
ABSTRACT
An HIV-1 vaccine is needed to curtail the HIV epidemic. Only one (RV144) out of the 6 HIV-1 vaccine efficacy trials performed showed efficacy. A potential mechanism of protection is the induction of functional antibodies to V1V2 region of HIV envelope. The 2 main current approaches to the generation of protective immunity are through broadly neutralizing antibodies (bnAb) and induction of functional antibodies (non-neutralizing Abs with other potential anti-viral functions). Passive immunization using bnAb has advanced into phase II clinical trials. The induction of bnAb using mimics of the natural Env trimer or B-cell lineage vaccine design is still in pre-clinical phase. An attempt at optimization of protective functional antibodies will be assessed next with the efficacy trial (HVTN702) about to start. With on-going optimization of prime/boost strategies, the development of mosaic immunogens, replication competent vectors, and emergence of new strategies designed to induce bnAb, the prospects for a preventive HIV vaccine have never been more promising.
Introduction
HIV infection is a major global health issue, affecting 36.7 million people world-wide.Citation1 New infections continue to occur, with 2.1 million cases in 2015.Citation1 The number of people living with HIV on antiretroviral therapy (ART) reached 17 million in 2015.Citation1 ART has dramatically reduced morbidity and mortality in individuals with HIV infectionCitation2 and can also prevent HIV transmission.Citation3-5 However, it cannot eradicate HIV infection due to the persistence of a latent viral reservoir (mean half-life of 44 months).Citation6,7 Thus, the need for ART is lifelong and the cost is substantialCitation8,9 and may be difficult to sustain economically.Citation10 Although ART is highly efficacious in preventing transmission in the setting of mother to child transmission,Citation11 in sexual transmission through the treatment of infected partners in serodiscordant relationships,Citation12 through pre-exposure,Citation4,13,14 or post-exposure prophylaxisCitation15 scale-up difficulties and costs may make widespread implementation challenging. Furthermore, adherence is critical to the efficacy of biomedical preventive interventions but has been varied across study populations.Citation16 According to Fauci and Marston, “even if HIV prevention efforts were optimally implemented to achieve a new infection rate of near zero, recidivism could threaten this success.” Thus an HIV vaccine is essential as it is a more sustainable solution.Citation17 Modeling data suggest that a 70% efficacious vaccine introduce in 2027 with strong uptake and 5 y of protection could reduce annual new infections by 44% over the first decade and by 78% in 2070.Citation18 Therefore, an effective universal prophylactic vaccine can potentially curtail and end the worldwide HIV pandemic.
The development of a universal effective HIV vaccine is an exceptionally difficult biomedical challenge. Firstly, no case of natural eradication of HIV infection has been identified, thus causal mechanisms of protection have not been definitively established.Citation19-21 Therefore, immune responses induced by HIV infection may not be effective in preventing HIV infection. Secondly, the extreme diversity of HIV is a major obstacle as strains belonging to different subtypes can differ by up to 35% in their envelope (Env) proteins.Citation22,23 Thus, vaccine immunogens derived from a particular clade may not be effective against other clades. To generate an efficacious global vaccine, immunogens capable of generating protective responses covering most major strains are required.
This review intends to summarize data from the 6 HIV-1 vaccine efficacy trials done to date (), to delineate potential protective responses, and to explore new vaccine candidates that are currently being developed.
Table 1. Summary of HIV-1 vaccine efficacy trials.
Lessons from efficacy trials and correlates of risk
Most of the licensed vaccines against both bacterial and viral infections, including diphtheria, tetanus, pertussis, Haemophilus Influenzae Type b, pneumococcus, hepatitis A, hepatitis B, varicella, measles, rubella, polio, and influenza, prevention of infection correlates with the induction of antibodies.Citation24,25 Furthermore, pilot studies of recombinant HIV-1 Env glycoprotein subunit (rgp120) vaccines conferred protection of chimpanzees from intravenous and mucosal challenge with homologous and heterologous HIV-1 strains.Citation26-28 Therefore, initial HIV-1 vaccine approaches (VAX003 and VAX004) focused primarily on the generation of neutralizing antibodies (nAb).
VAX003 and 004
VAX003 was a double-blind, randomized trial of AIDSVAX® B/E (a bivalent vaccine composed of rgp120 from subtype B, strain MN and subtype CRF01_AE, strain A244) in injection drug users (IDU) in Thailand.Citation29 VAX004 was a double-blind, randomized trial of AIDSVAX® B/B (a bivalent vaccine composed of subtype B rgp120 from strains MN and GNE8) conducted among men who have sex with men (MSM) and women at high risk for heterosexual transmission of HIV-1 in North America and The Netherlands.Citation30 Despite the development of anti-gp120 antibody responses, both vaccines did not demonstrate protection. Correlates of risk analysis found that higher nAb to HIV-1MN, CD4 blocking Ab and antibody-dependent, cell-mediated viral inhibition (ADCVI) were associated with reduced infection rates among vaccine recipients in VAX004.Citation31,32
Given the disappointing results from the VAX003 and VAX004 trials and data supporting the importance of cell mediated immunity in controlling viral replication in rhesus macaques (RM)Citation33-35 and human elite controllers,Citation36-38 attention turned to the use of T-cell vaccines to induce HIV-specific cellular immune responses.
STEP and phambili studies
The STEP study was a double-blind, randomized trial of the MRKAd5 HIV-1 gag/pol/nef sub-type B vaccine in individuals at high risk of HIV-1 acquisition in the Americas, Caribbean and Australia.Citation39 The vaccine consisted of a 1:1:1 mixture of 3 separate replication-defective adenovirus (Ad) 5 vectors, each expressing the gag gene from HIV-1 strain CAM-1, the pol gene from HIV-1 strain IIIB, and the nef gene from HIV-1 strain JR-FL. Despite eliciting IFN-γ ELISPOT responses in 75% of vaccinees, the vaccine did not prevent HIV-1 infection and had no effect on plasma viral load. Instead, it was associated with an increased incidence of HIV-1 acquisition in male vaccinees who were Ad5 seropositive pre-vaccination or were uncircumcised. Thus, the trial and was stopped after the first interim analysis.Citation39
Subsequent comparative analyses between cases with HIV-1 infection and non-cases did not reveal differences in HIV-specific immunologic responses.Citation40 However, vaccine-induced T-cell responses exerting selective pressure on breakthrough viruses was evident in sieve analyses.Citation41
The Phambili study was a double-blind, randomized trial designed to evaluate the MRKAd5 HIV-1 gag/pol/nef sub-type B vaccine in individuals in South Africa where HIV clade C is predominant. This study was halted following the Step study's interim analysis and subsequent analysis also found no efficacy.Citation42
RV144
The next efficacy trial to occur was the RV144 trial, a randomized, double-blind trial that evaluated 4 priming injections of ALVAC-HIV [vCP1521], recombinant canarypox vector expressing HIV-1 Gag and Pro (subtype B LAI strain) and CRF01_AE (subtype E) HIV-1 gp120 (92TH023) linked to the transmembrane anchoring portion of gp41 (LAI) plus 2 booster injections, AIDSVAX® B/E (bivalent HIV-1 gp120 subunit vaccine containing a subtype E Env from strain A244 (CM244) and a subtype B Env from strain MN), co-formulated with alum.Citation43 The rationale for the prime boost strategy was to induce both cellular and humoral responses.Citation44,45 The RV144 trial was the only efficacy trial to date that demonstrated efficacy, 60% at 12 months (post hoc analysis)Citation46 that declined to 31% at 3.5 y (modified intention-to-treat analysis).Citation43
The finding of vaccine efficacy in RV144, despite the induction of only weakly nAb was paradigm changing.Citation47 The investigation of correlates of risk using specimens from week 26 (2 weeks post last vaccination) from 41 cases of vaccine recipients who acquired HIV-1 and 205 control (vaccine recipients who did not acquire HIV-1) identified that the development of non-neutralizing, binding IgG to scaffolded gp70-variable regions 1 and 2 (V1V2) of HIV-1 Env proteins was inversely correlated with infection and Env-specific IgA was directly correlated with infection. It is important to bear in mind that neither low levels of IgG to V1V2 nor high levels of Env-specific IgA in vaccinees were associated with higher rates of infection than placebo recipients. Thus, there was no vaccine-associated enhancement of infection.Citation48,49 Therefore, these results suggest that IgG to V1V2 may have contributed to protection against HIV-1 infection. In the setting of low vaccine induced Env IgA, avidity of IgG for Env, antibody-dependent cellular cytotoxicity (ADCC), nAb and Env-specific CD4+ T cells were inversely correlated with the risk of infection.Citation48
Subsequently a number of studies further exploring the potential mechanisms of protection of RV144 have been performed.Citation50-52 Sieve analysis comparing sequences of breakthrough virus isolates found that isolates from vaccine recipients were less likely to have a lysine at K169 of the Env V2 region than placebo recipients, suggesting selective effects of vaccine induced immune responses on V1V2 on breakthrough HIV-1 viruses.Citation53 Further characterization of RV144 vaccine-induced V2 Ab found that the Ab recognized both conformational (on gp70-V1V2) and linear epitopes (residues 165–178, immediately N-terminal to the putative α4β7 binding motif in the mid-loop region of V2) and were cross-reactive with the V1V2 regions from many HIV-1 subtypes (A, B, C and CRF01_AE).Citation54 In addition, epitope mapping of IgG to V1V2 indicated binding also involved the lysine residue at amino acid 169 in the Env V2 region. The V2 Ab isolated from RV144 vaccinees were able to mediate ADCC against RV144 breakthrough Env-target cells, and this ADCC activity was dependent on position 169 in breakthrough Envs.Citation55
Though AIDSVAX® B/E was part of the vaccine regimen in both RV144 and VAX003, only RV144 showed efficacy. Overall, both vaccines predominantly elicited gp120 specific IgG1Ab. However, compared with VAX003, RV144 induced more gp120-specific IgG3, which was associated with enhance Ab mediated effector functions, including ADCC and Ab-dependent cellular phagocytosis (ADCP).Citation56,57 In comparison to VAX003, RV144 elicited significantly higher IgG3 to V1V2, which was also correlated with lower HIV-1 infection risk.Citation58 Furthermore, when compared with VAX003 and VAX004, complement activation by V1V2 Ab, measured as C3d deposition on a panel of gp70-V1V2-coated beads was stronger and detected more frequently in RV144. Positive V1V2-specific complement activating Ab was also correlated with lower infection risk in RV144 vaccine recipients.Citation59
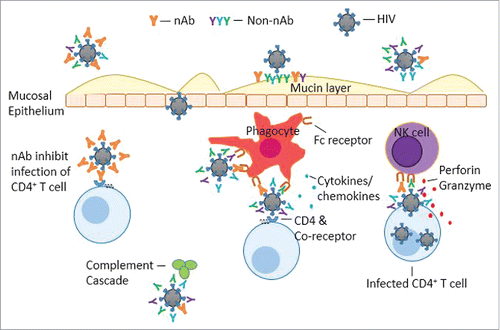
Therefore, though only weakly neutralizing, Ab induced by RV144 may potentially mediate protection through aggregating or immobilizing virions in mucin layers, impeding transverse through mucosal barriers,Citation60-62 via IgG Fcγ receptor (FcγR) dependent ADCVI including ADCC,Citation55-57 ADCP,Citation56,57 Ab-mediated release of cytokines or chemokines and complement-mediated killing.Citation50-52,59,63,64 (). These non or weakly-neutralizing Ab with other potential anti-viral functions will subsequently be referred to as “functional Ab.”
Figure 1. Schematic representation of potential mechanisms of protection by vaccine induced antibodies. Broadly neutralizing antibodies (BnAb) can bind to HIV virions from diverse strains, inactivating the virions. However, bnAb may not completely block mucosal portal of entry. Beyond neutralization, Abs may mediate protection through binding to cognate epitopes on HIV, immobilizing virions in mucin layers. Ab can also elicit immune responses through complement mediated lysis as well as recognition by FC receptors on effector cells (Natural Killer, NK cells and phagocytes) and mediate antibody dependent cell-mediated viral inhibition (ADCVI), including antibody dependent cytotoxicity (ADCC), Ab-dependent cellular phagocytosis (ADCP) and Ab-mediated release of cytokines or chemokines.
The association between Env-specific IgA antibodies and lack of protection was perplexing at firstCitation51 Subsequent analysis found that there was significant enrichment of higher Env IgA/IgG ratio among infected vs uninfected vaccinees. Furthermore, some Env-specific IgA from RV144 vaccinees were able to block the binding of an IgG monoclonal Ab that mediated ADCC. In addition, an Env-specific IgA monoclonal Ab isolated from an RV144 vaccinee also inhibited ADCC, measured as the ability of natural killer cells to kill HIV-1-infected CD4+ T cells coated with RV144-induced IgG. Thus, these data support the hypothesis that higher IgA may modulate vaccine-induced immunity by diminishing ADCC effector function.Citation65
HVTN 505
The last efficacy trial conducted to date is the HVTN 505 trial, a randomized, placebo-controlled trial of a prime boost, DNA/rAd5 vaccine consisting of a 6-plasmid DNA vaccine (expressing clade B Gag, Pol, and Nef, and Env proteins from clades A, B, and C) with rAd5 vector boost (expressing clade B Gag-Pol fusion protein and Env glycoproteins from clades A, B, and C).Citation66 The trial was halted prematurely due to lack of efficacy. The vaccine induced both cellular and humoral responses. However, these were not associated with protection.Citation66
In summary, none of the vaccine candidates that have completed efficacy trials to date induced strong bnAb responses. CD8+ T cell responses were induced in STEP, Phambili and HVTN505 studies but were not associated with protection. Only one trial, RV144 demonstrated efficacy and protection was associated with functional binding antibodies. However, efficacy was of suboptimal magnitude and was not durable.
The way forward
The current major goal in HIV vaccine design is to elicit a protective immune response mediated primarily by antibodies that are able to recognize a range of diverse strains. One approach is focused on the induction of broadly neutralizing antibodies (bnAb) with activities against major strains that are common in human transmission. Another approach is the induction of protective functional antibodies as well as T cell responses through prime boost strategies.Citation67-69 A number of HIV-1 vaccines at different stages of development are currently in the pipeline ().
Table 2. HIV Vaccine Pipeline.
Broadly neutralizing antibodies
A detailed review of bnAb in HIV infection is beyond the scope of this review. Excellent reviews on this topic have been recently published.Citation70-72 BnAb, antibodies capable of neutralizing diverse circulating strains from multiple clade groups, can be present in 20–30% of individuals with HIV-1 infection.Citation73-77 They usually develop 2–4 y after HIV-1 infection, in the presence of continual antigen stimulation from viral replication.Citation75,77,78 HIV Env protein, composed of 3 gp120 and 3 gp41 monomers, is the main target for bnAb.Citation71,79 BnAb can be directed at the CD4-binding site on gp120, the glycopeptides on V1V2 of gp120, the glycans on V3 region, the membrane proximal external region (MPER) on gp41, the contiguous region of gp120-gp41 and the glycan shield.Citation71,79
Passive immunization using broadly neutralizing antibodies
The efficacy of bnAb as passive immunotherapy has been demonstrated in RM models. A single infusion of broadly neutralizing antibody (PGT121,Citation80,81 VRC01,Citation81-83 10E8Citation83 and 3BNC11781) can prevent infection from a single high-dose Simian/Human Immunodeficiency Virus (SHIV) challenge. A single infusion of VRC01, 3BNC117, and/or 10–1074 has also been shown to delay SHIV acquisition after repeated, low dose, weekly intrarectal challenges, for up to 23 challenges.Citation84
The site of antibody interception of SHIV and the mechanisms by which bnAb mediates protection have not been clearly elucidated. In a recent study by Liu et al., the protective efficacy of PGT121 against intravaginal challenge with SHIV-SF162P3 was first established in 12 female RM, with no detectable plasma SHIV RNA for over 6 months following challenge. In another group of 24 RM given the same bnAb and challenge, low levels of SHIV RNA were detectable in at least one tissue distal to the site of infection, in 7/8 monkeys necropsied at day 1, 3 and 7 post challenge. SHIV RNA and DNA were not detectable in all tissues sampled at the day 10 necropsy. Viral clearance was not associated with SHIV-specific T cell responses. Instead, it seemed to be associated with the activation of innate immune responses. These data demonstrated that PGT121 did not completely block the challenge virus at the mucosal portal of entry. Instead, some virions appeared to transit to distal tissues, but seemed to be vulnerable to immune-mediated elimination as they were progressively cleared over a period of ∼7 days.Citation85
VRC01, a bnAb that binds HIV-1 at the gp120 CD4 binding site, is currently the bnAb that has the most human passive immunotherapy data. A recent phase I study in healthy adults found that VRC01 administered intravenously or subcutaneously was safe and well tolerated.Citation86 The AMP (antibody mediated prevention) study, consisting of 2 phase II protocols, aims to evaluate the safety and efficacy of the VRC01 in reducing HIV-1 acquisition, among 2700 men and transgender persons who have sex with men in the US and South America (HVTN704/HPTN 085, clinicaltrials.gov NCT02716675) and in 1500 women in Sub-Saharan Africa (HVTN 703/HPTN 081, clinicaltrials.gov NCT02568215), is currently underway ().
The use of bnAb as passive immunotherapy in its current form will be challenging to implement widely, due to the production costs, the healthcare infrastructures necessary for infusions and the need for repeated administrations. Thus, new research is taking place to explore the introduction of bnAb using vectored immunoprophylaxis, where adeno-associated virus (AAV) vectors are used to deliver the genes encoding bnAb to muscle tissues, thereby enabling long-term production and systemic distribution. This technique has been shown to protect humanized mice as well as RM against high dose intravenous and repeated mucosal challenges.Citation87-89 The first human trial, IAVI A003 (clinicaltrials.gov NCT01937455) a phase I, randomized, blinded, dose-escalation study of rAAV vector coding for PG9 Ab in 24 healthy men is close to completion ().
Administration of Ab that are not broadly neutralizing may also mediate protection. α4β7 integrin monoclonal antibody (mAb) binds strongly to memory CD4+ T cells (α4β7hi), selectively inhibits interactions with MAdCAM-1 on endothelial cells, thereby interferes with the trafficking of lymphocytes to the gut. In RM models, administration of α4β7 integrin mAb to uninfected RM prevented or delayed infection after low dose vaginal simian immunodeficiency virus (SIV) challenge.Citation90 More recently, infusion of α4β7 integrin mAb, in RM SIV models, was associated with sustained control of plasma viremia even months after ART and α4β7 integrin mAb were discontinued.Citation91 The mechanism for persistent virologic control remains to be defined, but was not associated with nAb or classical cell mediated immune responses. This major breakthrough show that antibody administration has the potential to modulate anti-HIV immune responses and has implications for both HIV-1 prevention and cure.
Eliciting broadly neutralizing antibodies through immunization
The promising results from bnAb as passive immunotherapy in animal models have spurred great interest in the design of HIV-1 vaccines capable of inducing bnAb.Citation70,92 Though a number of bnAb and their target epitopes have already been identified (discussed above), generating bnAb against HIV-1 is no simple endeavor as the mechanisms for the development of bnAb have not been clearly elucidated.Citation92 An immunogen that can elicit bnAb responses has still not been identified and the high levels of somatic mutations in bnAb suggest complex maturation pathways.Citation93-95 The SOSIP gp140 trimer is a mimic of the natural Env trimer, where the gp120-gp41 interactions are stabilized by an intermolecular disulfide bond, and the gp41-gp41 interactions are stabilized by an isoleucine-to-proline substitution at position 559 in the N-terminal heptad repeat region of gp41.Citation96 It can be further modified to express epitopes of bnAb and to react with bnAb.Citation97 Immunization with SOSIP trimers induced nAb in rabbits and to a lesser extent in RM but bnAb responses were not generated.Citation98 The next step that has been proposed is the use of a longitudinal series of SOSIP trimers from multiple variants to induce bnAb.Citation70,98
Recently, computational redesign in the HR1 (a region that undergoes drastic conformational change during viral fusion with host cells) of SOSIP trimers has led to improved yield, purity and stability of trimers that closely mimic the native, pre-fusion trimer.Citation99 Furthermore, these trimers, presented on nanoparticles were able to bind bnAb and trigger B cells carrying cognate VRC01 B cell receptors.Citation100 These gp140-nanoparticles can be manufactured in CHO cell systems thus scalability is possible.Citation100
An alternative pathway to elicit bnAb is B-cell lineage vaccine design.Citation101 Unmutated common ancestors, the putative naïve B cell receptors of bnAb could be targeted with relevant priming Env immunogens to trigger affinity maturation, followed by boosting to develop breadth. A germline-targeting, gp120 outer domain immunogen (OD-GT8) on self-assembling nanoparticles, was able to activate VRC01-class precursors, select productive mutations, create a pool of memory phenotype B cells and induced antibodies that showed characteristics of the VRC01-class bnAb, in a germline reverted VRC01 H-chain knock-in mouse model (VRCO1 gH)Citation102; as well as isolate VRC01-class precursor naïve B cells from HIV-uninfected donors.Citation103 Furthermore, administration of boosting immunogens (BG505 core-GT3 nanoparticle (NP) and BG505 SOSIP-GT3 trimer) drove the maturation of eOD-GT8 60-mer primed B cells toward VRC01-class bnAb and induced broad neutralization of near-native (N276A) viruses and weak neutralization of a fully native virus in VRC01 gH mice.Citation104 These data showed that B-cell lineage vaccine design has the potential to induce bnAb.
Eliciting bnAb through vaccination is still at the very early stages of development and is unlikely to progress to efficacy trial in the near future. On the other hand, more evidence and experience exist for the induction of functional antibodies and T cell responses through prime boost strategies. New data and insights are emerging from studies that build on the success and refinement RV144, on the use of mosaic antigens and the development of new vectors.
Studies building on RV144
The Pox-Protein Public-Private Partnership (P5) is a diverse group of organizations committed to building on the success of RV144, with the goal to deepen the understanding of immune responses associated with HIV prevention and to produce a licensable HIV-1 vaccine, with efficacy of 50% for 3 years, that can result in significant impact in Southern Africa. Two clinical trials, RV305 (clinicaltrials.gov NCT01435135) and RV306 (clinicaltrials.gov NCT01931358) were designed to evaluate whether additional boosting with components of RV144 can increase the durability of immune responses induced by RV 144 ().
In RV305, HIV-uninfected RV144 volunteers (n = 162) who completed all vaccinations 6–8 y earlier, were randomized to receive 2 injections of ALVAC-HIV or AIDSVAX® B/E or both, or placebo, at day 0 and 6 months. The original RV305 study activities are completed, and analyses are well underway. Weak residual HIV-specific ab responses from RV144 were present at the baseline, and rose dramatically after additional boosting with AIDSVAX® B/E. Responses declined 6 months post first boost, but increased again, albeit to lower levels following the second boost. IgG responses against gp120 and scaffolded gp70-V1V2 at weeks 2 and 26 in the groups receiving ALVAC-HIV/AIDSVAX® B/E or AIDSVAX® B/E alone were significantly higher than levels at peak immunogenicity in RV144. There were no significant differences between groups receiving ALVAC-HIV/AIDSVAX® B/E vs AIDSVAX® B/E alone. ALVAC-HIV alone did not induce significant titers against any of the capture antigens. These data suggest that late boosting with AIDSVAX® B/E with or without ALVAC-HIV may overcome suboptimal efficacy induced by the RV144 regimen. Unfortunately, RV305 vaccinations did not improve the durability of Ab responses when compared with RV144.Citation105 IgG responses to HIV-1 gp120 and gp70V1V2 scaffolds were also induced in cervico-vaginal mucous and seminal plasma after boosting with AIDSVAX® B/E with/without ALVAC-HIV. IgG responses were not detectable in anogenital secretions from the group receiving ALVAC-HIV alone and were also not detectable in rectal secretion from any groups. IgA responses were absent in anogenital secretions from all groups.Citation106
RV306 recapitulated the original RV144 regimen in 360 vaccine naïve participants with an additional late boost of AIDSVAX® B/E at week 48 or ALVAC-HIV/AIDSVAX® B/E at week 48, or 60 or 72. The aim was to provide more in depth characterization of vaccine induced innate, cell-mediated and humoral immune responses in the systemic as well as mucosal compartments. Clinical activities are completed, and analyses have begun. Preliminary results showed that IgG to gp120 and gp70V1V2 scaffolds declined significantly at week 50 if no additional boost was given. Additional boost at week 48 maintained IgG to cognate gp120 proteins at levels similar to the RV144 series at week 26, and significantly increased IgG to gp70V1V2 scaffolds. Responses were similar between groups receiving AIDSVAX® B/E, with/without ALVAC-HIV.Citation107 Tier 1 nAb responses were also improved in all groups that received additional boost and was higher with boost at week 60 and 72 when compared with boost at week 48.Citation108 Furthermore, IgG to gp70V1V2 CRF01_AE scaffolds and IgG to HIV-1 gp120 was detected in all anogenital secretions at peak immunogenicity (week 26), with the highest IgG levels to HIV-1 gp120 in cervico-vaginal mucus followed by seminal plasma and rectal secretions.Citation109
Therefore, preliminary data from RV305 and RV306 suggest that additional boosting could potentially improve efficacy of RV144. Furthermore, vaccine-induced Ab may play a role in mediating protection in anogenital mucosa, the sites of initial viral entry ().
HVTN 100 (clinicaltrials.gov NCT02404311, currently ongoing) is a phase I/II randomized, double blind, placebo-controlled trial using a prime-boost regimen that is a variant of RV144, based on clade C HIV-1 with ALVAC-HIV [vCP2438] prime (expressing clade C gp120 and clade B gp41, gag and protease) and bivalent subtype C gp120 boost, with MF59® (water-in-oil emulsion) instead of alum as the adjuvant, in HIV-seronegative low risk adults in South Africa. This vaccine was designed to increase subtype C coverage, improve antibody durability and to investigate whether IgG to V1V2 is a correlate of protection. Preliminary data presented at AIDS 2016 showed that the vaccine has passed all predetermined criteria for moving on to phase III efficacy study (HVTN 702), namely i) prevalence of IgG binding antibodies to at least 2 of 3 gp120 vaccine antigens ≥ 75%; ii) non-inferior IgG binding antibody magnitude to gp120 vaccine antigens when compared with RV144; iii) non-inferior response rate of Env-specific CD4+ T cells expressing IL-2, IFN-Υ or CD40L when compared with RV144; and iv) prevalence of IgG binding antibodies to at least 1 clade C V1V2 Env ≥ 56%.Citation110 HVTN 702 (NCT02968849), aiming to assess efficacy and safety of ALVAC [vCP2438] prime, gp120 with MF59® boost in 5400 participants in South Africa is currently enrolling ().
Mosaic vaccine
All the HIV-1 vaccines that have progressed to efficacy trials to date have predominantly been regional and clade-specific. The goal of mosaic HIV-1 vaccine is to generate immune responses that cover the diverse spectrum of circulating HIV-1 isolates, potentially resulting in a single vaccine that can be rolled out globally. Polyvalent mosaic immunogens are generated from natural sequences via computational optimization so that they resemble natural proteins but systematically include common potential epitopes, providing diversity coverage comparable to that afforded by thousands of separate peptides.Citation111
Mosaic HIV-1 antigens delivered by replication-incompetent Ad26 vectorsCitation112 or DNA prime-recombinant vaccinia boost regimensCitation113 have been shown to augment both the breadth and depth of antigen-specific T cell responses when compared with consensus or natural sequence HIV-1 antigens in RM. In RM, Ad/MVA (modified vaccinia Ankara) or Ad/Ad prime boost vectors expressing bivalent HIV-1 mosaic Env/Gag/Pol elicited neutralizing, and functional non-neutralizing Ab, robust T cell immune response and afforded significant reduction in the per-exposure acquisition risk following repetitive, intrarectal SHIV challenges.Citation114 Furthermore, Ad26 mosaic prime and gp140 boost was associated with 40% protection after 6 intrarectal challenges in RM.Citation115 HIV-V-A002 (NCT02218125), a phase I study of MVA mosaic vaccine in 25 participants in the USA has recently been completed. HIV-V-A004 (NCT02315703), an international phase I study using Ad26 mosaic vectors prime, followed by boosting with Ad26 or MVA mosaic vectors and/or high, low or no clade C gp140 in 394 volunteers is at the final data collection stage (). Results from these studies will inform the optimization of mosaic vaccine regimens that may advance into efficacy trials in 2017.
New vectors
Vectors are critical elements of a successful vaccine. They deliver HIV-1 antigens into host cells and stimulate immune responses.Citation116,117 A number of new vectors, including Ad26, Ad35, replication competent Ad4 and CMV vectors are currently being developed.
Results from the 3 efficacy trials using of replication-defective Ad5 vectors have been disappointing.Citation39,42,66 Furthermore, Ad5 seropositivity was associated with transient increased incidence of HIV-1 infection in male vaccinees in the STEP study.Citation39,118 Though the mechanism for this has not been clearly elucidated, the hypothesis that expansion of Ad5-specific CD4+ T-cells in mucosal tissues with Ad5 vaccination, leading to increased target cell availability and thus increased risk of infection has been proposed.Citation119 Therefore, concern regarding preexisting vector immunity prompted the development of other Ad vectors with lower seroprevalence, including Ad26 and Ad35.Citation120,121 Ad26 and Ad35 studies have shown potential in phase I/II human vaccine trials.Citation122-125 A recent randomized, double-blind, placebo-controlled trial in 218 healthy adults using Ad26.EnvA.01 and Ad35.Env vectors in both homologous and heterologous combinations elicited humoral and cellular immune responses in nearly all participants. Furthermore, preexisting Ad26 or Ad35 neutralizing antibody had no effect on vaccine safety and no significant impact on immunogenicity.Citation125
Replication competent, enteric-coated, Ad4 has been used to prevent acute respiratory disease from adenoviruses by the US military.Citation126 Two phase I trials of replication competent, rAd4 HIV-1 vaccine, NCT01989533 (Ad4 Mosaic Gag/Ad4 clade C Env150 via intranasal or oral routes) and NCT02771730 (Ad4 Mosaic Gag or Ad4 clade C Env150 or both prime, AIDSVAX® B/E/aluminum hydroxide boost) are currently underway ().
Replicating rhesus (Rh) CMV vectors have shown promise in RM models. Vaccination of RM with RhCMV/SIV vectors led to the establishment of indefinitely persistent, high-frequency, SIV-specific effector memory T-cell responses at potential sites of SIV replication and was associated with durable aviremic control of SIV infection in 50% of RM,Citation127 and eventual absence of detectable plasma- or tissue-associated virus in some RM.Citation128 Currently, no human data are available on the safety and efficacy of CMV vector vaccines.
In addition to technical challenges, another major obstacle to HIV vaccine product development is the requirement for increasingly large numbers of participants and resources needed for efficacy assessment. HIV prevention efficacy trials can be conducted efficiently only in large target populations with HIV incidence ≥ 2 per 100-person years, as low event rates would necessitate very large studies and incur huge costs to achieve statistical power.Citation129 Implementation of non-vaccine prevention modalities (NVPM) will lead to very welcome declining HIV incidence in many populations. Lower incidence complicates preventive vaccine trial design by either driving enrollment requirements up substantially, or forcing identification of populations or sub-populations where NVPM implementation is not lowering incidence for some reason beyond the control of the study team. Use of NVPM by vaccine trial participants will also need to be addressed in future vaccine efficacy trials including powering trials to account for background NVPM use and inclusion of NVPM as part of the study intervention where appropriate.Citation130
Conclusion
A great deal has been learned from HIV vaccine research over the last 30 years. Data from completed HIV-1 vaccine efficacy trials support the role of functional Env Ab in reducing infection risk.Citation52 Replication in a follow-up efficacy trial is pending. The field eagerly awaits results from HVTN702, which will provide an opportunity to assess this pox-protein regimen in light of RV144 correlates of risk as well as generate novel hypotheses on mechanisms of protection. Data from phase II studies of passive immunization with bnAb will become available in the next few years and will be critical for proof of concept that bnAb can prevent HIV acquisition in humans. The elicitation of bnAb through vaccination is still in preclinical stages. The recent improvements in the design and generation native ENV trimer will likely aid advancement. Polyvalent mosaic antigens expressed on viral vectors and replicating viral vectors have shown great promise in RM models, are currently under investigation in phase I/II studies and have high potential to expand the HIV vaccine pipeline. The knowledge that has been gained, the results that show partial efficacy, and the robust pipeline give us hope. However, the immense complexity and multitude of unanswered questions remain enormous challenges that have to be overcome.
Disclaimer
The opinions expressed herein are those of the authors and do not represent the official position of the U.S. Army, or The Department of Defense. Trade names are used for identification purposes only and do not imply endorsement.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgment
We would like to thank Dr Sandhya Vasan for her thoughtful suggestions on the design of .
Funding
This work was supported by the U.S. Army Medical Research and Materiel Command (Military Infectious Diseases Research Program) through Cooperative Agreements (W81XWH-11–2–0174) with the Henry M. Jackson Foundation for the Advancement of Military Medicine.
References
- UNAIDS. Gobal AIDS update 2016. Geneva, Switzerland: Joint United Nations Programme on HIV/AIDS (UNAIDS); 2016 (accessed 11 Feb 2017); available at: http://www.unaids.org/sites/default/files/media_asset/global-AIDS-update-2016-en.pdf
- World Health Organization. Global update on HIV treatment 2013: Results, impact and opportunities. Geneva, Switzerland: WHO; 2013 (accessed 11 Feb 2017); available at: http://apps.who.int.iris/bitstream/10665/85326/1/9789241505734_eng.pdf?ua=1
- Cohen MS, Chen YQ, McCauley M, Gamble T, Hosseinipour MC, Kumarasamy N, Hakim JG, Kumwenda J, Grinsztejn B, Pilotto JH, et al. Prevention of HIV-1 infection with early antiretroviral therapy. N Engl J Med 2011; 365:493-505; PMID:21767103; http://dx.doi.org/10.1056/NEJMoa1105243
- Grant RM, Lama JR, Anderson PL, McMahan V, Liu AY, Vargas L, Goicochea P, Casapía M, Guanira-Carranza JV, Ramirez-Cardich ME, et al. Preexposure chemoprophylaxis for HIV prevention in men who have sex with men. N Engl J Med 2010; 363:2587-99; PMID:21091279; http://dx.doi.org/10.1056/NEJMoa1011205
- Baeten JM, Donnell D, Ndase P, Mugo NR, Campbell JD, Wangisi J, Tappero JW, Bukusi EA, Cohen CR, Katabira E, et al. Antiretroviral prophylaxis for HIV prevention in heterosexual men and women. N Engl J Med 2012; 367:399-410; PMID:22784037; http://dx.doi.org/10.1056/NEJMoa1108524
- Finzi D, Blankson J, Siliciano JD, Margolick JB, Chadwick K, Pierson T, Smith K, Lisziewicz J, Lori F, Flexner C, et al. Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat Med 1999; 5:512-7; PMID:10229227; http://dx.doi.org/10.1038/8394
- Siliciano JD, Kajdas J, Finzi D, Quinn TC, Chadwick K, Margolick JB, Kovacs C, Gange SJ, Siliciano RF. Long-term follow-up studies confirm the stability of the latent reservoir for HIV-1 in resting CD4+ T cells. Nat Med 2003; 9:727-8; PMID:12754504; http://dx.doi.org/10.1038/nm880
- Granich R, Gupta S, Montaner J, Williams B, Zuniga JM. Pattern, determinants, and impact of hiv spending on care and treatment in 38 high-burden low- and middle-income countries. J Int Assoc Provid AIDS Care 2016; 15:91-100; PMID:26715490; http://dx.doi.org/10.1177/2325957415623261
- UNAIDS. The Gap Report. Geneva, Switzerland: Joint United Nations Programme on HIV/AIDS (UNAIDS); 2014 (accessed 11 Feb 2017); available at: http://www.unaids.org/sites/default/files/media-asset/UNAIDS_Gap_report_en.pdf
- Deeks SG, Lewin SR, Ross AL, Ananworanich J, Benkirane M, Cannon P, Chomont N, Douek D, Lifson JD, Lo YR, et al. International AIDS Society global scientific strategy: towards an HIV cure 2016. Nat Med 2016; 22:839-50; PMID:27400264; http://dx.doi.org/10.1038/nm.4108
- Siegfried N, van der Merwe L, Brocklehurst P, Sint TT. Antiretrovirals for reducing the risk of mother-to-child transmission of HIV infection. Cochrane Database Syst Rev 2011; 7:CD003510.
- Cohen MS, Chen YQ, McCauley M, Gamble T, Hosseinipour MC, Kumarasamy N, Hakim JG, Kumwenda J, Grinsztejn B, Pilotto JH, et al. Antiretroviral Therapy for the Prevention of HIV-1 Transmission. N Engl J Med 2016; 375:830-9; PMID:27424812
- Okwundu CI, Uthman OA, Okoromah CA. Antiretroviral pre-exposure prophylaxis (PrEP) for preventing HIV in high-risk individuals. Cochrane Database Syst Rev 2012; 7:CD007189; PMID:22786505
- McCormack S, Dunn DT, Desai M, Dolling DI, Gafos M, Gilson R, Sullivan AK, Clarke A, Reeves I, Schembri G, et al. Pre-exposure prophylaxis to prevent the acquisition of HIV-1 infection (PROUD): effectiveness results from the pilot phase of a pragmatic open-label randomised trial. Lancet 2016; 387:53-60; PMID:26364263; http://dx.doi.org/10.1016/S0140-6736(15)00056-2
- Ford N, Shubber Z, Calmy A, Irvine C, Rapparini C, Ajose O, Beanland RL, Vitoria M, Doherty M, Mayer KH, et al. Choice of antiretroviral drugs for postexposure prophylaxis for adults and adolescents: a systematic review. Clin Infect Dis 2015; 60(Suppl 3):S170-6; PMID:25972499; http://dx.doi.org/10.1093/cid/civ092
- Doblecki-Lewis S, Cohen S, Liu A. Clinical treatment options infectious diseases: update on PrEP implementation, adherence, and advances in delivery. Curr Treat Options Infect Dis 2015; 7:101-12; PMID:25931990; http://dx.doi.org/10.1007/s40506-015-0046-4
- Fauci AS, Marston HD. Ending AIDS–is an HIV vaccine necessary? N Engl J Med 2014; 370:495-8; PMID:24499210; http://dx.doi.org/10.1056/NEJMp1313771
- Harmon TM, Fisher KA, McGlynn MG, Stover J, Warren MJ, Teng Y, Näveke A. Exploring the potential health impact and cost-effectiveness of AIDS vaccine within a comprehensive HIV/AIDS response in low- and middle-income countries. PloS one 2016; 11:e0146387; PMID:26731116; http://dx.doi.org/10.1371/journal.pone.0146387
- Pantaleo G, Koup RA. Correlates of immune protection in HIV-1 infection: what we know, what we don't know, what we should know. Nat Med 2004; 10:806-10; PMID:15286782; http://dx.doi.org/10.1038/nm0804-806
- Johnston MI, Fauci AS. HIV vaccine development–improving on natural immunity. N Engl J Med 2011; 365:873-5; PMID:21899447; http://dx.doi.org/10.1056/NEJMp1107621
- Letvin NL. Progress and obstacles in the development of an AIDS vaccine. Nat Rev Immunol 2006; 6:930-9; PMID:17124514; http://dx.doi.org/10.1038/nri1959
- Korber B, Gaschen B, Yusim K, Thakallapally R, Kesmir C, Detours V. Evolutionary and immunological implications of contemporary HIV-1 variation. Br Med Bull 2001; 58:19-42; PMID:11714622; http://dx.doi.org/10.1093/bmb/58.1.19
- Gaschen B, Taylor J, Yusim K, Foley B, Gao F, Lang D, Novitsky V, Haynes B, Hahn BH, Bhattacharya T, et al. Diversity considerations in HIV-1 vaccine selection. Science 2002; 296:2354-60; PMID:12089434; http://dx.doi.org/10.1126/science.1070441
- Plotkin SA. Vaccines: correlates of vaccine-induced immunity. Clin Infect Dis 2008; 47:401-9; PMID:18558875; http://dx.doi.org/10.1086/589862
- Thakur A, Pedersen LE, Jungersen G. Immune markers and correlates of protection for vaccine induced immune responses. Vaccine 2012; 30:4907-20; PMID:22658928; http://dx.doi.org/10.1016/j.vaccine.2012.05.049
- Berman PW, Gregory TJ, Riddle L, Nakamura GR, Champe MA, Porter JP, Wurm FM, Hershberg RD, Cobb EK, Eichberg JW. Protection of chimpanzees from infection by HIV-1 after vaccination with recombinant glycoprotein gp120 but not gp160. Nature 1990; 345:622-5; PMID:2190095; http://dx.doi.org/10.1038/345622a0
- el-Amad Z, Murthy KK, Higgins K, Cobb EK, Haigwood NL, Levy JA, Steimer KS. Resistance of chimpanzees immunized with recombinant gp120SF2 to challenge by HIV-1SF2. AIDS 1995; 9:1313-22; PMID:8605050; http://dx.doi.org/10.1097/00002030-199512000-00003
- Berman PW, Murthy KK, Wrin T, Vennari JC, Cobb EK, Eastman DJ, Champe M, Nakamura GR, Davison D, Powell MF, et al. Protection of MN-rgp120-immunized chimpanzees from heterologous infection with a primary isolate of human immunodeficiency virus type 1. J Infect Dis 1996; 173:52-9; PMID:8537682; http://dx.doi.org/10.1093/infdis/173.1.52
- Pitisuttithum P, Gilbert P, Gurwith M, Heyward W, Martin M, van Griensven F, Hu D, Tappero JW, Choopanya K, Bangkok Vaccine Evaluation Group. Randomized, double-blind, placebo-controlled efficacy trial of a bivalent recombinant glycoprotein 120 HIV-1 vaccine among injection drug users in Bangkok, Thailand. J Infect Dis 2006; 194:1661-71; PMID:17109337; http://dx.doi.org/10.1086/508748
- Flynn NM, Forthal DN, Harro CD, Judson FN, Mayer KH, Para MF, rgp120 HIV Vaccine Study Group. Placebo-controlled phase 3 trial of a recombinant glycoprotein 120 vaccine to prevent HIV-1 infection. J Infect Dis 2005; 191:654-65; PMID:15688278; http://dx.doi.org/10.1086/428404
- Gilbert PB, Peterson ML, Follmann D, Hudgens MG, Francis DP, Gurwith M, Heyward WL, Jobes DV, Popovic V, Self SG, et al. Correlation between immunologic responses to a recombinant glycoprotein 120 vaccine and incidence of HIV-1 infection in a phase 3 HIV-1 preventive vaccine trial. J Infect Dis 2005; 191:666-77; PMID:15688279; http://dx.doi.org/10.1086/428405
- Forthal DN, Gilbert PB, Landucci G, Phan T. Recombinant gp120 vaccine-induced antibodies inhibit clinical strains of HIV-1 in the presence of Fc receptor-bearing effector cells and correlate inversely with HIV infection rate. J Immunol 2007; 178:6596-603; PMID:17475891; http://dx.doi.org/10.4049/jimmunol.178.10.6596
- Shiver JW, Fu TM, Chen L, Casimiro DR, Davies ME, Evans RK, Zhang ZQ, Simon AJ, Trigona WL, Dubey SA, Irwin CE, Safrit JT, Mittler J, Weinberger L, et al. Replication-incompetent adenoviral vaccine vector elicits effective anti-immunodeficiency-virus immunity. Nature 2002; 415:331-5; PMID:11797011; http://dx.doi.org/10.1038/415331a
- Jin X, Bauer DE, Tuttleton SE, Lewin S, Gettie A, Blanchard J, Irwin CE, Safrit JT, Mittler J, Weinberger L, et al. Dramatic rise in plasma viremia after CD8(+) T cell depletion in simian immunodeficiency virus-infected macaques. J Exp Med 1999; 189:991-8; PMID:10075982; http://dx.doi.org/10.1084/jem.189.6.991
- Schmitz JE, Kuroda MJ, Santra S, Sasseville VG, Simon MA, Lifton MA, Racz P, Tenner-Racz K, Dalesandro M, Scallon BJ, et al. Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes. Science 1999; 283:857-60; PMID:9933172; http://dx.doi.org/10.1126/science.283.5403.857
- Klein MR, van Baalen CA, Holwerda AM, Kerkhof Garde SR, Bende RJ, Keet IP, Eeftinck-Schattenkerk JK, Osterhaus AD, Schuitemaker H, Miedema F. Kinetics of Gag-specific cytotoxic T lymphocyte responses during the clinical course of HIV-1 infection: a longitudinal analysis of rapid progressors and long-term asymptomatics. J Exp Med 1995; 181:1365-72; PMID:7699324; http://dx.doi.org/10.1084/jem.181.4.1365
- Emu B, Sinclair E, Hatano H, Ferre A, Shacklett B, Martin JN, McCune JM, Deeks SG. HLA class I-restricted T-cell responses may contribute to the control of human immunodeficiency virus infection, but such responses are not always necessary for long-term virus control. J Virol 2008; 82:5398-407; PMID:18353945; http://dx.doi.org/10.1128/JVI.02176-07
- Altfeld M, Kalife ET, Qi Y, Streeck H, Lichterfeld M, Johnston MN, Burgett N, Swartz ME, Yang A, Alter G, et al. HLA alleles associated with delayed progression to AIDS contribute strongly to the initial CD8(+) T cell response against HIV-1. PLoS Med 2006; 3:e403; PMID:17076553; http://dx.doi.org/10.1371/journal.pmed.0030403
- Buchbinder SP, Mehrotra DV, Duerr A, Fitzgerald DW, Mogg R, Li D, Gilbert PB, Lama JR, Marmor M, Del Rio C, et al. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): a double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 2008; 372:1881-93; PMID:19012954; http://dx.doi.org/10.1016/S0140-6736(08)61591-3
- McElrath MJ, De Rosa SC, Moodie Z, Dubey S, Kierstead L, Janes H, Defawe OD, Carter DK, Hural J, Akondy R, et al. HIV-1 vaccine-induced immunity in the test-of-concept Step Study: a case-cohort analysis. Lancet 2008; 372:1894-905; PMID:19012957; http://dx.doi.org/10.1016/S0140-6736(08)61592-5
- Rolland M, Tovanabutra S, deCamp AC, Frahm N, Gilbert PB, Sanders-Buell E, Heath L, Magaret CA, Bose M, Bradfield A, et al. Genetic impact of vaccination on breakthrough HIV-1 sequences from the STEP trial. Nat Med 2011; 17:366-71; PMID:21358627; http://dx.doi.org/10.1038/nm.2316
- Gray GE, Allen M, Moodie Z, Churchyard G, Bekker LG, Nchabeleng M, Mlisana K, Metch B, de Bruyn G, Latka MH, et al. Safety and efficacy of the HVTN 503/Phambili study of a clade-B-based HIV-1 vaccine in South Africa: a double-blind, randomised, placebo-controlled test-of-concept phase 2b study. Lancet Infect Dis 2011; 11:507-15; PMID:21570355; http://dx.doi.org/10.1016/S1473-3099(11)70098-6
- Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E, et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N Engl J Med 2009; 361:2209-20; PMID:19843557; http://dx.doi.org/10.1056/NEJMoa0908492
- Nitayaphan S, Pitisuttithum P, Karnasuta C, Eamsila C, de Souza M, Morgan P, Polonis V, Benenson M, VanCott T, Ratto-Kim S, et al. Safety and immunogenicity of an HIV subtype B and E prime-boost vaccine combination in HIV-negative Thai adults. J Infect Dis 2004; 190:702-6; PMID:15272397; http://dx.doi.org/10.1086/422258
- Lu S. Heterologous prime-boost vaccination. Curr Opin Immunol 2009; 21:346-51; PMID:19500964; http://dx.doi.org/10.1016/j.coi.2009.05.016
- Robb ML, Rerks-Ngarm S, Nitayaphan S, Pitisuttithum P, Kaewkungwal J, Kunasol P, Khamboonruang C, Thongcharoen P, Morgan P, Benenson M, et al. Risk behaviour and time as covariates for efficacy of the HIV vaccine regimen ALVAC-HIV (vCP1521) and AIDSVAX B/E: a post-hoc analysis of the Thai phase 3 efficacy trial RV 144. Lancet Infect Dis 2012; 12:531-7; PMID:22652344; http://dx.doi.org/10.1016/S1473-3099(12)70088-9
- Montefiori DC, Karnasuta C, Huang Y, Ahmed H, Gilbert P, de Souza MS, McLinden R, Tovanabutra S, Laurence-Chenine A, Sanders-Buell E, et al. Magnitude and breadth of the neutralizing antibody response in the RV144 and Vax003 HIV-1 vaccine efficacy trials. J Infect Dis 2012; 206:431-41; PMID:22634875; http://dx.doi.org/10.1093/infdis/jis367
- Haynes BF, Gilbert PB, McElrath MJ, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, et al. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N Engl J Med 2012; 366:1275-86; PMID:22475592; http://dx.doi.org/10.1056/NEJMoa1113425
- Vasan S, Rerks-Ngarm S, Gilbert P, Haynes B, Nitayapan S, Pitisuttihum P, Kaewkungwal J, Excler JL, Robb M, Michael N, et al. Letter to the Editor on: The RV144 vaccine regimen was not associated with enhancement of infection. Hum Vaccin Immunother 2015; 11:1036-7; PMID:25746053; http://dx.doi.org/10.1080/21645515.2015.1010970
- O'Connell RJ, Excler JL. HIV vaccine efficacy and immune correlates of risk. Curr HIV Res 2013; 11:450-63; PMID:24033301; http://dx.doi.org/10.2174/1570162X113116660052
- Corey L, Gilbert PB, Tomaras GD, Haynes BF, Pantaleo G, Fauci AS. Immune correlates of vaccine protection against HIV-1 acquisition. Sci Translat Med 2015; 7:310rv7; PMID:26491081; http://dx.doi.org/10.1126/scitranslmed.aac7732
- Kim JH, Excler JL, Michael NL. Lessons from the RV144 Thai phase III HIV-1 vaccine trial and the search for correlates of protection. Annu Rev Med 2015; 66:423-37; PMID:25341006; http://dx.doi.org/10.1146/annurev-med-052912-123749
- Rolland M, Edlefsen PT, Larsen BB, Tovanabutra S, Sanders-Buell E, Hertz T, deCamp AC, Carrico C, Menis S, Magaret CA, et al. Increased HIV-1 vaccine efficacy against viruses with genetic signatures in Env V2. Nature 2012; 490:417-20; PMID:22960785; http://dx.doi.org/10.1038/nature11519
- Zolla-Pazner S, deCamp AC, Cardozo T, Karasavvas N, Gottardo R, Williams C, Morris DE, Tomaras G, Rao M, Billings E, et al. Analysis of V2 antibody responses induced in vaccinees in the ALVAC/AIDSVAX HIV-1 vaccine efficacy trial. PloS one 2013; 8:e53629; PMID:23349725; http://dx.doi.org/10.1371/journal.pone.0053629
- Liao HX, Bonsignori M, Alam SM, McLellan JS, Tomaras GD, Moody MA, Kozink DM, Hwang KK, Chen X, Tsao CY, et al. Vaccine induction of antibodies against a structurally heterogeneous site of immune pressure within HIV-1 envelope protein variable regions 1 and 2. Immunity 2013; 38:176-86; PMID:23313589; http://dx.doi.org/10.1016/j.immuni.2012.11.011
- Chung AW, Ghebremichael M, Robinson H, Brown E, Choi I, Lane S, Dugast AS, Schoen MK, Rolland M, Suscovich TJ, et al. Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines. Sci Translat Med 2014; 6:228ra38; PMID:24648341; http://dx.doi.org/10.1126/scitranslmed.3007736
- Tay MZ, Liu P, Williams LD, McRaven MD, Sawant S, Gurley TC, Xu TT, Dennison SM, Liao HX, Chenine AL, et al. Antibody-Mediated Internalization of Infectious HIV-1 Virions Differs among Antibody Isotypes and Subclasses. PLoS Pathog 2016; 12:e1005817; PMID:27579713; http://dx.doi.org/10.1371/journal.ppat.1005817
- Yates NL, Liao HX, Fong Y, deCamp A, Vandergrift NA, Williams WT, Alam SM, Ferrari G, Yang ZY, Seaton KE, et al. Vaccine-induced Env V1-V2 IgG3 correlates with lower HIV-1 infection risk and declines soon after vaccination. Sci Transl Med 2014; 6:228ra39; PMID:24648342; http://dx.doi.org/10.1126/scitranslmed.3007730
- Perez LG, Martinez D, deCamp AC, Pinter A, Berman PW, Francis D, Sinangil F, Lee C, Greene K, Gao H, et al. V1V2-specific complement activating antibodies as a correlate of HIV-1 infection risk in RV144. In submission 2016.
- Liu P, Yates NL, Shen X, Bonsignori M, Moody MA, Liao HX, Fong Y, Alam SM, Overman RG, Denny T, et al. Infectious virion capture by HIV-1 gp120-specific IgG from RV144 vaccinees. J Virol 2013; 87:7828-36; PMID:23658446; http://dx.doi.org/10.1128/JVI.02737-12
- Fahrbach KM, Malykhina O, Stieh DJ, Hope TJ. Differential binding of IgG and IgA to mucus of the female reproductive tract. PloS one 2013; 8:e76176; PMID:24098437; http://dx.doi.org/10.1371/journal.pone.0076176
- Gunn BM, Schneider JR, Shansab M, Bastian AR, Fahrbach KM, Smith AD, Mahan AE, Karim MM, Licht AF, Zvonar I, et al. Enhanced binding of antibodies generated during chronic HIV infection to mucus component MUC16. Mucosal Immunol 2016; 9:1549-58; PMID:26960182; http://dx.doi.org/10.1038/mi.2016.8
- Su B, Moog C. Which antibody functions are important for an HIV vaccine? Front Immunol 2014; 5:289; PMID:24995008; http://dx.doi.org/10.3389/fimmu.2014.00289
- Forthal D, Hope TJ, Alter G. New paradigms for functional HIV-specific nonneutralizing antibodies. Curr Opin HIV AIDS 2013; 8:393-401; PMID:23924999; http://dx.doi.org/10.1097/COH.0b013e328363d486
- Tomaras GD, Ferrari G, Shen X, Alam SM, Liao HX, Pollara J, Bonsignori M, Moody MA, Fong Y, Chen X, et al. Vaccine-induced plasma IgA specific for the C1 region of the HIV-1 envelope blocks binding and effector function of IgG. Proc Natl Acad Sci U S A 2013; 110:9019-24; PMID:23661056; http://dx.doi.org/10.1073/pnas.1301456110
- Hammer SM, Sobieszczyk ME, Janes H, Karuna ST, Mulligan MJ, Grove D, Koblin BA, Buchbinder SP, Keefer MC, Tomaras GD, et al. Efficacy trial of a DNA/rAd5 HIV-1 preventive vaccine. N Engl J Med 2013; 369:2083-92; PMID:24099601; http://dx.doi.org/10.1056/NEJMoa1310566
- Zolla-Pazner S. A critical question for HIV vaccine development: which antibodies to induce? Science 2014; 345:167-8; PMID:25013066; http://dx.doi.org/10.1126/science.1256526
- Stephenson KE, D'Couto HT, Barouch DH. New concepts in HIV-1 vaccine development. Curr Opin Immunol 2016; 41:39-46; PMID:27268856; http://dx.doi.org/10.1016/j.coi.2016.05.011
- Safrit JT, Fast PE, Gieber L, Kuipers H, Dean HJ, Koff WC. Status of vaccine research and development of vaccines for HIV-1. Vaccine 2016; 34:2921-5; PMID:26993335; http://dx.doi.org/10.1016/j.vaccine.2016.02.074
- Burton DR, Hangartner L. Broadly neutralizing antibodies to HIV and their role in vaccine design. Annu Rev Immunol 2016; 34:635-59; PMID:27168247; http://dx.doi.org/10.1146/annurev-immunol-041015-055515
- Sadanand S, Suscovich TJ, Alter G. Broadly neutralizing antibodies against HIV: new insights to inform vaccine design. Annu Rev Med 2016; 67:185-200; PMID:26565674; http://dx.doi.org/10.1146/annurev-med-091014-090749
- Stephenson KE, Barouch DH. Broadly neutralizing antibodies for HIV eradication. Curr HIV/AIDS Rep 2016; 13:31-7; PMID:26841901; http://dx.doi.org/10.1007/s11904-016-0299-7
- Simek MD, Rida W, Priddy FH, Pung P, Carrow E, Laufer DS, Lehrman JK, Boaz M, Tarragona-Fiol T, Miiro G, et al. Human immunodeficiency virus type 1 elite neutralizers: individuals with broad and potent neutralizing activity identified by using a high-throughput neutralization assay together with an analytical selection algorithm. J Virol 2009; 83:7337-48; PMID:19439467; http://dx.doi.org/10.1128/JVI.00110-09
- Doria-Rose NA, Klein RM, Manion MM, O'Dell S, Phogat A, Chakrabarti B, Hallahan CW, Migueles SA, Wrammert J, Ahmed R, et al. Frequency and phenotype of human immunodeficiency virus envelope-specific B cells from patients with broadly cross-neutralizing antibodies. J Virol 2009; 83:188-99; PMID:18922865; http://dx.doi.org/10.1128/JVI.01583-08
- Sather DN, Armann J, Ching LK, Mavrantoni A, Sellhorn G, Caldwell Z, Yu X, Wood B, Self S, Kalams S, et al. Factors associated with the development of cross-reactive neutralizing antibodies during human immunodeficiency virus type 1 infection. J Virol 2009; 83:757-69; PMID:18987148; http://dx.doi.org/10.1128/JVI.02036-08
- Hraber P, Seaman MS, Bailer RT, Mascola JR, Montefiori DC, Korber BT. Prevalence of broadly neutralizing antibody responses during chronic HIV-1 infection. AIDS 2014; 28:163-9; PMID:24361678; http://dx.doi.org/10.1097/QAD.0000000000000106
- Landais E, Huang X, Havenar-Daughton C, Murrell B, Price MA, Wickramasinghe L, Ramos A, Bian CB, Simek M, Allen S, et al. Broadly neutralizing antibody responses in a large longitudinal sub-saharan HIV primary infection cohort. PLoS pathogens 2016; 12:e1005369; PMID:26766578; http://dx.doi.org/10.1371/journal.ppat.1005369
- Gray ES, Madiga MC, Hermanus T, Moore PL, Wibmer CK, Tumba NL, Werner L, Mlisana K, Sibeko S, Williamson C, et al. The neutralization breadth of HIV-1 develops incrementally over four years and is associated with CD4+ T cell decline and high viral load during acute infection. J Virol 2011; 85:4828-40; PMID:21389135; http://dx.doi.org/10.1128/JVI.00198-11
- Kwong PD, Mascola JR, Nabel GJ. Broadly neutralizing antibodies and the search for an HIV-1 vaccine: the end of the beginning. Nat Rev Immunol 2013; 13:693-701; PMID:23969737; http://dx.doi.org/10.1038/nri3516
- Moldt B, Rakasz EG, Schultz N, Chan-Hui PY, Swiderek K, Weisgrau KL, Piaskowski SM, Bergman Z, Watkins DI, Poignard P, et al. Highly potent HIV-specific antibody neutralization in vitro translates into effective protection against mucosal SHIV challenge in vivo. Proc Natl Acad Sci U S A 2012; 109:18921-5; PMID:23100539; http://dx.doi.org/10.1073/pnas.1214785109
- Shingai M, Donau OK, Plishka RJ, Buckler-White A, Mascola JR, Nabel GJ, Nason MC, Montefiori D, Moldt B, Poignard P, et al. Passive transfer of modest titers of potent and broadly neutralizing anti-HIV monoclonal antibodies block SHIV infection in macaques. J Exp Med 2014; 211:2061-74; PMID:25155019; http://dx.doi.org/10.1084/jem.20132494
- Rudicell RS, Kwon YD, Ko SY, Pegu A, Louder MK, Georgiev IS, Wu X, Zhu J, Boyington JC, Chen X, et al. Enhanced potency of a broadly neutralizing HIV-1 antibody in vitro improves protection against lentiviral infection in vivo. J Virol 2014; 88:12669-82; PMID:25142607; http://dx.doi.org/10.1128/JVI.02213-14
- Pegu A, Yang ZY, Boyington JC, Wu L, Ko SY, Schmidt SD, McKee K, Kong WP, Shi W, Chen X, et al. Neutralizing antibodies to HIV-1 envelope protect more effectively in vivo than those to the CD4 receptor. Sci Translat Med 2014; 6:243ra88; PMID:24990883; http://dx.doi.org/10.1126/scitranslmed.3008992
- Gautam R, Nishimura Y, Pegu A, Nason MC, Klein F, Gazumyan A, Buckler-White A, Sadjadpour R, Wang K, Mankoff Z, et al. A single injection of anti-HIV-1 antibodies protects against repeated SHIV challenges. Nature 2016; 533:105-9; PMID:27120156; http://dx.doi.org/10.1038/nature17677
- Liu J, Ghneim K, Sok D, Bosche WJ, Li Y, Chipriano E, Berkemeier B, Oswald K, Borducchi E, Cabral C, et al. Antibody-mediated protection against SHIV challenge includes systemic clearance of distal virus. Science 2016; 353:1045-9; PMID:27540005; http://dx.doi.org/10.1126/science.aag0491
- Ledgerwood JE, Coates EE, Yamshchikov G, Saunders JG, Holman L, Enama ME, DeZure A, Lynch RM, Gordon I, Plummer S, et al. Safety, pharmacokinetics and neutralization of the broadly neutralizing HIV-1 human monoclonal antibody VRC01 in healthy adults. Clin Exp Immunol 2015; 182:289-301; PMID:26332605; http://dx.doi.org/10.1111/cei.12692
- Johnson PR, Schnepp BC, Zhang J, Connell MJ, Greene SM, Yuste E, Desrosiers RC, Clark KR. Vector-mediated gene transfer engenders long-lived neutralizing activity and protection against SIV infection in monkeys. Nat Med 2009; 15:901-6; PMID:19448633; http://dx.doi.org/10.1038/nm.1967
- Balazs AB, Chen J, Hong CM, Rao DS, Yang L, Baltimore D. Antibody-based protection against HIV infection by vectored immunoprophylaxis. Nature 2012; 481:81-4; http://dx.doi.org/10.1038/nature10660
- Balazs AB, Ouyang Y, Hong CM, Chen J, Nguyen SM, Rao DS, An DS, Baltimore D. Vectored immunoprophylaxis protects humanized mice from mucosal HIV transmission. Nat Med 2014; 20:296-300; PMID:24509526; http://dx.doi.org/10.1038/nm.3471
- Byrareddy SN, Kallam B, Arthos J, Cicala C, Nawaz F, Hiatt J, Kersh EN, McNicholl JM, Hanson D, Reimann KA, et al. Targeting alpha4beta7 integrin reduces mucosal transmission of simian immunodeficiency virus and protects gut-associated lymphoid tissue from infection. Nat Med 2014; 20:1397-400; PMID:25419708; http://dx.doi.org/10.1038/nm.3715
- Byrareddy SN, Arthos J, Cicala C, Villinger F, Ortiz KT, Little D, Sidell N, Kane MA, Yu J, Jones JW, et al. Sustained virologic control in SIV+ macaques after antiretroviral and alpha4beta7 antibody therapy. Science 2016; 354:197-202; PMID:27738167; http://dx.doi.org/10.1126/science.aag1276
- Fauci AS. An HIV Vaccine: Mapping Uncharted Territory. JAMA 2016; 316:143-4; PMID:27404178; http://dx.doi.org/10.1001/jama.2016.7538
- Scheid JF, Mouquet H, Ueberheide B, Diskin R, Klein F, Oliveira TY, Pietzsch J, Fenyo D, Abadir A, Velinzon K, et al. Sequence and structural convergence of broad and potent HIV antibodies that mimic CD4 binding. Science 2011; 333:1633-7; PMID:21764753; http://dx.doi.org/10.1126/science.1207227
- Walker LM, Huber M, Doores KJ, Falkowska E, Pejchal R, Julien JP, Wang SK, Ramos A, Chan-Hui PY, Moyle M, et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011; 477:466-70; PMID:21849977; http://dx.doi.org/10.1038/nature10373
- Burton DR, Ahmed R, Barouch DH, Butera ST, Crotty S, Godzik A, Kaufmann DE, McElrath MJ, Nussenzweig MC, Pulendran B, et al. A Blueprint for HIV Vaccine Discovery. Cell Host Microbe 2012; 12:396-407; PMID:23084910; http://dx.doi.org/10.1016/j.chom.2012.09.008
- Sanders RW, Vesanen M, Schuelke N, Master A, Schiffner L, Kalyanaraman R, Paluch M, Berkhout B, Maddon PJ, Olson WC, et al. Stabilization of the soluble, cleaved, trimeric form of the envelope glycoprotein complex of human immunodeficiency virus type 1. J Virol 2002; 76:8875-89; PMID:12163607; http://dx.doi.org/10.1128/JVI.76.17.8875-8889.2002
- Sanders RW, Derking R, Cupo A, Julien JP, Yasmeen A, de Val N, Kim HJ, Blattner C, de la Peña AT, Korzun J, et al. A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS pathogens 2013; 9:e1003618; PMID:24068931; http://dx.doi.org/10.1371/journal.ppat.1003618
- Sanders RW, van Gils MJ, Derking R, Sok D, Ketas TJ, Burger JA, Ozorowski G, Cupo A, Simonich C, Goo L, et al. HIV-1 VACCINES. HIV-1 neutralizing antibodies induced by native-like envelope trimers. Science 2015; 349:aac4223; PMID:26089353; http://dx.doi.org/10.1126/science.aac4223
- Kong L, He L, de Val N, Vora N, Morris CD, Azadnia P, Sok D, Zhou B, Burton DR, Ward AB, et al. Uncleaved prefusion-optimized gp140 trimers derived from analysis of HIV-1 envelope metastability. Nat Commun 2016; 7:12040; PMID:27349805; http://dx.doi.org/10.1038/ncomms12040
- He L, de Val N, Morris CD, Vora N, Thinnes TC, Kong L, Azadnia P, Sok D, Zhou B, Burton DR, et al. Presenting native-like trimeric HIV-1 antigens with self-assembling nanoparticles. Nat Commun 2016; 7:12041; PMID:27349934; http://dx.doi.org/10.1038/ncomms12041
- Haynes BF, Kelsoe G, Harrison SC, Kepler TB. B-cell-lineage immunogen design in vaccine development with HIV-1 as a case study. Nat Biotechnol 2012; 30:423-33; PMID:22565972; http://dx.doi.org/10.1038/nbt.2197
- Jardine JG, Ota T, Sok D, Pauthner M, Kulp DW, Kalyuzhniy O, Skog PD, Thinnes TC, Bhullar D, Briney B, et al. HIV-1 VACCINES. Priming a broadly neutralizing antibody response to HIV-1 using a germline-targeting immunogen. Science 2015; 349:156-61; PMID:26089355; http://dx.doi.org/10.1126/science.aac5894
- Jardine JG, Kulp DW, Havenar-Daughton C, Sarkar A, Briney B, Sok D, Sesterhenn F, Ereño-Orbea J, Kalyuzhniy O, Deresa I, et al. HIV-1 broadly neutralizing antibody precursor B cells revealed by germline-targeting immunogen. Science 2016; 351:1458-63; PMID:27013733; http://dx.doi.org/10.1126/science.aad9195
- Briney B, Sok D, Jardine JG, Kulp DW, Skog P, Menis S, Jacak R, Kalyuzhniy O, de Val N, Sesterhenn F, et al. Tailored immunogens direct affinity maturation toward HIV neutralizing antibodies. Cell 2016; 166:1459-70 e11; PMID:27610570; http://dx.doi.org/10.1016/j.cell.2016.08.005
- Rerks-Ngarm S, Pitisuttithum P, Excler JL, Nitayaphan S, Kaewkungwal J, Premsri N, Kunasol P, Karasavvas N, Schuetz A, Ngauy V, et al. Randomized, double blind evaluation of late boost strategies for HIV-uninfected vaccine recipients in the RV144 HIV vaccine efficacy trial. In submission 2016.
- Akapirat S, Karnasuta C, Ngauy V, O'Connell RJ, Pitisuttithum P, Rerks-Ngarm S, Michael NL, De Souza MS, Kim JH, Karasavvas N. Antibody Responses in Anogenital Secretions of RV305 a Late Boost Vaccination of RV144 Volunteers. CROI. Boston, MA, USA, 2014.
- Akapirat S, Madnote S, Pitisuttithum P, Nitayaphan S, Chariyalertsak S, Puangkaew J, Rittiroongrad S, Chantakulkij S, Phogat S, Sinangil F, et al. Characterization of HIV-1 Envelope Antibody Responses Following ALVAC-HIV and AIDSVAXB/E Prime-boost Vaccinations with Varying Late Boosts (RV306). HIVR4P. Chicago, IL, USA, 2016.
- Wieczorek L, Pitisutthithum P, Nitayaphan S, Chariyalertsak S, Kaewkungwal J, Molnar S, Blaskowski S, Schoen J, Gao H, Greene K, et al. Delayed Boost of ALVAC®HIV and AIDSVAX® B/E gp120 Following the RV144 Regimen, Significantly Increases HIV-1 Neutralizing Antibody Responses. HIVR4P. Chicago, IL, USA, 2016.
- Akapirat S, Madnote S, Pitisuttithum P, Nitayaphan S, Chariyalertsak S, Rittiroongrad S, Puangkaew J, Chantakulkij S, Phogat S, Sinangil F, et al. Characterization of Antibody Responses in Anogenital Secretions of ALVAC-HIV and AIDSVAXB/E Prime-boost Vaccinations with Varying Late Boosts (RV306). HIVR4P. Chicago, IL, USA, 2016.
- Bekker LG, Laher F, Moodie Z, Tomaras G, Grunenberg N, Allen M, Daniels B, Innes C, Mngadi K, Malahleha M, et al. Meeting the “Go” Criteria: immunogenicity from HVTN100, a phase 1/2 randomized, double blind, placebo-controlled trial of clade C ALVAC-® (vCP2438) and Bivalent Subtype C gp120/MF59® in HIV-uninfected South African adults. AIDS 2016. Durban, South Africa, 2016.
- Fischer W, Perkins S, Theiler J, Bhattacharya T, Yusim K, Funkhouser R, Kuiken C, Haynes B, Letvin NL, Walker BD, et al. Polyvalent vaccines for optimal coverage of potential T-cell epitopes in global HIV-1 variants. Nat Med 2007; 13:100-6; PMID:17187074; http://dx.doi.org/10.1038/nm1461
- Barouch DH, O'Brien KL, Simmons NL, King SL, Abbink P, Maxfield LF, Sun YH, La Porte A, Riggs AM, Lynch DM, et al. Mosaic HIV-1 vaccines expand the breadth and depth of cellular immune responses in rhesus monkeys. Nat Med 2010; 16:319-23; PMID:20173752; http://dx.doi.org/10.1038/nm.2089
- Santra S, Liao HX, Zhang R, Muldoon M, Watson S, Fischer W, Fischer W, Theiler J, Szinger J, Balachandran H. Mosaic vaccines elicit CD8+ T lymphocyte responses that confer enhanced immune coverage of diverse HIV strains in monkeys. Nat Med 2010; 16:324-8; PMID:20173754; http://dx.doi.org/10.1038/nm.2108
- Barouch DH, Stephenson KE, Borducchi EN, Smith K, Stanley K, McNally AG, Liu J, Abbink P, Maxfield LF, Seaman MS, et al. Protective efficacy of a global HIV-1 mosaic vaccine against heterologous SHIV challenges in rhesus monkeys. Cell 2013; 155:531-9; PMID:24243013; http://dx.doi.org/10.1016/j.cell.2013.09.061
- Barouch DH, Alter G, Broge T, Linde C, Ackerman ME, Brown EP, Borducchi EN, Smith KM, Nkolola JP, Liu J, et al. Protective efficacy of adenovirus/protein vaccines against SIV challenges in rhesus monkeys. Science 2015; 349:320-4; PMID:26138104; http://dx.doi.org/10.1126/science.aab3886
- Ewer KJ, Lambe T, Rollier CS, Spencer AJ, Hill AV, Dorrell L. Viral vectors as vaccine platforms: from immunogenicity to impact. Curr Opin Immunol 2016; 41:47-54; PMID:27286566; http://dx.doi.org/10.1016/j.coi.2016.05.014
- Barouch DH, Picker LJ. Novel vaccine vectors for HIV-1. Nat Rev Microbiol 2014; 12:765-71; PMID:25296195; http://dx.doi.org/10.1038/nrmicro3360
- Duerr A, Huang Y, Buchbinder S, Coombs RW, Sanchez J, del Rio C, Casapia M, Santiago S, Gilbert P, Corey L, et al. Extended follow-up confirms early vaccine-enhanced risk of HIV acquisition and demonstrates waning effect over time among participants in a randomized trial of recombinant adenovirus HIV vaccine (Step Study). J Infect Dis 2012; 206:258-66; PMID:22561365; http://dx.doi.org/10.1093/infdis/jis342
- Benlahrech A, Harris J, Meiser A, Papagatsias T, Hornig J, Hayes P, Lieber A, Athanasopoulos T, Bachy V, Csomor E, et al. Adenovirus vector vaccination induces expansion of memory CD4 T cells with a mucosal homing phenotype that are readily susceptible to HIV-1. Proc Natl Acad Sci U S A 2009; 106:19940-5; PMID:19918060; http://dx.doi.org/10.1073/pnas.0907898106
- D'Souza MP, Yang OO. Adenovirus vectors as HIV-1 vaccines: where are we? What next? AIDS 2015; 29:395-400; PMID:25630039; http://dx.doi.org/10.1097/QAD.0000000000000548
- Barouch DH, Kik SV, Weverling GJ, Dilan R, King SL, Maxfield LF, Clark S, Ng'ang'a D, Brandariz KL, Abbink P, et al. International seroepidemiology of adenovirus serotypes 5, 26, 35, and 48 in pediatric and adult populations. Vaccine 2011; 29:5203-9; PMID:21619905; http://dx.doi.org/10.1016/j.vaccine.2011.05.025
- Baden LR, Walsh SR, Seaman MS, Tucker RP, Krause KH, Patel A, Johnson JA, Kleinjan J, Yanosick KE, Perry J, et al. First-in-human evaluation of the safety and immunogenicity of a recombinant adenovirus serotype 26 HIV-1 Env vaccine (IPCAVD 001). J Infect Dis 2013; 207:240-7; PMID:23125444; http://dx.doi.org/10.1093/infdis/jis670
- Barouch DH, Liu J, Peter L, Abbink P, Iampietro MJ, Cheung A, Alter G, Chung A, Dugast AS, Frahm N, et al. Characterization of humoral and cellular immune responses elicited by a recombinant adenovirus serotype 26 HIV-1 Env vaccine in healthy adults (IPCAVD 001). J Infect Dis 2013; 207:248-56; PMID:23125443; http://dx.doi.org/10.1093/infdis/jis671
- Keefer MC, Gilmour J, Hayes P, Gill D, Kopycinski J, Cheeseman H, Cashin-Cox M, Naarding M, Clark L, Fernandez N, et al. A phase I double blind, placebo-controlled, randomized study of a multigenic HIV-1 adenovirus subtype 35 vector vaccine in healthy uninfected adults. PloS one 2012; 7:e41936; PMID:22870265; http://dx.doi.org/10.1371/journal.pone.0041936
- Baden LR, Karita E, Mutua G, Bekker LG, Gray G, Page-Shipp L, Walsh SR, Nyombayire J, Anzala O, Roux S, et al. Assessment of the safety and immunogenicity of 2 novel vaccine platforms for HIV-1 prevention: a randomized trial. Ann Intern Med 2016; 164:313-22; PMID:26833336; http://dx.doi.org/10.7326/M15-0880
- Gaydos CA, Gaydos JC. Adenovirus vaccines in the U.S. military. Mil Med 1995; 160:300-4; PMID:7659229
- Hansen SG, Ford JC, Lewis MS, Ventura AB, Hughes CM, Coyne-Johnson L, Whizin N, Oswald K, Shoemaker R, Swanson T, et al. Profound early control of highly pathogenic SIV by an effector memory T-cell vaccine. Nature 2011; 473:523-7; PMID:21562493; http://dx.doi.org/10.1038/nature10003
- Hansen SG, Piatak M, Jr., Ventura AB, Hughes CM, Gilbride RM, Ford JC, Oswald K, Shoemaker R, Li Y, Lewis MS, et al. Immune clearance of highly pathogenic SIV infection. Nature 2013; 502:100-4; PMID:24025770; http://dx.doi.org/10.1038/nature12519
- Donnell D, Hughes JP, Fleming TR. Challenges in the design of HIV prevention trials in the United States. J Acquir Immun Defic Syndr 2010; 55(Suppl 2):S136-40; PMID:21406984; http://dx.doi.org/10.1097/QAI.0b013e3181fbcb61
- Janes H, Gilbert P, Buchbinder S, Kublin J, Sobieszczyk ME, Hammer SM. In pursuit of an HIV vaccine: designing efficacy trials in the context of partially effective nonvaccine prevention modalities. AIDS Res Hum Retroviruses 2013; 29:1513-23; PMID:23597282; http://dx.doi.org/10.1089/aid.2012.0385