
Abstract
Alginate lyases catalyze the degradation of alginate, a complex copolymer of α-L-guluronate and its C5 epimer β-D-mannuronate. The enzymes have been isolated from various kinds of organisms with different substrate specificities, including algae, marine mollusks, marine and terrestrial bacteria, and some viruses and fungi. With the progress of structural biology, many kinds of alginate lyases of different polysaccharide lyases families have been characterized by obtaining crystal structures, and the catalytic mechanism has also been elucidated. Combined with various studies, we summarized the source, classification and properties of the alginate lyases from different polysaccharide lyases families. The relationship between substrate specificity and protein sequence was also investigated.
Introduction
Alginate is the most abundant polysaccharide (about 40% of dry weight) of brown algae, which consists of β-D-mannuronate (M) and α-L-guluronate (G) as monomeric units.Citation1 These units are linked in 3 kinds of different blocks, poly β-D-mannuronate (polyM), poly α-L-guluronate (polyG) and the heteropolymer (polyMG).Citation2 Some bacteria can also synthesize alginates to protect them from detrimental factors such as antibiotics and drought.Citation3,4 Commercial alginates are produced by extraction from biomass of brown algae such as Laminaria hyperborea, Macrocystis pyrifera, Laminaria japonica and etc. Alginate oligosaccharides are depolymerization products of alginate by alginate lyase or physicochemical method. They have attracted increasing attention due to their wide applications in food and pharmaceutical industry.Citation5-7 They can be used as growth promoters for plants and therapeutic agents such as anticoagulants and tumor inhibitors.Citation8-10 They can also induce the cytokine production and regulate the blood sugar as well as lipid.Citation11,12 Due to the high-efficiency and high-specificity, alginate lyases for mild degradation have been the focus for various fields.
Alginate lyase can degrade alginate by β-elimination of glycosidic bonds and produce unsaturated oligosaccharides with double bonds at the non-reducing end.Citation13 A number of alginate lyases have been identified, cloned, purified and characterized from various sources,Citation14-18 including marine and terrestrial bacteria, marine mollusks and algae. They can be classed into 2 groups due to their substrate specificities,Citation13 molecular weightsCitation19 and action modes. Now alginate lyases, especially endolytic alginate lyases, have been widely used in the production of alginate oligosaccharides,Citation20 the elucidation of the fine structures of alginateCitation21-23 and the preparation of protoplast of red and brown algae.Citation24-26 Furthermore, alginate lyase also shows great potential application in treatment of cystic fibrosis by degrading the polysaccharide biofilm of bacterium.Citation27-29
Source and Classification
Alginate lyases have been isolated from various sources, including marine algae, marine mollusks (Littorina spp., Haliotis spp, Turbo cornutus.), and a wide range of marine and terrestrial bacteria. In addition, some lyases have been isolated from fungi and viruses (). The alginate lyases can be classified into 2 groups due to their substrate specificities, one is G block-specific lyase (polyG lyase, EC4.2.2.11), and the other is M block-specific lyase (polyM lyase, EC4.2.2.3). This classification has been widely accepted, but some enzymes show activities toward both polyM and polyG,Citation30-33 which may degrade alginate more effectively. In terms of the mode of action, alginate lyase can be grouped into endolytic and exolytic alginate lyase.Citation13 Endolytic alginate lyase cleaves glycosidic bonds inside alginate polymer and releases unsaturated oligosaccharides (di-, tri-, and tetra-saccharides) as main products,Citation30 while exolytic alginate lyase can further degrade oligosaccharides into monomers.Citation31-33 Based on the analysis of hydrophobic cluster of primary structures, alginate lyases can be grouped into 7 families of Polysaccharide Lyase (PL) family (), PL-5, -6, -7, -14,-15, -17, and -18.Citation34 Most of endolytic bacterial alginate lyases are assigned to PL-5 and PL-7. The most exolytic alginate lyases are grouped into PL-15 and PL-17 families. Most alginate lyases from bacteria are assigned into PL-5, -7, -15, and -17 families. The lyases isolated from marine mollusks and viruses are collected in PL-14 family. The bifunctional alginate lyases belong to PL-18 family, while other lyases are dispersed in other 6 families. According to the 3-dimensional structures, the alginate lyase are grouped into 3 families,Citation35 including parallel β-helix family, (α/α)6 barrel family, jelly-roll family. The alginate lyases of PL-6 belong to parallel β-helix family,Citation36 the PL-5 and -15 members are grouped into (α/α) 6 barrel family,Citation37,38 while -7 and -14 are assigned into jelly-roll family.Citation39,40 The bifunctional alginate lyases of PL-18 are newly characterized and shared a similar sandwich structure.Citation41 Furthermore,the alginate lyases can also be grouped into 3 types based on their molecular masses: small (25–30 kDa), medium-sized (around 40 kDa), and large lyases (>60 kDa).
Table 1. Alginate lyase of different PL families from different sources and their substrate specificities
Table 2. Characteristics of alginate lyases from different PL families
Structure and Mechanism of Action
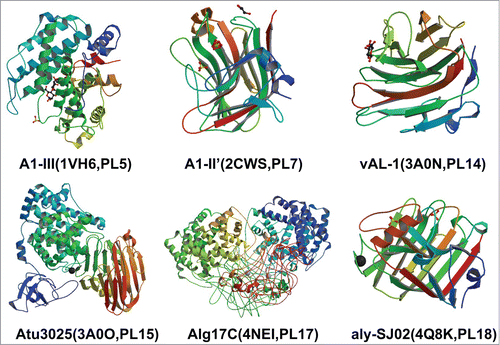
The three-dimensional structures of various alginate lyases have been elucidated in . The structure of alginate lyase A1-III from Sphingomonas sp. A1 has been resolved complexed with a trisaccharide product.Citation37 The overall structure of A1-III is abundant in helixes and has a deep tunnel-like cleft in a novel (α/α)-barrel structure as the catalytic active domain. The structure presented the possibility that alginate molecules might penetrate into the cleft to interact with the catalytic site of A1-III. This is the only alginate lyase with resolved structure in PL-5. As for PL-7, there are 5 alginate lyases with resolved structures.Citation19,38,Citation42-44 The overall structures of these alginate lyases are all β-sandwich fold like structure with a large active cleft covered by 2 short flexible loops. The overall structure of alginate lyase from Chlorella virus for PL-14 show 2 antiparallel β-sheets with a deep cleft showing a β-jelly roll fold.Citation40 The alginate lyases of PL-17 are known as exolytic alginate lyases and only one has resolved structure with a combination of (α/α) barrel and β-sandwich fold.Citation45 The arrangement of (α/α) helixes produces an open barrel structure and its additional helix serves to maintain rigidity of the barrel. The β-sandwich fold domain is composed of 3 co-planer layers of antiparallel β-strands arranged as sheets and further supported by 4 small helices near the top layer, which interject between the 2 domains. The alginate lyase Aly-SJ02 of PL-18 displays a β-jelly roll scaffold composed mainly of 2 anti-parallels β-sheets.Citation41
Figure 1. The overall structures of alginate lyases from different families.Citation41
Alginate lyase catalyzes the degradation of alginate by β-elimination mechanism, breaking the glycosidic bond between monomers and producing a resulted double bond between the C4 and C5 carbons of the sugar rings. During the elimination, the 4-O-glycosidic bond is eliminated and simultaneously yields oligosaccharides containing 4-deoxy-L-erythro-hex-4-enopyranosyluronic acid as the nonreducing terminal moiety.Citation11 With the 3-dimensional structures of several alginate lyases been resolved,Citation41,45,46 the catalytic mechanism have been elucidated to be 3 steps. The first step is the removal of the negative charge on the carboxyl anion-essentially neutralizing the charge by a salt bridge (Histidine or Lysine) and then a general base-catalyzed abstraction of the proton on C5, where one residue may be required as the proton abstractor and another as the proton donor or the residue act as both proton donor and acceptor. Finally, a transfer of electrons from the carboxyl group forms a double bond between C4 and C5, resulting in the elimination of the 4-O-glycosidic bond.
Substrate Specificity
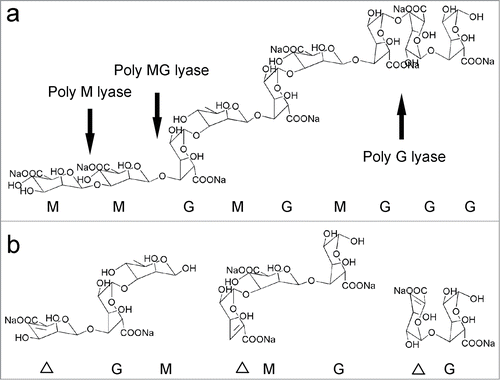
Alginates are composed of 4 different types of linkage such as M-M, M-G, G-G, and G-M with various extent of each linkage (). Alginate lyases are classified based on their dominant cleaving action on the different types of substrates as shown in Citation47. The alginate lyases preferring M-rich alginates are assigned into polyM lyases (EC 4. 2. 2. 3), while lyases preferring G-rich alginates are grouped into polyG lyases (EC 4. 2. 2. 11). Although an alginate lyase may be named as polyM lyase or polyG lyase, the enzyme usually displays moderate to low processivity for the other homopolymer. This confusing variable may result from the quality and purity of substrate used for these determinations. These concerns are currently being addressed by using the substrate with high purity and highly sensitive and accurate techniques to analyze the structure of the products. There are several reports of alginate lyases with specific substrate specificities, such as the lyase from ATCC43367 which is reported to cleave only M-M linkages.Citation48 An alginate lyase isolated from marine mollusk Lambis sp displays 100% of activity for polyG while 0% of activity for polyM.Citation49 What is more, an alginate lyase from Klebsiella pneumonia showing activities toward G-blocks and MG-blocks is modified with cleavage specificity for G-G linkage by site-mutagenesis.Citation50 However, some alginate lyases show activities on both polyM and polyG and are regarded as bifunctional lyases, such as alginate lyase AlySJ-02 from Pseudoalteromonas sp. SM0524, AlyPEEC from Pseudoalteromonas sp IAM14594, Aly from Pseudoalteromonas sp. 272, and AlyA from Pseudoalteromonas atlantica AR06.
Figure 2. The substrate specificity of alginate lyase and structures of degradation products. The polysaccharide alginate containing 3 kinds of blocks (M, G, MG) is cleaved to produce a 4-deoxy-L-erythro-hex-4- enepyranosyluronate moiety (open triangle) at the newly formed non-reducing end of the product.Citation47
Relationship Between Substrate Specificity and Structure
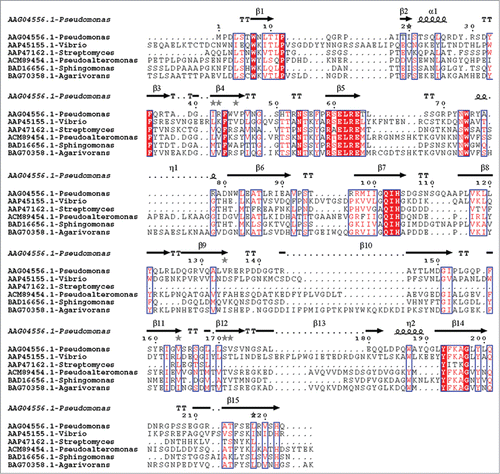
The alginate lyases of PL7 family have been well investigated. There are 25 characterized alginate lyases in PL-7 family, and 5 of them have been investigated for structure-function relationship by obtaining crystal structures. The structural basis for depolymerization of alginate lyase has been elucidated.Citation51 As shown in , the alginate lyases contain 3 highly conserved regions, (R/E) (S/T/N) EL, Q (I/V) H, YFKAG (V/I) YNQ. According to the crystal structures and the sequence analysis of several PL family 7 lyases, those conserved regions form the cavity composed of a jelly-roll β-sandwich structure, and the cavity is assumed to bind a suitable substrate. Thus, conserved amino acids residues are thought to play an important role in catalytic activity or folding of the structure. It has been reported that Y195, H119, Q117 and R72 may be involved in the catalytic site for ALY-1.Citation52 Therefore, the amino acids in catalytic and substrate binding sites would be highly conserved depending on the substrate specificity. The polyM-specific alginate lyases contain QVH in conserved regions and polyG-specific enzymes involve QIH in conserved regions, while the polyMG lyases include QIH in the conserved regions. In addition, the substitution of hydrophobic residues in the isoleucine site of domain QIH could have enormous influence on the high affinity to polyG block. For this perspective, the substrate specificity is determined by the isoleucine of QIH domain and valine of QVH domain. The isoleucine of QIH domain is confirmed to be indispensible for reorganization of the polyG or G-G bond.Citation53 So, if an alginate lyase with unknown substrate specificity contained isoleucine or valine in Q (I/V) H region, this enzyme could degrade polyM or polyG and polyMG, respectively. The region YFKAG (V/I) YNQ has been reported from various alginate lyases with different specificities, indicating that this region may be of functional or structural importance for alginate degradation independent of substrate preference. Considering that the alginate lyase totally lost activity with deletion of this region, the results suggest that the region may be needed to catalyze alginate degradation.
Figure 3. Multiple alignment of amino acid sequences of alginate lyases of PL 7 family. The conserved residues were overlaid with red and 3 highly conserved regions-(R/E) (S/T/N) EL, Q (I/V) H, YFKAG (V/I) YNQ were boxed in blue.
Functionality and Promising Application of Alginate Lyase
Depolymerized alginate produced by enzymatic hydrolysis with low DP possesses various kinds of biological activities.Citation8-14 Oligomeric alginates obtained from lyase degradation of alginate act as oligosaccharines and regulate physiological processes in plants and microorganisms, such as promoting the growth of Bifidobacterium spp., enhancing the germination and shoot elongation in plants. The oligosaccharides exhibit many physiological activities, such as antitumor, stimulating production of cytokines, regulating the blood lipids and sugars. The efficient alginate lyases w are key tools for production of functional oligosaccharides from alginate.
The alginate lyases with different substrate specificities are invaluable tools for determining sequence of substrate and preparing the oligosaccharides with specific structures. Alginate lyases possess different substrate specificities depending on the differences in amino acids and distributing of monosaccharide residues in substrate. Various enzymes could recognize 4 different types of linkages such as M-M, M-G, G-M, and G-G. The alginate lyases of PL-5 family prefer to degrade polyM substrate. The AlgA from Pseudomonas sp E03 hydrolyzes the polyM substrate in an endolytic manner and releases a range of oligomannuronate with low DP.Citation11 The AlxM from Photobacterium sp. ATCC43367 is also a polyM-specific alginate lyase.Citation48 And a polyM lyase from Sargassum fluitans specifically depolymerize polyM.Citation54 The alginate lyases specific for G-G linkage have not been described except for a mutant enzyme with cleavage specificity for G-G linkage.Citation14 This G-G linkage specific enzyme is constructed by mutating the gene encoding Klebsiella pneumonia AlyA, which cleaves both G-G and G-M linkages. The enzyme isolated from Klebsiella pneumonia shows activity only toward G-G linkage.Citation50 The polyMG-specific alginate lyase from Stenotrophomas maltophilia KJ-2 preferably degrades the glycosidic bond in M-G linkage than that in G-M linkage.Citation55 The alginate lyases have been used to analyze alginate fine structure to understand how chemical composition influences the physical properties of alginate. The combination of alginate lyases with different substrate specificities has used for determining the sequence and block distribution of alginate.Citation56
In addition, the enzymes with strict substrate specificities could be used for preparation alginate with specific structures. The AlxMB can be used for preparation of homopolymannuronate blocks and strictly alternating sequences of mannuronic and guluronic acids due to its strict substrate specificity toward polyM blocks.Citation57 A simple method for preparation of polyG blocks has been developed by using a polyG lyase from Flavobacterium multivolum.Citation58 The enzyme could degrade both G- and MG- blocks but M blocks. The guluronan lyase from Klebsiella aeruginosa has no action on polyM, but can readily cleave G-G linkages in guluronan. Based on the substrate specificity of this enzyme, it has been used for determining the activity of C-5 epimerase.Citation59
Recently, the alginate lyases have been used to treat the cystic fibrosis in combination with antibiotics.Citation27-29 When alginate lyase is co-administered with antibiotics such as gentamycin, the killing efficiency of mucoid Pseudomonas aeruginosa in respiratory tract increases. Therefore, the enzymes are expected to become useful agents for the treatment of bacterial mucoid biofilm-dependent diseases. What is more, the alginate lyases are also used to prepare the protoplast of algae for gene engineering and study Fucus cell wall development. With the consumption of fossil source, the alginate, being regarded as a renewable source for the production of bioethanol, has obtained increased attention. The saccharification of alginate for production of bioethanol requires synergistic effects of alginate lyases with endo- and exo- action modes. The engineered microbial platforms for direct production of bioethanol from alginate have been reported.Citation60-62
In conclusion, the enzymes are important tools in a broad spectrum of biological roles and applications. Alginate lyases with different substrate specificities can be used to produce oligosaccharides with various biological functionalities. Moreover, they can also be used for determination of the fine structure of alginate and the preparation of tailor-made alginate. Furthermore, alginate lyases with activities toward acetylated alginate can be utilized for the treatment of cystic fibrosis in combination with antibiotics. Therefore, due to the various activities and biological functionalities, alginate lyases have been widely applied. And with the continuing developments for screening and characterizing the enzymes, the applications of alginate lyases will become increasing wider in the near future.
Conclusion
The alginate lyases have important roles and biotechnological applications. Especially, the enzymes were key tools for production of oligosaccharides from alginate and determination of the fine structure of alginate. In addition, the alginate lyases play essential role in saccharification of the acidic polysaccharides for production of bioethanol with synergistic effect of endo- and exo-type alginate lyases. There were several hundred kinds of alginate lyases from various sources have been isolated and characterized. These enzymes differed from each other in substrate specificities, properties (including activities and structures) and degradation products. With the progress of structural biology, many kinds of alginate lyases of different PL families have been characterized by obtaining crystal structures, and the catalytic mechanism has also been elucidated. In future, screening and characterization of novel enzymes with high activities and broad substrate specificities would enhance and expand the utilization of the lyases to produce oligosaccharides with novel structures and biological activities for applications in various fields.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
The research in our lab mentioned in this paper was supported by the National Key Technology Support Program (2013BAB01B01), Special Fund for Marine Scientific Research (201305015-2), National Natural Science Foundation of China (31370811) and Development Fund for Collaborative Innovation Center of Glycoscience of Shandong University.
References
- Gacesa P. Enzymic degradation of alginates. Int J Biochem 1992; 24 545-52; PMID:1516726
- Arne H, Larsen B, Smidsrqd O. Studies on the sequence of uronic acid residues in alginic acid. Acta Chemica Scandinavica 1967; 21 691-704.
- Gimmestad M, Ertesvag H, Heggeset TMB, Aarstad O, Svanem BIG, Valla S. Characterization of Three New Azotobacter vinelandii Alginate Lyases, One of Which Is Involved in Cyst Germination. J Bacteriol 2009; 191 4845-53; PMID:19482920
- Jain S, Ohman DE. Role of an alginate lyase for alginate transport in mucoid Pseudomonas aeruginosa. Infect Immun 2005; 73:6429-36; PMID:16177314
- Fremond B, Malandain C, Guyomard C, Chesne C, Guillouzo A, Campion JP. Correction of Bilirubin Conjugation in the Gunn Rat Using Hepatocytes Immobilized in Alginate Gel Beads as an Extracorporeal Bioartificial Liver. Cell Transplant 1993; 2:453-60.
- SteinlechnerMaran R, Eberl T, Kunc M, Margreiter R, Gnaiger E. Oxygen dependence of respiration in coupled and uncoupled endothelial cells. Am J Physiol-Cell Ph 1996; 271:C2053-C2061; PMID:8997208
- Snyder H, Frey P, Thomas DFM, Caesar R, Palmer J, Atala A, Nasrallah P, Odonnell B, Aaronson I, Puri P, Endoscopic Treatment of Vesicoureteral Reflux with Chondrocyte-Alginate Suspension - Discussion. J Urology 1994; 152:644-4.
- Hu XK, Jiang XL, Hwang HM, Liu SL, Guan HS. Promotive effects of alginate-derived oligosaccharide on maize seed germination. J Appl Phycol 2004; 16:73-6; PMID:15692188
- Iwamoto M, Kurachi M, Nakashima T, Kim D, Yamaguchi K, Oda T, Iwamoto Y, Muramatsu T. Structure-activity relationship of alginate oligosaccharides in the induction of cytokine production from RAW264.7 cells. FEBS letters 2005; 579s) 4423-9; PMID:16055120
- An QD, Zhang GL, Wu HT, Zhang ZC, Zheng GS, Luan L, Murata Y, Li X. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp LXA and its potential application in protection against pathogens. J Appl Microbiol 2009; 106:161-70; PMID:19054241
- Zhang RX, Zhou JH, Ha ZP, Zhang YX, Gu GM. Hypoglycemic effect of Rehmannia glutinosa oligosaccharide in hyperglycemic and alloxan-induced diabetic rats and its mechanism. J Ethnopharmacol 2004; 90:39-43; PMID:14698506
- Zong W-R, Cheong K-L, Wu D-T, Li J, Zhao J, Li S-P. Preparation and purification of raffinose family oligosaccharides from Rehmannia glutinosa Libosch. by fast protein liquid chromatography coupled with refractive index detection. Separat Purificat Technol 2014; 138:98-103
- Wong TY, Preston LA, Schiller NL, ALGINATE LYASE: review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications, Annual Review Microbiol 2000; 54:289-340; PMID:11018131
- Kobayashi T, Uchimura K, Miyazaki M, Nogi Y, Horikoshi K, A new high-alkaline alginate lyase from a deep-sea bacterium Agarivorans sp, Extremophiles 2009; 13:121-9; PMID:19002649; http://dx.doi.org/10.1007/s00792-008-0201-7
- Suzuki H, Suzuki K, Inoue A, Ojima T, A novel oligoalginate lyase from abalone, Haliotis discus hannai, that releases disaccharide from alginate polymer in an exolytic manner, Carbohydr Res 2006; 341:1809-19; PMID:16697989
- Hashimoto W, Miyake O, Ochiai A, Murata K, Molecular identification of Sphingomonas sp. A1 alginate lyase (A1-IV') as a member of novel polysaccharide lyase family 15 and implications in alginate lyase evolution, J Biosci Bioeng 2005; 99:48-54; PMID:16233753
- Huang L, Zhou J, Li X, Peng Q, Lu H, Du Y. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. J Ind Microbiol Biotechnol 2013; 40:113-22; PMID:23111633
- Cao L, Xie L, Xue X, Tan H, Liu Y, Zhou S. Purification and characterization of alginate lyase from streptomyces species strain A5 isolated from banana rhizosphere. J Agricult Food Chem 2007; 55:5113-7; PMID:17536832
- Osawa T, Matsubara Y, Muramatsu T, Kimura M, Kakuta Y. Crystal structure of the alginate (poly alpha-l-guluronate) lyase from Corynebacterium sp. at 1.2 A resolution. J Mol Biol 2005; 345:1111-8; PMID:15644208
- Li L, Jiang X, Guan H, Wang P. Preparation, purification and characterization of alginate oligosaccharides degraded by alginate lyase from Pseudomonas sp. HZJ 216. Carbohydr Res 2011; 346:794-800; PMID:21371694; http://dx.doi.org/10.1016/j.carres.2011.01.023
- Aarstad OA, Tondervik A, Sletta H, Skjak-Braek G. Alginate sequencing: an analysis of block distribution in alginates using specific alginate degrading enzymes Biomacromolecules 2012; 13:106-16; PMID:22148348; http://dx.doi.org/10.1021/bm2013026
- Boyd J, Turvey JR, Structural studies of alginic acid, using a bacterial poly-alpha-l-guluronate lyase. Carbohydr Res 1978; 66:187-94.
- K. Ostgaard. Determination of alginate composition by a simple enzymatic assay. Hydrobiologia 1993; 261:513-20.
- Inoue A, Mashino C, Kodama T, Ojima T. Protoplast preparation from laminaria japonica with recombinant alginate lyase and cellulase. Marine Biotechnol 2011; 13:256-63; PMID:20393774
- Hu X, Jiang XL, Guan HS. Isolation of protoplasts from undaria pinnatifida by alginate lyase digestion. J Ocean U China 2003; 2:58-61
- Butler DM, Ostgaard K, Boyen C, Evans LV, Jensen A, Kloareg B. Isolation Conditions for High Yields of Protoplasts from Laminaria-Saccharina and Laminaria-Digitata (Phaeophyceae). J Exp Bot 1989; 40:1237-46
- Islan GA, Bosio VE, Castro GR. Alginate lyase and ciprofloxacin co-immobilization on biopolymeric microspheres for cystic fibrosis treatment. Macromol Biosci 2013; 13:1238-48; PMID:23966229
- Islan GA, Martinez YN, Illanes A, Castro G. Development of novel alginate lyase cross-linked aggregates for the oral treatment of cystic fibrosis. Rsc Adv 2014; 4:11758-65.
- Hatch RA, Schiller NL. Alginate lyase promotes diffusion of aminoglycosides through the extracellular polysaccharide of mucoid Pseudomonas aeruginosa. Antimicrob Agents ChemPeriodical 1998; 42:974-7; PMID:9559826
- Kim HT, Ko HJ, Kim N, Kim D, Lee D, Choi IG, Woo HC, Kim MD, Kim KH. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol Lett 2012; 34:1087-92; PMID:22391735; http://dx.doi.org/10.1007/s10529-012-0876-9
- Kim HT, Chung JH, Wang D, Lee J, Woo HC, Choi IG, Kim KH. Depolymerization of alginate into a monomeric sugar acid using Alg17C, an exo-oligoalginate lyase cloned from Saccharophagus degradans 2-40. Appl Microbiol Biotechnol 2012; 93:2233-9; PMID:22281843; http://dx.doi.org/10.1007/s00253-012-3882-x
- Park HH, Kam N, Lee EY, Kim HS. Cloning and characterization of a novel oligoalginate lyase from a newly isolated bacterium sphingomonas sp MJ-3. Marine Biotechnol 2012; 14:189-202; PMID:21826589; http://dx.doi.org/10.1007/s10126-011-9402-7
- Jagtap SS, Hehemann JH, Polz MF, Lee JK, Zhao HM. Comparative biochemical characterization of three exolytic oligoalginate lyases from vibrio splendidus reveals complementary substrate scope, temperature, and pH adaptations. Appl Environ Microbiol 2014; 80:4207-14; PMID:24795372
- Lombard V, Bernard T, Rancurel C, Brumer H, Coutinho PM, Henrissat B. A hierarchical classification of polysaccharide lyases for glycogenomics. Biochem J 2010; 432:437-44; PMID:20925655
- Hehemann JH, Boraston AB, Czjzek M. A sweet new wave: structures and mechanisms of enzymes that digest polysaccharides from marine algae. Curr Opin Struc Biol 2014; 28:77-86; PMID:25136767; http://dx.doi.org/10.1016/j.sbi.2014.07.009
- Huang WJ, Matte A, Li YG, Kim YS, Linhardt RJ, Su HS, Cygler M. Crystal structure of chondroitinase B from Flavobacterium heparinum and its complex with a disaccharide product at 1.7 angstrom resolution. J Mol Biol 1999; 294:1257-69; PMID:10600383
- Yoon HJ, Hashimoto W, Miyake O, Murata K, Mikami B. Crystal structure of alginate lyase A1-III complexed with trisaccharide product at 2.0 angstrom resolution. J Mol Biol 2001; 307:9-16; PMID:11243798
- Ochiai A, Yamasaki M, Mikami B, Hashimoto W, Murata K. Crystal structure of exotype alginate lyase Atu3025 from agrobacterium tumefaciens. J Biol Chem 2010; 285:24519-28; PMID:20507980
- Yamasaki M, Moriwaki S, Miyake O, Hashimoto W, Murata K, Mikami B. Structure and function of a hypothetical Pseudomonas aeruginosa protein PA1167 classified into family PL-7 - A novel alginate lyase with a beta-sandwich fold. J Biological Chem 2004; 279:31863-72; PMID:15136569
- Ogura K, Yamasaki M, Yamada T, Mikami B, Hashimoto W, Murata K. Crystal structure of family 14 polysaccharide Lyase with pH-dependent modes of action. J Biol Chem 2009; 284:35572-9; PMID:19846561
- Dong S, Wei TD, Chen XL, Li CY, Wang P, Xie BB, Qin QL, Zhang XY, Pang XH, Zhou BC, et al. Molecular insight into the role of the N-terminal extension in the maturation, substrate recognition, and catalysis of a bacterial alginate lyase from polysaccharide lyase family 18. J Biol Chem 2014; 289:29558-69; PMID:25210041
- Thomas F, Lundqvist LC, Jam M, Jeudy A, Barbeyron T, Sandstrom C, Michel G, Czjzek M. Comparative characterization of two marine alginate lyases from Zobellia galactanivorans reveals distinct modes of action and exquisite adaptation to their natural substrate. J Biol Chem 2013; 288:23021-37; PMID:23782694
- Yamasaki M, Ogura K, Hashimoto W, Mikami B, Murata K. A structural basis for depolymerization of alginate by polysaccharide lyase family-7. J Mol Biol 2005; 352:11-21; PMID:16081095
- Yamasaki M, Moriwaki S, Miyake O, Hashimoto W, Murata K, Mikami B. Structure and function of a hypothetical Pseudomonas aeruginosa protein PA1167 classified into family PL-7: a novel alginate lyase with a beta-sandwich fold. J Biol Chem 2004; 279:31863-72; PMID:15136569
- Park D, Jagtap S, Nair SK. Structure of a PL17 Family Alginate Lyase Demonstrates Functional Similarities among Exotype Depolymerases. J Biol Chem 2014; 289:8645-55; PMID:24478312
- Mikami B, Ban M, Suzuki S, Yoon HJ, Miyake O, Yamasaki M, Ogura K, Maruyama Y, Hashimoto W, Murata K. Induced-fit motion of a lid loop involved in catalysis in alginate lyase A1-III. Acta Crystallogr D 2012; 68:1207-16; PMID:22948922
- Zhu BW, Huang LS, Tan HD, Qin YQ, Du YG, Yin H. Characterization of a new endo-type polyM-specific alginate lyase from Pseudomonas sp. Biotechnol 2014; Lett); PMID:25257600; http://dx.doi.org/10.1007/s10529-014-1685-0
- Malissard M, Duez C, Guinand M, Vacheron MJ, Michel G, Marty N, Joris B, Thamm I, Ghuysen JM. Sequence of a gene encoding a (Poly Mana) alginate lyase active on pseudomonas-aeruginosa alginate. FEMS Microbiol Lett 1993; 110:101-6; PMID:8319887
- Sil'chenko AS, Kusaikin MI, Zakharenko AM, Zvyagintseva TN. Isolation from the marine mollusk Lambis sp and catalytic properties of an alginate lyase with rare substrate specificity. Chem Nat Compd+, 2013; 49:215-8
- Tondervik A, Klinkenberg G, Aarstad OA, Drablos F, Ertesvag H, Ellingsen TE, Skjak-Braek G, Valla S, Sletta H. Isolation of Mutant Alginate Lyases with Cleavage Specificity for Di-guluronic Acid Linkages. J Biol Chem 2010; 285:35284-92; PMID:20826807
- Yamasaki M, Ogura K, Hashimoto W, Mikami B, Murata K, A structural basis for depolymerization of alginate by polysaccharide lyase family-7. J Mol Biol 2005; 352:11-21; PMID:16081095
- Kim HS, Lee CG, Lee EY, Alginate Lyase: Structure, Property, and Application. Biotechnol Bioproc E 2011; 16:843-51.
- Deng SP, Ye J, Xu QQ, Zhang HZ, Structural and Functional Studies on Three Alginate Lyases from Vibrio alginolyticus, Protein Peptide Lett 2014; 21:179-87; PMID:24050202
- Brown BJ, Preston JF, Ingram LO, Cloning of Alginate Lyase Gene (Alxm) and Expression in Escherichia-Coli, Appl Environ Microbiol 1991; 57:1870-2; PMID:1872617
- Lee SI, Choi SH, Lee EY, Kim HS, Molecular cloning, purification, and characterization of a novel polyMG-specific alginate lyase responsible for alginate MG block degradation in Stenotrophomas maltophilia KJ-2. Appl Microbiol Biotechnol 2012; 95:1643-53; PMID:22805784; http://dx.doi.org/10.1007/s00253-012-4266-y
- Ostgaard K. Enzymatic microassay for the determination and characterization of alginates. Carbohyd Polym 1992; 19:51-9
- Malissard M, Chavagnat F, Duez C, Vacheron MJ, Guinand M, Michel G, Ghuysen JM, Overproduction and properties of the mannuronate alginate lyase AlxMB. FEMS Microbiol Lett 1995; 126:105-11; PMID:7705601
- Takeuchi T, Murata K, Kusakabe I. A method for depolymerization of alginate using the enzyme-system of flavobacterium-multivolum. J Jpn Soc Food Sci 1994; 41:505-11.
- Rodde RSH, Ostgaard K, Larsen BA, Mannuronan C-5 epimerase activity in protoplasts of laminaria-digitata. Hydrobiologia 1993; 261:577-81.
- Takeda H, Yoneyama F, Kawai S, Hashimoto W, Murata K. Bioethanol production from marine biomass alginate by metabolically engineered bacteria. Energ Environ Sci 2011; 4:2575-81.
- Enquist-Newman M, Faust AME, Bravo DD, Santos CNS, Raisner RM, Hanel A, Sarvabhowman P, Le C, Regitsky DD, Cooper SR, et al. Efficient ethanol production from brown macroalgae sugars by a synthetic yeast platform. Nature 2014; 505:239-43; PMID:24291791; http://dx.doi.org/10.1038/nature12771
- Wargacki AJ, Leonard E, Win MN, Regitsky DD, Santos CNS, Kim PB, Cooper SR, Raisner RM, Herman A, Sivitz AB, et al. An Engineered Microbial Platform for Direct Biofuel Production from Brown Macroalgae. Science 2012; 335:308-13; PMID:22267807