JW. Rigid membranes of Malayan Ovalocytes: A likely genetic barrier against malaria. Blood 1984; 63:1385-92; PMID:6722355
- Landau I, Miltgen F, Boulard Y, Chabaud AG, Baccam D. Etudes sur les gamétocytes des Plasmodium du groupe “vivax:” Morphologie, évolution, prise par les Anophèles et infectivité des microgamétocytes de Plasmodium yoelii. Annales de Parasitologie Humaine et Comparée 1979; 54:145-61.
- Sutherland CJ. Surface antigens of Plasmodium falciparum gametocytes – A new class of transmission-blocking vaccine targets? Mol Biochem Parasitol 2009; 166:93-8; PMID:19450726; http://dx.doi.org/10.1016/j.molbiopara.2009.03.007
- Williamson KC. Pfs230: from malaria transmission-blocking vaccine candidate toward function. Parasite Immunol 2003; 25:351-9; PMID:14521577; http://dx.doi.org/10.1046/j.1365-3024.2003.00643.x
- Jones S, Grignard L, Nebie I, Chilongola J, Dodoo D, Sauerwein R, Theisen M, Roeffen W, Singh SK, Singh RK, et al. Naturally acquired antibody responses to recombinant Pfs230 and Pfs48/45 transmission blocking vaccine candidates. J Infect 2015; 71:117-27; PMID:25869538; http://dx.doi.org/10.1016/j.jinf.2015.03.007
- Singh SK, Roeffen W, Andersen G, Bousema T, Christiansen M, Sauerwein R, Theisen M. A plasmodium falciparum 48/45 single epitope R0.6C subunit protein elicits high levels of transmission blocking antibodies. Vaccine 2015; 33:1981-6; PMID:25728318; http://dx.doi.org/10.1016/j.vaccine.2015.02.040
- Ouédraogo AL, Roeffen W, Luty AJ, de Vlas SJ, Nebie I, Ilboudo-Sanogo E, Cuzin-Ouattara N, Teleen K, Tiono AB, Sirima SB, et al. Naturally acquired immune responses to Plasmodium falciparum sexual stage antigens Pfs48/45 and Pfs230 in an area of seasonal transmission. Infect Immun 2011; 79:4957-64; http://dx.doi.org/10.1128/IAI.05288-11
- Kumar R, Ray PC, Datta D, Bansal GP, Angov E, Kumar N. Nanovaccines for malaria using Plasmodium falciparum antigen Pfs25 attached to gold nanoparticles. Vaccine 2015; 33:5064-71; PMID:26299750; http://dx.doi.org/10.1016/j.vaccine.2015.08.025
- Kumar R, Angov E, Kumar N. Potent malaria transmission-blocking antibody responses elicited by Plasmodium falciparum Pfs25 expressed in Escherichia coli after successful protein refolding. Infect Immun 2014; 82:1453-9; PMID:24421036; http://dx.doi.org/10.1128/IAI.01438-13
- Wu Y, Ellis RD, Shaffer D, Fontes E, Malkin EM, Mahanty S, Fay MP, Narum D, Rausch K, Miles AP, et al. Phase 1 trial of malaria transmission blocking vaccine candidate Pfs25 and Pvs25 formulated with montanide ISA 51. PLoS One 2008; 3:e2636; PMID:18612426; http://dx.doi.org/10.1371/journal.pone.0002636
- Nacher M. Does the shape of Plasmodium falciparum gametocytes have a function? Med Hypotheses 2004; 62:618-9; PMID:15050117; http://dx.doi.org/10.1016/j.mehy.2003.11.011