
ABSTRACT
Lyme disease is the most common vector-borne disease in North America. The etiological agent is the spirochete Borreliella burgdorferi, transmitted to mammalian hosts by the Ixodes tick. In recent years there has been an increase in the number of cases of Lyme disease. Currently, there is no vaccine on the market for human use. We describe the development of a novel synthetically engineered DNA vaccine, pLD1 targeting the outer-surface protein A (OspA) of Borreliella burgdorferi. Immunization of C3 H/HeN mice with pLD1 elicits robust humoral and cellular immune responses that confer complete protection against a live Borreliella burgdorferi bacterial challenge. We also assessed intradermal (ID) delivery of pLD1 in Hartley guinea pigs, demonstrating the induction of robust and durable humoral immunity that lasts at least 1 year. We provide evidence of the potency of pLD1 by showing that antibodies targeting the OspA epitopes which have been associated with protection are prominently raised in the immunized guinea pigs. The described study provides the basis for the advancement of pDL1 as a potential vaccine for Lyme disease control.
Background
Lyme disease is highly prevalent in Europe and North America.Citation1 The etiological agents are spirochetes of the Borreliella burgdorferi group and related species.Citation2 These bacteria are transmitted by ticks, with small mammals serving as reservoirs. In the United States, Borreliella burgdorferi is the most prevalent Lyme disease-associated species, and the number of reported cases have doubled over the past 15 years.Citation3 Thirty-thousand cases of Lyme disease are reported to the CDC in the US each year, and predicts the number of clinician-diagnosed cases exceeds 300,000 a year.Citation3,Citation4 The majority of patients respond adequately to antibiotic treatment. However, a significant number of patients develop persistent symptoms with local and systemic manifestations that vary in severity and duration that can last for months to years,Citation5 termed post-treatment Lyme disease syndrome (PTLDS). The armamentarium available to the medical community to tackle the increasing incidence of Lyme disease, and to effectively prevent or treat PTLDS is limited.
To develop effective countermeasures against Lyme disease several targets and mechanisms have been investigated. Multiple studies have demonstrated that the transmission of Borreliella Burgdorferi from tick to mammalian host can be interrupted with antibodies targeting the OspA antigen.Citation6–9 OspA mediates the attachment of the spirochetes to the tick midgut, and its expression is downregulated during the blood-feeding process when the bacteria migrate from the midgut to salivary glands; consequently, anti-OspA antibodies harbored in the host can block bacterial transmission during hematophagy. Vaccines targeting OspA have been developed. The recombinant OspA protein vaccine, LYMErix demonstrated efficacy, and was briefly available on the market.Citation10 The efficacy of this recombinant protein OspA vaccine was modest, 49% after 2 immunizations, and yearly boosts were required. Furthermore, there were concerns surrounding reports of immune cross-reactivity between an epitope present in OspA and the human LFA-1 antigen, and the postulation that vaccination may initiate an autoreactive immune mechanism leading to Lyme-associated arthritis.Citation11 Additionally, poor representation of the vaccine in the media and lack of public enthusiasm resulted in poor sales. The vaccine was ultimately pulled from the market in 2002. Post hoc assessment revealed the vaccine to be safe with no difference in arthritis cases between vaccinated and controls, and the postulation that the OspA vaccine-elicited cross-reactive anti-LFA-1 responses that could mediate arthritis was ultimately withdrawn.Citation11,Citation12 Currently, there is no Lyme disease vaccine available for human use.
Newer engineered synthetic nucleic acid vaccines delivered by advanced CELLECTRA® adaptive electroporation have proven to be highly effective in eliciting robust and long-lasting immunity in multiple clinical studies against infectious disease targets, including Zika, Ebola and MERS.Citation13–15 In these studies, rapid seroconversion at durable high titers, neutralizing antibodies, passive transfer protection to animal challenges from human sera, and induction of CD4 and CD8 T cells has been observed in most subjects.Citation13–15 Furthermore, in other therapeutic studies involving HPV-related cervical infections, strong T cell responses and T cell invasion into the infected tissues as well as viral clearance have been observed.Citation16,Citation17 Based on efficacy in clearing HPV infection in the cervix, a phase III study is in progress. Collectively these studies have established a strong safety database in over 1600 individuals, and demonstrated that this advanced platform can reproducibly drive humoral and cellular immunity in humans with relevance to the prevention of infections or clearance of infectious disease. Potentially the induction of T cell responses by this vaccine platform may afford improved B cell help to generate durable humoral immunity.Citation18
The current manuscript describes the development of a synthetic consensus (SynCon®) OspA DNA vaccine, pLD1 in combination with CELLECTRA® in vivo delivery technology. The data were benchmarked against an OspA and OspC chimeric recombinant protein vaccine, VANGUARDcrLyme, an approved veterinary vaccine, henceforth referred to as crLyme. The data show that pDL1 DNA vaccine elicits robust humoral and cellular immune responses in mice against the OspA antigen that confers protection against Borreliella burgdorferi spirochete tissue colonization following a live bacterial challenge. Also, intradermal immunization of guinea pigs with pLD1 leads to a durable and robust antibody response which persists for at least a year. Moreover, antibodies from the immunized guinea pigs effectively compete for OspA binding with a borreliacidal human mAb 319–44.Citation19,Citation20
Methods
Cell culture
Human embryonic kidney 293 T cells were cultured in Dulbecco modified eagle’s medium (DMEM) and supplemented with 10% heat-inactivated fetal bovine serum (FBS), along with 100 U/ml of penicillin and streptomycin each. Cells were incubated at 37°C and 5% CO2.
Immunofluorescence imaging
For Immunofluorescence analysis, 1.5 × 106 cells were seeded per well into a 6-well cell culture plate, and transfected with either GFP (transfection control), pVAX or pLD1 using the Lipofectamine 3000 transfection kit (ThermoFisher, Waltham, MA) following the manufacturer’s instructions. Forty-eight hours later transfected cells were washed thrice with PBS, and incubated overnight at 4°C with a mouse anti-OspA monoclonal antibody (Clone 0551 LSBio, LS-C103210). Cells were washed with PBS and incubated with Alexa fluor® 555 donkey anti-mouse secondary antibody (Life Technologies, A-31570). The nucleus was stained with 4′, 6-diamidino-2-phenylindole (DAPI) at room temperature for 10 min. Images were captured using ImageXpress Pico automated cell imaging system (Molecular Devices).
pDNA vaccine design
A proprietary synthetic consensus sequence was generated to target OspA serotype 1 (ST1). (pLD1). The sequences were codon and RNA optimized.Citation21,Citation22 The resulting nucleotide sequences were synthesized, digested with BamHI and XhoI, and cloned into the expression vector pVAX under the control of the human cytomegalovirus immediate-early promoter and a bovine growth hormone polyadenylation signal (Genscript, Piscataway, New Jersey).
Animals
Female Hartley Guinea pigs (8–10 weeks) weighing 500–600 g and C3 H/HeN female mice (6–8 weeks) were grouped and housed with ad libitum access to food and water. Animals were housed at Acculab (San Diego, CA 92121) or the University of Florida (Department of Infectious Diseases and Immunology, College of Veterinary Medicine, University of Florida, Gainesville, FL 32608). All housing, handling and treatment protocols were approved and handled according to the standards of the Institutional Animal Care and Use Committee.
Mouse immunizations
30 µl DNA vaccine pLD1 or pVAX formulations were administered by 25 G needle intramuscular to the tibialis anterior muscle and this step was followed by CELLECTRA® in vivo electroporation using the 3P array. The CELLECTRA® EP delivery consists of two sets of pulses with 0.2 Amp constant current. Second pulse set is delayed 3 s. Within each set, there are two 52 ms pulses with a 198 ms delay between the pulses. The vaccine dose was 25 µg of plasmid DNA per injection. Each group was dosed on week 0, 2 and 4 for a total of three immunizations.
Guinea pig immunizations
Hartley guinea pigs were immunized either by intradermal injection (Mantoux technique) of pLD1 on the abdominal left flank followed by CELLECTRA®-EP with the 3p array (as described above) or subcutaneous (SC) injection of a recombinant protein vaccine. Treatments consisted of a total of three immunizations each at a dose of 0.1 mg in 0.1 mL volume of pLD1 administrated at weeks 0, 3 and 6. The crLyme vaccine group of animals was immunized SC with 250 µl of vaccine, commercially available for Lyme disease prevention in dogs (VANGUARDcrLYME by Zoetis).
Mouse splenocytes isolation
Briefly, spleens from mice were collected individually in 5 mL of RPMI1640 media supplemented with 10% FBS (R10), processed into single-cell suspensions with a gentle MACS Dissociator (Miltenyi Biotec, Auburn, CA), then centrifuged at 1500 rpm for 5 min. Cell pellets were resuspended in 5 mL of ACK lysis buffer (Life Technologies, Carlsbad, CA) for 5 min at room temperature, and 10 ml PBS was then added to dilute the lysis buffer. The samples were again centrifuged at 1500 rpm for 5 min, cell pellets resuspended in R10, and then passed through a 45 um nylon filter before use in ELISpot assay.
Enzyme-linked immunospot (ELISpot) assays
To assess cellular IFNγ responses, mouse ELISpot assays were performed using commercial Mabtech pre-coated IFNy ELISpot kits (Mabtech 3321–4APW-10, Sweden). The 96-well ELISpot plates were washed 3 times with sterile 1X PBS and blocked with R10 medium for 1 h. The R10 was removed and 200,000 mouse splenocytes in R10 media were added to each well and incubated at 37°C in 5% CO2 the presence of peptide pool 1, 2 or 3, each consisting of 15-mers overlapping by 9 amino acids, together these pools spanned the length of OspA protein, DMSO (negative control), ConA (positive control for mouse). After 18–20 h, plates were washed and developed according to the manufacturer’s protocols, and IFN-γ positive spots were counted by an automated ELISpot reader (CTL, Shaker Heights, OH). OspA peptide-reactive responses were calculated by subtracting the number of spots in DMSO from peptide wells. Results (Cellular responses or Interferon-gamma ELISpot responses) are shown as individual animal spot-forming units (SFU)/106 PBMCs.
Enzyme-linked immunosorbent assays (ELISAs)
ELISAs were performed to determine sera IgG antigen-binding titers. Nunc ELISA plates were coated with 1 µg/ml recombinant OspA protein (Immune Technology, New York, NY) in DPBS overnight at 4°C. Plates were washed three times then blocked with 3% BSA DPBS with 0.05% Tween 20 for 2 h at 37°C. Plates were then washed and incubated with serial dilutions of mouse or guinea pig sera and incubated for 2 h at 37°C. Plates were again washed and then incubated with HRP conjugated-species specific secondary antibodies (1 in 10,000 dilution) and incubated for 1 h at 37°C. After the final wash, plates were developed using SureBlue TMB Component peroxidase substrate and the reaction stopped with TMB stop reagent (KPL). Plates are then read at 450 nm within 30 min using a SpectraMax Plus384 Microplate Reader (Molecular Devices).
Competition inhibition enzyme-linked immunosorbent assays (ciELISAs)
ciELISAs were performed to determine the presence of antibodies in the serum after OspA immunization in guinea pigs competed with 319–44 human monoclonal antibody known to bind the OspA epitope targeted by the LA-2 antibody.Citation19,Citation20 ELISA plates were coated with 0.5 µg/ml recombinant OspA protein (Immune Technology, New York, NY) in DPBS overnight at 4°C. Plates were washed three times then blocked with 3% BSA DPBS with 0.05% Tween 20 for 2 h at 37°C. Plates were then washed and incubated with serial dilutions (1 in 3) of guinea pig sera and incubated for 2 h at 37°C. Plates were washed and then incubated with 0.75 µg/ml of 319–14 human antibody for 1 h at room temperature. HRP conjugated- anti-human kappa light chain (Bethyl Laboratories) was added for 1 h at room temperature. After the final wash, plates were developed using SureBlue TMB component peroxidase substrate and the reaction stopped with TMB stop reagent (KPL). Plates were then read at 450 nm within 30 min using a SpectraMax Plus384 Microplate Reader (Molecular Devices).
Cultivation of Borreliella burgdorferi
Low-passage B. burgdorferi strain 297 was maintained at 23°C in Barbour-Stoenner-Kelley medium containing 6% normal rabbit serum (BSK Media, Complete; Sigma Aldrich, St. Louis, MO; Cat # B8291) and then temperature-shifted to 37°C. Increased expression of OspC as a virulence factor and presence of OspA as a target for the vaccine was confirmed by Coomassie staining of whole borrelial lysates separated by SDS-PAGE.
Live bacterial challenge
Mice were challenged with 1 × 105 bacteria injected subcutaneously at the sternum as described previously.Citation23 Bacteria were confirmed OspA+ by PCR. All the mice were bled 2 days before the challenge from a facial vein using. Four weeks post-inoculation, mice were anesthetized for intracardiac bleeding, and euthanized for tissue collection that included joints, heart, ear and bladder, respectively.
Isolation of DNA and determination of the bacterial burden in infected tissues
DNA was isolated from the excised tissues using a Quick-DNA miniprep kit (Zymo Research; Cat # D3024). Bacterial burden in tissues was determined using genomic DNA as previously described.Citation24 Briefly, 80 ng of extracted genomic DNA was along with B. burgdorferi-specific flaB primers (200 nM) and probe (320 nM) or primers (400 nM) and probe (320 nM) directed against the mouse actin gene, as described elsewhere.Citation25 Standards were developed to quantify the number of B. burgdorferi flaB and mouse actin copies across a range of 10°–106 copies. Amplification data were acquired and analyzed using the MIC Real-time PCR system, and quantification of target DNA was accomplished as previously described.Citation26
Assessing bactericidal capacity of the serum
Freshly grown temperature-shifted B. burgdorferi strain 297 was counted, and 1 × 105 bacteria incubated with 200 µl two-fold serially diluted serum from the study animals for 18 h in 200ul tubes. At this stage, we included naïve serum from C3 H/HeN mice as a negative control as well as a heat-inactivated counterpart. 50 µl of the aliquots (in triplicate) were inoculated in 1.5 ml microfuge tubes with 1.2 ml of BSK complete media and sealed with parafilm to create microaerobiosis. These tubes were kept at 37°C for 2 weeks. The cultures were monitored daily for change in pH, and if pH drops down, the bacteria were visualized. The presence of bacteria visualized in all the tubes under a dark-field microscope. The highest dilution of the serum that prevented the growth of B. burgdorferi was recorded as a bactericidal titer of the serum.
Statistical analysis
Data were presented as mean ± SEM for all data points. The statistical difference between individual groups was assessed using Mann Whitney test with statistical significance set at p < .05.
Data availability
Data that support the findings of this study are available from the corresponding author upon reasonable request.
Results
SynCon® OspA DNA vaccine engineering
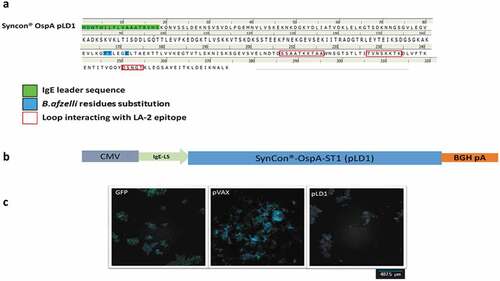
A synthetic consensus sequence was generated to target OspA serotype 1 (ST1), the vaccine is intended for North American use. Thirteen full-length protein sequences from 13 orthologous open reading frames of Borreliella burgdorferi were collected. A consensus was generated by alignment of the OspA protein sequences using ClustalW, sharing 98.5%-100% identity with each individual OspA sequence. Three residues of the consensus were replaced with the corresponding residues of an OspA protein sequence from B. afzelii in order to eliminate an epitope homology with human leucocyte function antigen 1 (LFA-1) (Y165 F, V166 T, T170 K).Citation27 The pLD1 construct was generated using this modified OspA consensus sequence ()). SynCon® OspA ST1 (Design A, pLD1), a potentially exposed N-terminal Cys17 was deleted from the full length modified OspA consensus to avoid possible post-translational modifications and covalent conjugations and the endogenous N-terminal leader sequence was replaced with an immunoglobulin E (IgE) leader sequence to enhance the translation of the construct which has been demonstrated to increase the immunogenicity of the translated product.Citation28 The final sequence was codon and RNA optimized and cloned into the pVAX plasmid (see )). Expression of the OspA was confirmed in vitro using immuno-fluorescence of 293 T cells transfected with pLD1 ()).
Figure 1. Engineering of SynCon® sequence of OspA DNA vaccine pLD1. (a) Protein sequence of pLD1. (b) Schematic of pLD1 construct, OspA antigen is encoded in a pVAX plasmid under CMV promoter. The IgE leader sequences were inserted at the 5ʹend to enhance protein expression. (c) Immunofluorescent image of 293 T cells transfected with GFP, pVAX or pLD1, nucleus stained for DAPI (blue) and immune-stained for anti-OspA (red Alexa fluor® 555 stain).
Characterization of anti-OspA immune response elicited by pLD1 vaccination in C3 H/HeN mice
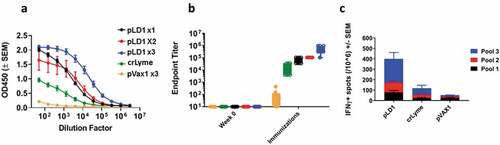
We evaluated the immunogenicity of the OspA antigen expressed after PLD1 vaccination in C3 H/HeN mice. Immunogenicity analyses revealed a robust immune response mediated by the pLD1 vaccine after a single dose, and the response was boosted after subsequent doses ()). On a different set of animals, we examined the cellular immune response to pLD1 vaccination against three pools of overlapping peptides, which together spanned the length of the OspA antigen. Here, groups of 8 mice received three doses of either pLD1, pVAX or crLyme, recombinant protein LD vaccine. All animals were female, we did not address the sex of the animals as a biological variable. Two weeks after the final immunization cellular IFN-γ response was evaluated to OspA peptide recall. As illustrated on ), pLD1 elicited a significantly stronger cellular response than the control (pVAX1) and crLyme groups, with the epitopes in pool 3 being where the cellular response predominantly targeted.
Figure 2. Robust humoral and cellular immune responses are elicited by pLD1 in C3 H/HeN: C3 H/HeN mice received the number of doses of pLD1, crLyme and pVAX as indicated and described in materials and methods. (a) OspA antigen binding of IgG in serial serum dilutions from C3 H/HeN mice measured two weeks after the indicated dose. Data shown represent OD450 nm values (± SEM) for sera OspA antigen binding for each group. (b) OspA-specific IgG endpoint serum binding titer (EBT). The antibody EBT titer was defined as the highest dilution of serum with OspA binding OD values two times the background of pre-bleed serum. (n = 8 per group). (c) C3 H/HeN mice received three doses of pLD1, pVAX, or crLyme vaccine as described in the materials and methods. T cell response was evaluated two weeks after the third dose by measuring the number of IFN-γ producing spot-forming units (SFUs) generated per 106 splenocytes after stimulation with OspA peptide pools. Data show mean SFU±SEM (n = 5 per group).
Single dose of pLD1 confers protection against Borreliella burgdorferi needle challenge in C3 H/HeN mice
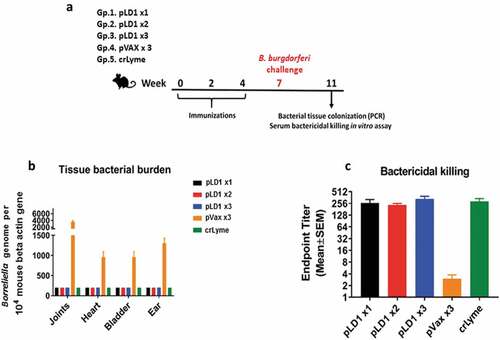
We evaluated the efficacy of pLD1 in a live bacterial challenge. Schematic of the challenge study is illustrated in ). C3 H/HeN mice are susceptible to Borreliella burgdorferi infection, and the model recapitulates some pathophysiology features of human Lyme disease, such as sub-acute arthritis, which is a consequence of the bacterial spread and colonization in the animal’s organs and tissues.Citation23,Citation29,Citation30 Three weeks after the final immunization mice were inoculated with 100,000 of Borreliella burgdorferi strain 297, and 4 weeks later bacterial burden was evaluated by PCR in a double-blinded manner. As shown in ), only the pVAX control group presented with detectable bacterial burden in joints, heart and bladder as well as ear tissues. In groups of mice that received one, two or three dose of the pLD1 vaccine or three doses of crLyme vaccine, a bacterial presence in the tissues tested was not detected. We proceeded to determine the presence of functional anti-B. burgdorferi antibodies in the animals’ serum employing a bacterial killing assay. This assay measures the dilution of serum at which the bacterial growth is enabled. As illustrated in ) bacterial growth was enabled at a serum dilution of approximately 256 for pLD1 and crLyme immunized mice indicating robust bactericidal activity. These results from both bacterial tissue burden and bacterial growth inhibition assay demonstrate the efficacy of pLD1 and crLyme vaccines against Borreliella burgdorferi infection in the C3 H/HeN mouse needle challenge model.
Figure 3. Immunization with pLD1 confers protection against Borreliella burgdorferi bacterial challenge in C3 H/HeN mice: (a) Schematic of the experimental protocol pertaining to Borreliella burgdorferi bacterial challenge study. (b) Bacterial burden in joints, heart, bladder and ear was assessed by PCR in C3 H/HeN mice four weeks after the bacterial inoculation. Data show the mean ± SEM amount of Borreliella genome per 10,000 internal β-actin gene in indicated tissues. (c) Bacterial killing/growth inhibition assay was evaluated under dark-field microscope: 100,000 bacteria were incubated at 37°C in presence of serially diluted serum (see Material and methods) to assess the endpoint titer ± SEM at which bacterial growth is visible (n = 8 per group).
Assessment of the durability of the humoral immune response elicited by pLD1 vaccine in Hartley guinea pigs
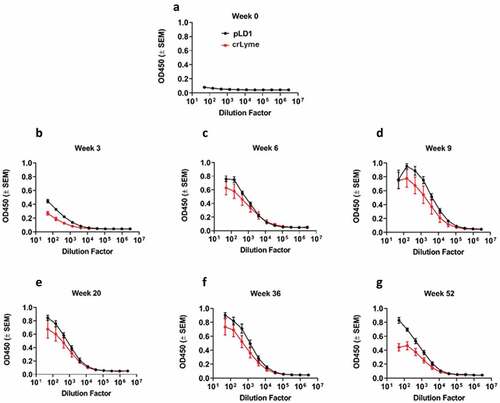
We assessed the immunogenicity of pLD1 in guinea pigs, an established model for intradermal vaccine delivery.Citation31,Citation32 We measured the OspA antigen-binding titers of IgG in the serum up to 52 weeks after initial vaccination. Animals in Group one received 0.1 mg of pLD1 vaccine via ID-CELLECTRA®EP delivery on weeks 0, 3 and 6. Group two received 250 µl (1/4 dose recommended for dogs) of crLyme vaccine administered subcutaneously at weeks 0, 3 and 6. Humoral immune responses against OspA antigen were measured up to Week 52. A robust humoral response against OspA was detected across both groups. We proceeded to evaluate the quality of the anti-OspA humoral immune response by assessing the generation of antibodies targeting the LA-2 epitope. To accomplish this we used the LA-2 equivalent mAb 319–44 in a competition assay with the serum from immunized guinea pigs.Citation20 Similar assays have previously been shown to be a pre-clinical correlate of protection.Citation33 The humoral response elicited by pLD1 and crLyme vaccines demonstrated the ability to compete and inhibit the binding of human mAb 319–44 (). These results underscore the potential of the pLD1 vaccine to provide durable antibody titers to OspA epitopes associated with correlates of protection.
Discussion
Previous studies employing DNA vaccines encoding OspA have shown promise in small animal studies,Citation34–36 however they have yet to advance to clinical testing. Here we have employed advanced delivery technology of in vivo EP to enhance the immunogenicity of the Lyme disease vaccine pLD1 after administration to the C3 H/HeN mouse muscle or Hartley guinea pig skin. Such technology increases in vivo gene expression up to 1000 fold, and the subsequent cellular and humoral immune responses elicited are substantively enhanced.Citation37 Here, the described study is the first to employ this vaccine delivery technology as a development concept for Lyme disease prevention. We show that a synthetically engineered vaccine pLD1 elicits robust and durable anti-OspA IgG levels, and confers protection against Borreliella burgdorferi infection in a C3 H/HeN mouse needle challenge model.
The pLD1 vaccine induced a robust anti-OspA immune response in C3 H/HeN mice (). The ability to induce strong T cell immunity is likely due to the intracellular antigen expression pathway associated with DNA vaccines, assuring antigen processing in the MHC class I and II pathways, driving broad T cell responses. Furthermore, the activation of Toll-like receptors by the un-methylated CpG motifs contained within synthetic nucleotides, and the activation of the intracellular STING pathway may provide innate signals that lead to enhanced cellular responses.Citation38,Citation39 Activated T cells can provide help to B cells leading to increased antibody levels and memory cell development, enhancing the humoral immunity. In reference to Lyme disease immune responses, an OspA T cell epitope that promotes OspA antibody response has been described,Citation18 supporting the importance of anti-OspA T cell responses in promoting antibody production.Citation40 Data here show one dose of pLD1 generates high antibody titers (see ,)), enough to provide protection against bacterial challenge ()).
We recently reported intradermal delivery of an Ebola GP Synthetic DNA vaccine resulted in 100% seroconversion and induction of T cell responses in 70% of subjects within weeks, by simple dose sparing ID formulations.Citation13 In the current study, we evaluated the immunogenicity of the pLD1 DNA vaccine in guinea pigs employing intradermal delivery. Guinea pigs represent an excellent ID immunization model, because unlike mice they possess a well-defined epidermis. An important aspect of targeting vaccine delivery to the skin is that it is rich in professional antigen-presenting cells that are capable of mediating host immune responses. ID delivery of pLD1 elicited a robust humoral immune response (). One limitation in this current study was that it did not consider the role of OspC-targeted immune responses of the comparator vaccine. crLyme is a chimeric vaccine containing OspA ST1 and a chimeric protein of multiple OspC antigens. Unlike OspA, OspC is not downregulated and is present on spirochetes entering the host, and vaccines containing OspC antigens may have the ability to elicit responses to spirochetes which have entered the host.Citation41 crLyme has a USDA-approved 15-month duration of immunity on its label. In contrast to OspA, over 30 different OspC genotypes have been defined, with many circulating in the USA.Citation42 The inclusion of OspC antigens may be considered in formulations.
Figure 4. Robust and sustained humoral response after intradermal immunization with pLD1 in Hartley guinea pigs. Guinea pigs were immunized at three-week intervals with pLD1 or the crLYME vaccine as described in materials and methods. (a–g) OspA antigen binding of IgG in serial serum dilutions from guinea pigs. Data shown represent OD450 nm values (± SEM) for each group from week 0 to week 52 (n = 5 per group).
The ability of serum from OspA-immunized guinea pigs to competitively inhibit the binding of OspA specific-319-44 mAb was also evaluated (). This type of competition assay can serve as a correlate of protective anti-OspA response as previously shown for the murine LA-2 antibody.Citation27,Citation33 The LA-2 antibody has been identified as protective against Borreliella burgdorferi infection in passive transfer and active immunization studies targeting OspA protein.Citation33,Citation43,Citation44 319–44 mAb has demonstrated the equivalent capability of mediating protection against Borreliella burgdorferi infection in antibody transfer experiments as the LA-2 mAb.Citation19,Citation20 In the context of the present study, antibodies from sera of pLD1 immunized guinea pigs provided inhibition of 319–44 mAb OspA binding. These data indicate the immune response elicited by pLD1 can generate antibodies which can bind to the same OspA epitopes as mAbs which have been shown to prevent transmission of Lyme disease spirochetes.
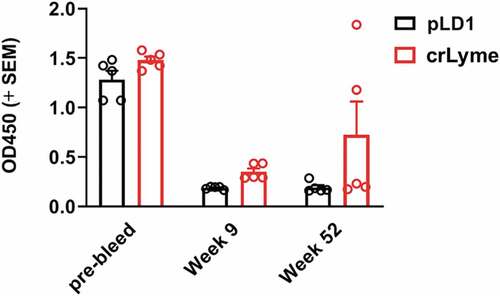
Figure 5. Immunization with pLD1 elicits humoral response against LA-2 epitope. A competitive inhibition ELISA was performed with serum from guinea pigs (dilution 1:50, n = 5 per group for Pre-bleed, week 9 & 52) and LA-2 binding mAb 319–44. O.D values for 319–44 mAb binding to OspA in presence of guinea pig sera from immunized animals.
While there is an obvious unmet need for a Lyme disease vaccine, it is important to consider the hurdles faced by previous vaccine programs which were ultimately not successful. The efficacy of this recombinant protein OspA vaccine was only 49% after 2 immunizations and required yearly boosts to maintain protective antibody titers. Furthermore, there was speculation about side effects due to the vaccine containing an epitope with homology to a human epitope in the LFA-1 protein. With pLD1 we have aimed to enhance the durability of the antibody titers which would potentially result a reduction in the frequency of boosts required to maintain protective serum titers. Additionally, we replaced the epitope with homology to human LFA-1 ()). Preliminary surveys are ongoing to assess the current market for a Lyme disease vaccine. With the increasing incidence and impact of this disease, we are optimistic about the demand and the level of utilization of such a medical countermeasure. Future studies are planned to address the limitations of the scope of work presented here. These will focus on efficacy testing in Borreliella burgdorferi infected tick challenges, which will reflect a more natural exposure route than the bacterial needle challenge model reported here. Studies will be designed to test vaccine efficacy after tick challenge in the dog, a large animal model which exhibits similarities in LD pathophysiology with humans.
Author contributions
GHG designed, analyzed and performed experiments, prepared manuscript; BS designed experiments, analyzed experiments, revised manuscript; HB designed and performed experiments; NC, JC, JY and CR designed DNA constructs and experiments; MM and BY performed experiments; RNE designed experiments; HP performed experiments; DBW revised manuscript, provided supervision; LHR provided supervision and revised manuscript; KEB provided supervision, designed experiments and revised manuscript; TRFS designed experiments, reviewed data, revised manuscript, provided supervision.
Disclosure of Potential Conflicts of Interest
GHG, HB, NC, JC, JY, CH, BY, HP, LH, KEB and TRFS are employees of Inovio Pharmaceuticals and as such receive salary and benefits, including ownership of stock and stock options, from the company. DBW has received grant funding, participates in industry collaborations, has received speaking honoraria, and has received fees for consulting, including serving on scientific review committees and board services. Remuneration received by DBW includes direct payments or stock options, and in the interest of disclosure he notes potential conflicts associated with this work with Inovio and possibly others. In addition, he has a patent DNA vaccine delivery pending to Inovio. All other authors report there are no competing interests.
Acknowledgments
We thank Maria Yang and team for the manufacturer of plasmid DNA vaccines.
Additional information
Funding
References
- Steere AC, Coburn J, Glickstein L. The emergence of lyme disease. J Clin Invest. 2004;113(8):1093–101. doi:10.1172/JCI21681.
- Stanek G, Wormser GP, Gray J, Strle F. Lyme borreliosis. Lancet. 2012;379(9814):461–73. doi:10.1016/S0140-6736(11)60103-7.
- Rosenberg R, Lindsey NP, Fischer M, Gregory CJ, Hinckley AF, Mead PS, Paz-Bailey G, Waterman SH, Drexler NA, Kersh GJ, et al. Vital signs: trends in reported vectorborne disease cases - United States and territories, 2004–016. MMWR Morb Mortal Wkly Rep. 2018;67(17):496–501. doi:10.15585/mmwr.mm6717e1.
- Nelson CA, Saha S, Kugeler KJ, Delorey MJ, Shankar MB, Hinckley AF, Mead PS. Incidence of clinician-diagnosed lyme disease, United States, 2005–2010. Emerg Infect Dis. 2015;21(9):1625–31. doi:10.3201/eid2109.150417.
- Steere AC, Angelis SM. Therapy for Lyme arthritis: strategies for the treatment of antibiotic-refractory arthritis. Arthritis Rheum. 2006;54(10):3079–86. doi:10.1002/art.22131.
- Fikrig E, Barthold SW, Kantor FS, Flavell RA. Long-term protection of mice from Lyme disease by vaccination with OspA. Infect Immun. 1992;60(3):773–77. doi:10.1128/IAI.60.3.773-777.1992.
- Fikrig E, Telford SR, Barthold SW, Kantor FS, Spielman A, Flavell RA. Elimination of Borrelia burgdorferi from vector ticks feeding on OspA-immunized mice. Proc Natl Acad Sci USA. 1992;89(12):5418–21. doi:10.1073/pnas.89.12.5418.
- Embers ME, Narasimhan S. Vaccination against Lyme disease: past, present, and future. Front Cell Infect Microbiol. 2013;3:6. doi:10.3389/fcimb.2013.00006.
- Philipp MT, Lobet Y, Bohm RP Jr, Roberts ED, Dennis VA, Gu Y, Lowrie RC Jr, Desmons P, Duray PH, England JD, et al. The outer surface protein A (OspA) vaccine against Lyme disease: efficacy in the rhesus monkey. Vaccine. 1997;15(17–18):1872–87. doi:10.1016/S0264-410X(97)00133-3.
- Sigal LH, Zahradnik JM, Lavin P, Patella SJ, Bryant G, Haselby R, Hilton E, Kunkel M, Adler-Klein D, Doherty T, et al. A vaccine consisting of recombinant borrelia burgdorferi outer-surface protein A to prevent lyme disease. Recombinant outer-surface protein A Lyme disease. N Engl J Med. 1998;339(4):216–22. doi:10.1056/NEJM199807233390402.
- Nigrovic LE, Thompson KM. The Lyme vaccine: a cautionary tale. Epidemiol Infect. 2007;135(1):1–8. doi:10.1017/S0950268806007096.
- Steere AC, Drouin EE, Glickstein LJ. Relationship between immunity to Borrelia burgdorferi outer-surface protein A (OspA) and Lyme arthritis. Clin Infect Dis. 2011;52(Suppl 3):s259–65. doi:10.1093/cid/ciq117.
- Tebas P, Kraynyak KA, Patel A, Maslow JN, Morrow MP, Sylvester AJ, Knoblock D, Gillespie E, Amante D, Racine T, et al. Intradermal SynCon(R) Ebola GP DNA vaccine is temperature stable and safely demonstrates cellular and humoral immunogenicity advantages in healthy volunteers. J Infect Dis. 2019;220:400–10. doi:10.1093/infdis/jiz132.
- Tebas P, Roberts CC, Muthumani K, Reuschel EL, Kudchodkar SB, Zaidi FI, White S, Khan AS, Racine T, Choi H, et al. Safety and immunogenicity of an Anti-Zika virus DNA vaccine - preliminary report. N Engl J Med. 2017. doi:10.1056/NEJMoa1708120.
- Modjarrad K, Roberts CC, Mills KT, Castellano AR, Paolino K, Muthumani K, Reuschel EL, Robb ML, Racine T, Oh M-D, et al. Safety and immunogenicity of an anti-Middle East respiratory syndrome coronavirus DNA vaccine: a phase 1, open-label, single-arm, dose-escalation trial. Lancet Infect Dis. 2019;19:1013–22. doi:10.1016/S1473-3099(19)30266-X.
- Morrow MP, Kraynyak KA, Sylvester AJ, Shen X, Amante D, Sakata L, Parker L, Yan J, Boyer J, Roh C, et al. Augmentation of cellular and humoral immune responses to HPV16 and HPV18 E6 and E7 antigens by VGX-3100. Mol Ther Oncol. 2016;3:16025. doi:10.1038/mto.2016.25.
- Trimble CL, Morrow MP, Kraynyak KA, Shen X, Dallas M, Yan J, Edwards L, Parker RL, Denny L, Giffear M, et al. Safety, efficacy, and immunogenicity of VGX-3100, a therapeutic synthetic DNA vaccine targeting human papillomavirus 16 and 18 E6 and E7 proteins for cervical intraepithelial neoplasia 2/3: a randomised, double-blind, placebo-controlled phase 2b trial. Lancet. 2015;386(10008):2078–88. doi:10.1016/S0140-6736(15)00239-1.
- Bockenstedt LK, Fikrig E, Barthold SW, Flavell RA, Kantor FS. Identification of a Borrelia burgdorferi OspA T cell epitope that promotes anti-OspA IgG in mice. J Immunol. 1996;157:5496–502.
- Wang Y, Esquivel R, Flingai S, Schiller ZA, Kern A, Agarwal S, Chu J, Patel A, Sullivan K, Wise MC, et al. Anti-OspA DNA-encoded monoclonal antibody prevents transmission of spirochetes in tick challenge providing sterilizing immunity in mice. J Infect Dis. 2019;219(7):1146–50. doi:10.1093/infdis/jiy627.
- Wang Y, Kern A, Boatright NK, Schiller ZA, Sadowski A, Ejemel M, Souders CA, Reimann KA, Hu L, Thomas WD, et al. Pre-exposure prophylaxis with OspA-specific human monoclonal antibodies protects mice against tick transmission of lyme disease spirochetes. J Infect Dis. 2016;214(2):205–11. doi:10.1093/infdis/jiw151.
- Andre S, Seed B, Eberle J, Schraut W, Bültmann A, Haas J. Increased immune response elicited by DNA vaccination with a synthetic gp120 sequence with optimized codon usage. J Virol. 1998;72(2):1497–503. doi:10.1128/JVI.72.2.1497-1503.1998.
- Deml L, Bojak A, Steck S, Graf M, Wild J, Schirmbeck R, Wolf H, Wagner R. Multiple effects of codon usage optimization on expression and immunogenicity of DNA candidate vaccines encoding the human immunodeficiency virus type 1 Gag protein. J Virol. 2001;75(22):10991–1001. doi:10.1128/JVI.75.22.10991-11001.2001.
- Sahay B, Patsey RL, Eggers CH, Salazar JC, Radolf JD, Sellati TJ. CD14 signaling restrains chronic inflammation through induction of p38-MAPK/SOCS-dependent tolerance. PLoS Pathog. 2009;5(12):e1000687. doi:10.1371/journal.ppat.1000687.
- Benhnia MR, Wroblewski D, Akhtar MN, Patel RA, Lavezzi W, Gangloff SC, Goyert SM, Caimano MJ, Radolf JD, Sellati TJ, et al. Signaling through CD14 attenuates the inflammatory response to Borrelia burgdorferi, the agent of lyme disease. J Immunol. 2005;174(3):1539–48. doi:10.4049/jimmunol.174.3.1539.
- Liang FT, Yan J, Mbow ML, Sviat SL, Gilmore RD, Mamula M, Fikrig E. Borrelia burgdorferi changes its surface antigenic expression in response to host immune responses. Infect Immun. 2004;72(10):5759–67. doi:10.1128/IAI.72.10.5759-5767.2004.
- Morrison TB, Ma Y, Weis JH, Weis JJ. Rapid and sensitive quantification of Borrelia burgdorferi-infected mouse tissues by continuous fluorescent monitoring of PCR. J Clin Microbiol. 1999;37(4):987.
- Willett TA, Meyer AL, Brown EL, Huber BT. An effective second-generation outer surface protein A-derived Lyme vaccine that eliminates a potentially autoreactive T cell epitope. Proc Natl Acad Sci USA. 2004;101(5):1303–08. doi:10.1073/pnas.0305680101.
- Bodles-Brakhop AM, Draghia-Akli R. DNA vaccination and gene therapy: optimization and delivery for cancer therapy. Expert Rev Vaccines. 2008;7(7):1085–101. doi:10.1586/14760584.7.7.1085.
- Moody KD, Barthold SW. Lyme borreliosis in laboratory mice. Lab Anim Sci. 1998;48:168–71.
- Miller JC, Ma Y, Crandall H, Wang X, Weis JJ. Gene expression profiling provides insights into the pathways involved in inflammatory arthritis development: murine model of Lyme disease. Exp Mol Pathol. 2008;85(1):20–27. doi:10.1016/j.yexmp.2008.03.004.
- Schultheis K, Schaefer H, Yung BS, Oh J, Muthumani K, Humeau L, Broderick KE, Smith TRF. Characterization of guinea pig T cell responses elicited after EP-assisted delivery of DNA vaccines to the skin. Vaccine. 2017;35(1):61–70. doi:10.1016/j.vaccine.2016.11.052.
- Smith TRF, Schultheis K, Morrow MP, Kraynyak KA, McCoy JR, Yim KC, Muthumani K, Humeau L, Weiner DB, Sardesai NY, et al. Development of an intradermal DNA vaccine delivery strategy to achieve single-dose immunity against respiratory syncytial virus. Vaccine. 2017;35(21):2840–47. doi:10.1016/j.vaccine.2017.04.008.
- Golde WT, Piesman J, Dolan MC, Kramer M, Hauser P, Lobet Y, Capiau C, Desmons P, Voet P, Dearwester D, et al. Reactivity with a specific epitope of outer surface protein A predicts protection from infection with the Lyme disease spirochete, Borrelia burgdorferi. Infect Immun. 1997;65(3):882–89. doi:10.1128/IAI.65.3.882-889.1997.
- Simon MM, Gern L, Hauser P, Zhong W, Nielsen PJ, Kramer MD, Brenner C, Wallich R. Protective immunization with plasmid DNA containing the outer surface lipoprotein A gene of Borrelia burgdorferi is independent of an eukaryotic promoter. Eur J Immunol. 1996;26(12):2831–40. doi:10.1002/eji.1830261206.
- Luke CJ, Carner K, Liang X, Barbour AG. An OspA-based DNA vaccine protects mice against infection with Borrelia burgdorferi. J Infect Dis. 1997;175(1):91–97. doi:10.1093/infdis/175.1.91.
- Wallich R, Siebers A, Jahraus O, Brenner C, Stehle T, Simon MM. DNA vaccines expressing a fusion product of outer surface proteins A and C from Borrelia burgdorferi induce protective antibodies suitable for prophylaxis but Not for resolution of Lyme disease. Infect Immun. 2001;69(4):2130–36. doi:10.1128/IAI.69.4.2130-2136.2001.
- Sardesai NY, Weiner DB. Electroporation delivery of DNA vaccines: prospects for success. Curr Opin Immunol. 2011;23(3):421–29. doi:10.1016/j.coi.2011.03.008.
- Bode C, Zhao G, Steinhagen F, Kinjo T, Klinman DM. CpG DNA as a vaccine adjuvant. Expert Rev Vaccines. 2011;10(4):499–511. doi:10.1586/erv.10.174.
- Klinman DM, Klaschik S, Sato T, Tross D. CpG oligonucleotides as adjuvants for vaccines targeting infectious diseases. Adv Drug Deliv Rev. 2009;61(3):248–55. doi:10.1016/j.addr.2008.12.012.
- Elsner RA, Hastey CJ, Baumgarth N. CD4+ T cells promote antibody production but not sustained affinity maturation during Borrelia burgdorferi infection. Infect Immun. 2015;83(1):48–56. doi:10.1128/IAI.02471-14.
- Izac JR, Marconi RT. Diversity of the lyme disease spirochetes and its influence on immune responses to infection and vaccination. Vet Clin North Am Small Anim Pract. 2019;49(4):671–86. doi:10.1016/j.cvsm.2019.02.007.
- Rhodes DV, Earnhart CG, Mather TN, Meeus PFM, Marconi RT. Identification of Borrelia burgdorferi ospC genotypes in canine tissue following tick infestation: implications for Lyme disease vaccine and diagnostic assay design. Vet J. 2013;198(2):412–18. doi:10.1016/j.tvjl.2013.07.019.
- Johnson BJ, Sviat SL, Happ CM, Dunn JJ, Frantz JC, Mayer LW, Piesman J. Incomplete protection of hamsters vaccinated with unlipidated OspA from Borrelia burgdorferi infection is associated with low levels of antibody to an epitope defined by mAb LA-2. Vaccine. 1995;13(12):1086–94. doi:10.1016/0264-410X(95)00035-Y.
- Schaible UE, Kramer MD, Eichmann K, Modolell M, Museteanu C, Simon MM. Monoclonal antibodies specific for the outer surface protein A (OspA) of Borrelia burgdorferi prevent Lyme borreliosis in severe combined immunodeficiency (scid) mice. Proc Natl Acad Sci USA. 1990;87(10):3768–72. doi:10.1073/pnas.87.10.3768.