
ABSTRACT
Circular RNAs (circRNAs) feature prominently in regulating the progression of tumors, including papillary thyroid carcinoma (PTC). This work is designated to delve into the role of circ_0062389 in PTC. Generally, quantitative real-time polymerase chain reaction (qRT-PCR) was employed to detect circ_0062389, miR-1179 and high mobility group box 1 (HMGB1) mRNA expression levels. RNase R assay was used to verify the circular characteristics of circ_0062389. After circ_0062389 was knocked down in PTC cells, CCK-8 assay was adopted to determine cell viability. Wound healing assay was leveraged to probe cell migration. Besides, Western blot assay was executed to examine the expression levels of HMGB1 and epithelial-mesenchymal transformation (EMT)-related markers (E-cadherin and N-cadherin). Dual-luciferase reporter assay was performed to authenticate the targeting relationships between miR-1179 and circ_0062389, as well as miR-1179 and HMGB1. Here, this work proved that circ_0062389 was greatly up-regulated in PTC tissues and cell lines. The high expression of circ_0062389 was related to large tumor size and positive lymphatic metastasis. Knocking down circ_0062389 could inhibit the proliferation, migration and EMT process of PTC cells. Besides, miR-1179 was a downstream molecule of circ_0062389. Furthermore, miR-1179 inhibitors could partially reverse the above effect of knocking down circ_0062389 on PTC cells. It was also confirmed that HMGB1 was a direct target of miR-1179 and mediated the effects of circ_0062389 and miR-1179 in PTC. Altogether, circ_0062389 can adsorb miR-1179, and regulate HMGB1 expression, thus playing a role in PTC.
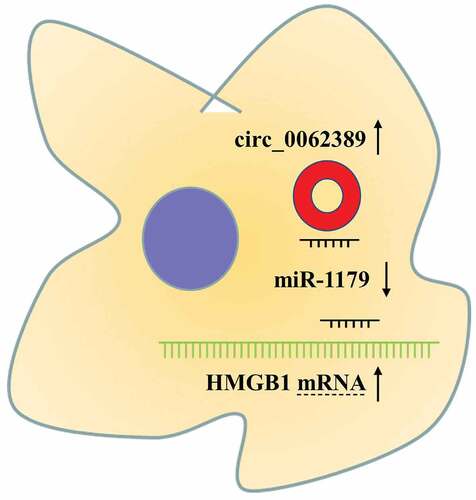
Graphical abstract
Introduction
Papillary thyroid cancer (PTC) is a common tumor, accounting for about 80%-85% of thyroid cancer cases [Citation1]. The 10-year survival rate of PTC patients exceeds 90% [Citation2]. However, some PTC patients still have a poor prognosis due to their older age, larger primary tumor, distant metastasis and lymph node metastasis [Citation3]. It is necessary to further improve the treatments of PTC, and it is of great significance to explore the pathogenesis of PTC for finding new targets for PTC treatment.
Circular RNAs (circRNAs) are non-coding RNAs with a covalently closed circular structure, without 5ʹ cap or 3ʹ poly A tail [Citation4]. CircRNA abnormal expression has been confirmed to be involved in many human diseases, especially tumors [Citation5]. Reportedly, many circRNAs are implicated in PTC progression, such as circRNA_102002, circ_LDLR and circPRMT56-[Citation6]. Circ_0062389, as a member of circRNAs, regulates CCNE1 expression by adsorbing miR-103a-3p to promote the progression of non-small cell lung cancer (NSCLC) [Citation7]. However, the function and molecular mechanism of circ_0062389 in the progression of PTC are still indeterminate.
MicroRNAs (miRNAs), non-coding RNAs with about 22 nucleotides, regulate gene expression via interacting with the 3ʹ untranslated region (3ʹUTR) of mRNA [Citation8]. Many miRNAs, as reported, are involved in PTC progression, such as miR-224-5p, miR-188-5p and miR-451a [Citation9–11]. In addition, miR-1179 can affect the progression of various tumors, such as oral cancer, gastric cancer and breast cancer [Citation12–14]. In PTC, miR-1179 can target HMGB1 or ABCA9 to inhibit PTC progressions [Citation15,Citation16]. However, the mechanism of miR-1179 dysregulation in PTC has not been completely elaborated.
High Mobility Group Box 1 (HMGB1) is involved in cancer cell proliferation, autophagy, metastasis and epithelial-mesenchymal transition (EMT) via regulating various signaling pathways [Citation17]. Reportedly, HMGB1 participates in the progression of many diseases, including PTC [Citation17]. However, the mechanism of HMGB1 dysregulation in PTC still awaits more verification.
This work aims to explore the expression characteristics and underlying mechanism of circ_0062389 in PTC. It was hypothesized that circ_0062389 was up-regulated in PTC tissues, and accelerate PTC cell growth, migration and EMT via modulating miR-1179/HMGB1 axis, and the experiments were performed to validate the scientific hypothesis mentioned above.
Materials and methods
Participants and tissue samples
Paired PTC tissues and adjacent tissues (n = 58) were available from Dalian Friendship Hospital, and all samples, stored at −80°C immediately after removal to avoid RNA loss, were confirmed by pathologists. None of the patients had received radiotherapy, chemotherapy or other treatment before the surgery. This study, with signed written informed consent from the patients, was approved by the Ethics Committee of Dalian Friendship Hospital.
Cell culture and transfection
Human PTC cell lines (K-1, SW1736, TPC-1, SW579) and human thyroid follicular epithelial cells Nthy-ori 3–1 were available from Cell Bank of Chinese Academy of Sciences (Shanghai, China). Notably, all cells were accordingly cultured in Roswell Park Memorial Institute (RPMI)-1640 medium with 10% fetal bovine serum (FBS), 100 IU/mL penicillin and 100 μg/mL streptomycin. The above materials or reagents were from Gibco (Carlsbad, CA, USA).
Small interfering RNA (siRNA) oligonucleotides targeting circ_0062389 (si-circ_0062389#1: 5ʹ-TGCAGACCTCCCGACCTCTTT-3ʹ, si-circ_0062389#2: 5ʹ-ATCTGCTGTCTGCAGACCTCC-3ʹ and si-circ_0062389#3: 5ʹ-GATCTGCTGTCTGCAGACCTC-3ʹ), miR-1179 mimics (5ʹ-AAGCAUUCUUUCAUUGGUUGG-3ʹ) and miR-1179 inhibitors (miR-1179 in, 5ʹ- CCAACCAAUGAAAGAAUGCUU-3ʹ) and their corresponding control (miR-NC, 5ʹ-UCACAACCUCCUAGAAAGAGUAGA-3ʹ) were synthesized by Genomeditech (Shanghai, China). The above vector plasmids or oligonucleotides were respectively transfected into PTC cell lines by lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) when the cells reached 80% confluence.
Quantitative real-time polymerase chain reaction (qRT-PCR) and RNase R digestion assay
20 mg/ml RNase R (Genesee, Guangzhou, China) was incubated with 10 μg RNA at 37°C for 20 min, and a control group without RNase R treatment was set. Total RNA was extracted from tissues and cell lines by TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and reverse transcription was performed with PrimeScript™ RT reagent Kit (Takara, Dalian, China). Following that, a SYBR Green Premix Ex TaqTM kit (Takara, Kusatsu, Japan) was adopted to perform qRT-PCR, with relative RNA expressions calculated by 2−ΔΔCt formula, and glycerol aldehyde-3-phosphate dehydrogenase (GAPDH) and U6 were regarded as internal references. Primer sequences are detailed in .
Table 1. Primer sequences
Cell counting kit-8 (CCK-8) assay
Cell proliferation was detected by CCK-8 assay. PTC cells were inoculated on a 96-well plate at a density of 2 × 103 cells/well and cultured, and 10 µL of CCK-8 solution (Solarbio, Beijing, China) was added into each well at 0 h, 24th h, 48th h, and 72th h. And the cells were accordingly incubated for another 4 h. Ultimately, the absorbance at 450 nm was measured and recorded by a microplate reader (Thermo-Fisher Scientific, Waltham, MA, USA).
Wound healing assay
Transfected cells were inoculated on a 6-well plate. When the cells reached 90% confluence, a cell-free region was created by scraping the monolayer cells with a 200 μL micropipette. Cells were cultured in serum-free medium, and the wound closure was measured at 0 h and 24th h. Wound width (%) = (0 h wound closure-24th h wound closure)/0 h wound closure ×100%。
Bioinformatics analysis
StarBase database (starbase.sysu.edu.cn) was used to predicted the binding sites between circRNA and miRNA, as well as miRNA and mRNA.
Dual-luciferase reporter assay
Wild-type and mutant reporter plasmids of circ_0062389 or HMGB1 3ʹUTR containing the putative binding sequence with miR-1179 were designed and constructed by Promega (Madison, WI, USA). Above luciferase reporters were respectively co-transfected with miR-1179 mimics or miR-NC into PTC cells, and the dual-luciferase reporter assay system (Promega, Madison, Wis., USA) was utilized to detect the luciferase activity of the cells in each group after 48 h.
Western blot assay
Total protein was respectively extracted from cells in different groups by radio-immunoassay assay (RIPA) lysis buffer (Beyotime, Shanghai, China), with the concentration of protein quantified by a bicinchoninic acid (BCA) detection kit (Beyotime, Shanghai, China). 30 μg protein samples in each group were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to polyvinyl fluoride (PVDF) membranes (Millipore, Billerica, MA, USA), which was then blocked with 5% skimmed milk. Then the PVDF membrane was incubated with primary antibodies anti‐HMGB1 (1:1000, ab18256, Abcam Inc., Cambridge, UK), anti‐E-cadherin (1:1000, ab40772, Abcam Inc., Cambridge, UK), anti‐N-cadherin (1:1000, ab98952, Abcam Inc., Cambridge, UK) or anti-GAPDH (1:1000, ab9485, Abcam Inc., Cambridge, UK) overnight at 4°C, and then incubated with the horseradish peroxidase-conjugated secondary antibody (1:2000, ab150077, Abcam Inc., Cambridge, UK) at ambient temperature for 1 h, and finally the ECL luminescence reagent (ThermoFisher Scientific, Waltham, MA, USA) was employed to detect the protein bands.
Statistical analysis
All experiments were repeated for three times before collecting data, with results expressed as ‘mean ± standard deviation’. SPSS17.0 software (SPSS Inc., Chicago, IL, USA) was used for statistical analysis and GraphPad Prism V5.0 (GraphPad Software, Inc., La Jolla, CA, USA) was used for drawing the plots. The PTC patients were grouped into circ_0062389 high expression group and circ_0062389 low group with the median expression of circ_0062389 as the threshold. The correlation between circ_0062389 expression and the clinicopathological characteristics of the patients was analyzed by Chi-square test. Student’s t test was adopted for difference analysis between two groups, and one-way analysis of variance was adopted for difference analysis among three or more groups. Pearson correlation was adopted for correlation analysis. Statistically, *P < 0.05, **P < 0.01, and ***P < 0.001 were meaningful.
Results
The aim of this work was to explore the expression pattern and function of circ_0062389 in PTC, and it was demonstrated that that circ_0062389 expression was raised in PTC tissues and cell lines. We also reported that, circ_0062389 could promote PTC cell viability, migration and invasion by sponging miR-1179 and upregulating the expression of HMGB1.
The expression and characteristics of circ_0062389 in PTC
To identify PTC-related circRNAs, a circRNA microarray (GSE93522) was analyzed to identify the differentially expressed circRNAs in 6 pairs of PTC tissues and adjacent tissues. According to the criteria (fold-change > 2.0 and P value < 0.05), heat maps and volcano plots were drawn, and the results showed that circ_103164 (also known as circ_0062389) expression was dramatically raised in PTC tissues (&B). Then, qRT-PCR also showed that in 58 PTC patients, circ_0062389 expression in PTC tissues was markedly higher than that in adjacent tissues () and the high expression of circ_0062389 was related to large tumor size and positive lymph node metastasis (). Circ_0062389 expression level in thyroid cancer cell lines (K-1, SW1736, TPC-1, SW579) was also significantly higher compared with that in Nthy-ori 3–1 cell line (). Considering the highest expression of circ_0062389 in SW1736 and SW579, these two cell lines were selected for the follow-up functional experiments.
Table 2. The relationship between circ_0062389 expression and clinicopathological features
Figure 1. Circ_0062389 is significantly up-regulated in PTC tissues and cell lines (a)&(b) Volcanic and heat maps showed that multiple circRNAs, including Circ_0062389, were differentially expressed in PTC tissues and adjacent tissues in GSE93522; (c) qRT-PCR was used to detect the expression of circ_0062389 in PTC tissues and adjacent tissues; (d) qRT-PCR was performed to evaluate the expression of circ_0062389 in PTC cell lines and normal cell line; (e)&(f) After RNase R treatment, qRT-PCR was used to detect the expression of circ_0062389 and PI4KA mRNA in total RNA. ***P < 0.001
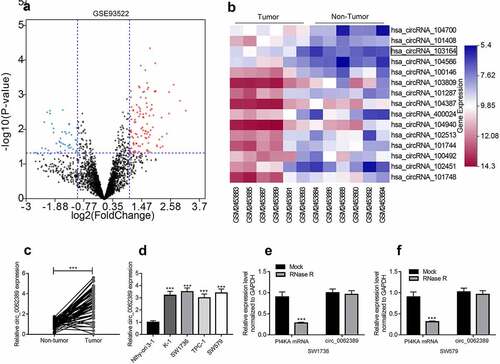
In addition, RNase R was used to treat total RNA and verify the circular character of circ_0062389. The results showed that circ_0062389 was resistant to RNase R digestion, whereas the linear PI4KA mRNA was degraded by RNase R (&F). Therefore, it was revealed that circ_0062389 was stable in PTC and may be related to the progression of PTC.
Circ_0062389 can promote the multiplication, migration and EMT of PTC cells
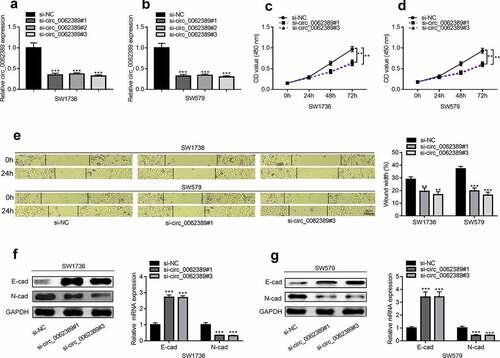
Considering that circ_0062389 was elevated in PTC tissues and cell lines, loss-of-function experiments were adopted to explore the function of circ_0062389 in PTC. Three circ_0062389 siRNAs (si-circ_0062389#1, si-circ_0062389#2 and si-circ_0062389#3) were transfected into SW1736 and SW579 cell lines, and the results showed that circ_0062389 siRNAs significantly decreased circ_0062389 expression in SW1736 and SW579 cell lines, and si-circ_0062389#1 and si-circ_0062389#3 had higher transfection efficiency, which were used for subsequent experiments (&B). CCK-8 assay uncovered that knocking down circ_0062389 in SW1736 and SW579 cell lines could significantly inhibit cell proliferation (&D). As against si-NC group, the ability of cell migration in si-circ_0062389#1 and si-circ_0062389#3 groups was significantly reduced (). Also, qRT-PCR and Western blot assays proved that circ_0062389 depletion significantly increased E-cadherin protein expression and decreased N-cadherin protein expression (&G). These data confirmed that knocking down circ_0062389 could inhibit the growth, migration and EMT of PTC cells.
Figure 2. Circ_0062389 can promote the proliferation, migration and EMT process of PTC cells (a)&(b) qRT-PCR was used to detect the expression of circ_0062389 after transfection of three circ_0062389 siRNA into PTC cells; (c)&(d) CCK-8 assay was used to evaluate the proliferation of PTC cells transfected with circ_0062389 siRNA; (e) Wound healing assay was adopted to measure cell migration in PTC cells after transfection of circ_0062389 siRNA; (f)&(g) qRT-PCR and Western blot assay were used to detect the expression of E-cadherin and N-cadherin at mRNA and protein levels in PTC cells transfected with circ_0062389 siRNA. **P < 0.01, and ***P < 0.001
Circ_0062389 is the molecular sponge of miR-1179 in PTC cells
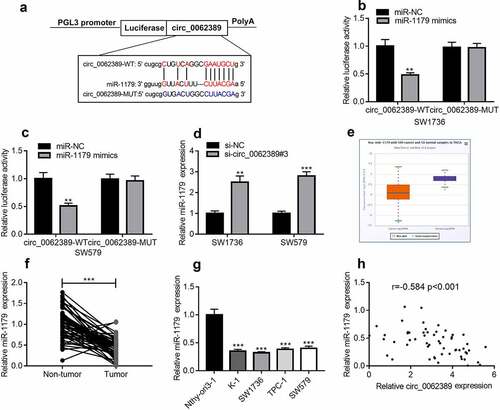
StarBase database showed that circ_0062389 contained potential complementary binding sites with miR-1179 (). Dual-luciferase reporter gene assay highlighted that miR-1179 mimics could decrease the luciferase activity of wild type circ_0062389 reporter plasmid, but did not significantly change that of mutant circ_0062389 reporter plasmid (&C). In addition, circ_0062389#3 with the highest knock-down efficiency was transfected into SW1736 and SW579 cell lines, and it was found that circ_0062389 inhibition significantly increased miR-1179 expression (). StarBase database confirmed that miR-1179 was dramatically declined in PTC tumor tissues relative to normal tissues (). qRT-PCR also found that miR-1179 expression in PTC tissues was greatly lower than that in adjacent tissues (), and its expression in PTC cell lines was also markedly lower than that in normal cell line (). Pearson correlation analysis confirmed that miR-1179 expression level was negatively correlated with circ_0062389 expression level in PTC tissues (). Thus, our data indicated that circ_0062389 was the molecular sponge of miR-1179 in PTC.
Figure 3. MiR-1179 is a target of circ_0062389 (a)StarBase database predicted that there was a binding site between miR-1179 and circ_0062389; (b)&(c) Dual-luciferase reporter gene assay confirmed that miR-1179 could negatively regulate the luciferase activity of circ_0062389-WT, rather than that of circ_0062389-MUT; (d) qRT-PCR was used to detect miR-1179 expression after PTC cells were transfected with circ_0062389 siRNA; (e) StarBase database was adopted to analyze miR-1179 expression in PTC tissue and normal tissue; (f) qRT-PCR was used to detect the expression of miR-1179 in PTC tissues and adjacent tissues; (g) qRT-PCR was used to detect miR-1179 expression in PTC cell lines and normal cell line; (h) The correlation analysis of circ_0062389 and miR-1179 expression in PTC tissues. **P < 0.01; and ***P < 0.001
Circ_0062389 participates in PTC through miR-1179
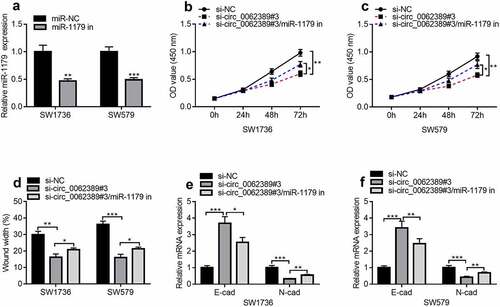
Next, whether circ_0062389 could play a role through miR-1179 was further explored. MiR-1179 inhibitors were transfected into SW1736 and SW579 cell lines and the results uncovered that miR-1179 inhibitors could dramatically reduce the expression of miR-1179 (). Then circ_0062389 siRNA and miR-1179 inhibitors were co-transfected into SW1736 and SW579 cell lines. CCK-8 assay showed that miR-1179 inhibitors could partially reverse the effect of knocking down circ_0062389 on cell proliferation (&C). Wound healing assay uncovered that co-transfection of circ_0062389 siRNA and miR-1179 inhibitors markedly increased cell migration compared with si-circ_0062389#3 group (). qRT-PCR confirmed that miR-1179 inhibitors could partially reverse the increase of E-cadherin mRNA expression and the decrease of N-cadherin mRNA expression caused by knocking down circ_0062389 (&F). The aforementioned findings revealed that circ_0062389 could regulate PTC cell growth, migration and EMT via modulating miR-1179.
Figure 4. Circ_0062389 exerts promoting effects in PTC through modulating miR-1179. (a) qRT-PCR was used to detect the expression of miR-1179 after transfection of miR-1179 inhibitors into PTC cells; (b)&(c) CCK-8 assay was used to detect the proliferation of PTC cells after co-transfection of circ_0062389 siRNA and miR-1179 inhibitors; (d) Wound healing assay was used to detect PTC cell migration after co-transfection of circ_0062389 siRNA and miR-1179 inhibitors into PTC cells; (e)&(f) qRT-PCR was used to detect the mRNA expression of E-cadherin and N-cadherin after co-transfection of circ_0062389 siRNA and miR-1179 inhibitors into PTC cells. *P < 0.05, **P < 0.01, and ***P < 0.001
Circ_0062389 regulates HMGB1 expression through miR-1179
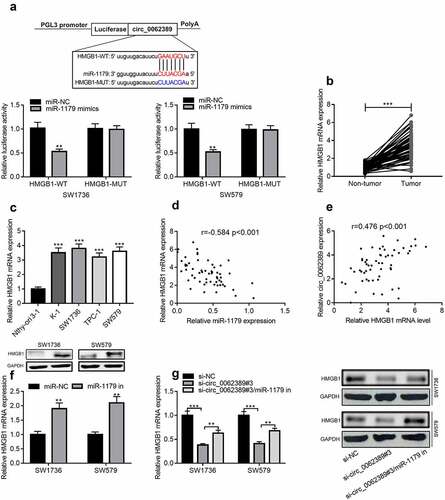
StarBase database was used to predict the downstream target of miR-1179, and HMGB1 3ʹUTR contained a binding site complementary to miR-1179. Dual-luciferase reporter gene assay confirmed that miR-1179 mimics could greatly reduce the luciferase activity of wild-type HMGB1 reporter plasmid, but had no obvious impact on that of mutant HMGB1 reporter plasmid (). qRT-PCR confirmed that HMGB1 mRNA was markedly raised in PTC tissues and cell lines (&C). Pearson correlation analysis highlighted that HMGB1 mRNA expression level was negatively correlated with miR-1179 expression level, and positively correlated with circ_0062389 expression level (&E). Additionally, miR-1179 inhibitors could significantly increase HMGB1 mRNA and protein expression levels (). In addition, miR-1179 inhibitors could partially reverse the inhibitory effect of knocking down circ_0062389 on HMGB1 mRNA and protein expression levels (). Therefore, the above experiments confirmed that circ_0062389 regulated HMGB1 expression through repressing miR-1179 in PTC cells.
Figure 5. Circ_0062389 can adsorb miR-1179 to regulate HMGB1 expression (a) StarBase database predicted that circ_0062389 and miR-1179 had a complementary binding site, and dual-luciferase reporter gene asasy confirmed that circ_0062389 could bind with miR-1179 directly; (b) qRT-PCR was used to evaluate the expression of HMGB1 mRNA in PTC tissues and adjacent tissues; (c) qRT-PCR was used to detect HMGB1 mRNA expression in PTC cell lines and human normal thyroid follicular epithelial cell line; (d)&(e) The correlation analysis of miR-1179 and HMGB1 mRNA, HMGB1 mRNA and circ_0062389 in PTC tumor tissues; (f)&(g) qRT-PCR and Western blot assay were used to detect the expression of HMGB1 mRNA and protein in PTC cells after transfection of miR-1179 inhibitors and co-transfection of circ_0062389 siRNA and miR-1179 inhibitors. **P < 0.01, and***P < 0.001
Discussion
CircRNAs, as reported, are pivotal in different biological processes, especially in tumorigenesis, progression and metastasis. CircRNAs may be promising biomarkers and targets in tumor diagnosis and treatment [Citation18]. Recently, as reported, circRNAs exert important regulatory parts in PTC [Citation6,Citation7,Citation19,Citation20]. For example, circRNA_102002 promotes PTC metastasis by regulating miR-488-3p/HAS2 axis [Citation19]; circ_LDLR promotes PTC progression via modulating miR-195-5p/LIPH axis [Citation20]; circPRMT5 promotes PTC cell multiplication and invasion via regulating miR-30 c/E2F3 axis [Citation6]. Circ_0062389 expression level is significantly raised in NSCLC and can promote cell proliferation and accelerate cell cycle progression [Citation7]. Here, the present study showed that circ_0062389 was remarkably up-regulated in PTC tumor tissues and cell lines, being resistant to RNase R digestion. The high expression of circ_0062389 was related to large tumor size and positive lymph node metastases. In addition, knocking down circ_0062389 could significantly inhibit the viability (**P < 0.01), migration (**P < 0.01) and EMT (***P < 0.001) of PTC cells. Taken together, these results confirm that circ_0062389 is structurally stable and is a cancer-promoter in the PTC progression.
CircRNAs can adsorb miRNAs and play a role as a competitive endogenous RNA (ceRNA) in PTC [Citation21]. In the present study, StarBase database and dual-luciferase reporter gene assay proved that miR-1179 could combine with circ_0062389. MiR-1179 is down-regulated in many kinds of tumors, and blocks cancer progression. For example, in cervical cancer, circ_0084927 can adsorb miR-1179 to regulate CDK2 expression, promote cell proliferation and inhibit apoptosis [Citation22]. MiR-1179 expression is markedly declined in oral cancer, and it can inhibit cell proliferation, enhance vincristine sensitivity and induce apoptosis by regulating MEK/ERK and PI3K/AKT signaling pathways [Citation12]. In gastric cancer, miR-1179 restrains cell proliferation by targeting HMGB115. Here, it was demonstrated that miR-1179 was remarkably decreased in PTC tumor tissues and cell lines, and was negatively correlated with circ_0062389 expression; miR-1179 inhibitors could partially reverse the suppressing effect of knocking down circ_0062389 on the malignant phenotypes of PTC cells. Therefore, it is concluded that circ_0062389 adsorbs miR-1179 in PTC, thus participating in PTC progression.
StarBase database and dual-luciferase reporter assay showed that HMGB1 was the target of miR-1179. HMGB1, a nuclear protein that exists in almost all eukaryotic cells, consists of 215 amino acid residues, and features prominently in maintaining the stability of genome structure [Citation23–25]. Importantly, HMGB1 is a crucial regulator in cancer biology. Reportedly, HMGB1 can promote the invasion of NSCLC cells by activating PI3K/Akt and NF-κB pathways to produce MMP-9[Citation24]; in metastatic pancreatic ductal adenocarcinoma, HMGB1 promotes EMT and invasion of cancer cells by acting on RAGE/NF-κB axis. In this work, it was observed that HMGB1 was dramatically raised in PTC tumor tissues and cell lines, and HMGB1 mRNA expression was negatively correlated with miR-1179 expression and positively correlated with circ_0062389 expression in PTC tumor tissues. In PTC cells, miR-1179 could negatively regulate HMGB1 expression, and miR-1179 inhibitors can partially reverse the inhibitory effect of knocking down circ_0062389 on HMGB1 expression. Therefore, circ_0062389 regulates HMGB1 expression via adsorbing miR-1179, to exert its regulatory function on the malignant phenotypes of PTC cells.
Conclusion
Collectively, circ_0062389 expression level is significantly raised in PTC tissues and cell lines, and circ_0062389 participates in PTC progression as a tumor-promoter. Mechanistically, circ_0062389 can adsorb miR-1179 and up-regulate HMGB1. This study may provide a theoretical basis for further exploring biomarkers and therapy targets for PTC diagnosis and treatment.
Highlights
· Circ_0062389 was up-regulated in PTC tissues and cell lines.
· Circ_0062389 could promote the proliferation, migration and epithelial-mesenchymal transition (EMT) of PTC cells.
· Circ_0062389 regulated HMGB1 expression through miR-1179.
Ethics statement
Our study was approved by the Ethics Review Board of Dalian Friendship Hospital.
Supplemental Material
Download ()Disclosure statement
The authors declare that they have no competing interests.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Supplementary material
supplemental data for this article can be accessed here
Additional information
Funding
Related Research Data
References
- Cabanillas ME, McFadden DG, Durante C. Thyroid cancer. Lancet. 2016;388(10061):2783–2795.
- Jiang C, Cheng T, Zheng X, et al. Clinical behaviors of rare variants of papillary thyroid carcinoma are associated with survival: a population-level analysis. Cancer Manag Res. 2018;10:465–472.
- Berdelou A, Lamartina L, Klain M, et al. Treatment of refractory thyroid cancer. Endocr Relat Cancer. 2018;25(4):R209–R223.
- Meng S, Zhou H, Feng Z, et al. CircRNA: functions and properties of a novel potential biomarker for cancer. Mol Cancer. 2017;16(1):94. .
- Shang Q, Yang Z, Jia R, et al. The novel roles of circRNAs in human cancer. Mol Cancer. 2019;18(1):6.
- Xue C, Cheng Y, Wu J, et al. Circular RNA CircPRMT5 accelerates proliferation and invasion of papillary thyroid cancer through regulation of miR-30c/E2F3 axis. Cancer Manag Res. 2020;12:3285–3291.
- She Y, Han Y, Zhou G, et al. hsa_circ_0062389 promotes the progression of non-small cell lung cancer by sponging miR-103a-3p to mediate CCNE1 expression. Cancer Genet. 2020;241:12–19.
- He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5(7):522–531.
- Zang CS, Huang HT, Qiu J, et al. MiR-224-5p targets EGR2 to promote the development of papillary thyroid carcinoma. Eur Rev Med Pharmacol Sci. 2020;24(9):4890–4900.
- Zhou P, Irving A, Wu H, et al. Validation of MicroRNA-188-5p inhibition power on tumor cell proliferation in papillary thyroid carcinoma. Cell Transplant. 2020;29:963689720918300.
- Wang Q, Shang J, Zhang Y, et al. MiR-451a restrains the growth and metastatic phenotypes of papillary thyroid carcinoma cells via inhibiting ZEB1. Biomed Pharmacother. 2020;127:109901.
- Gao Y, Xu H, Pu T. MicroRNA-1179 suppresses the proliferation and enhances vincristine sensitivity of oral cancer cells via induction of apoptosis and modulation of MEK/ERK and PI3K/AKT signalling pathways. AMB Express. 2020;10(1):149.
- Li Y, Qin C. MiR-1179 inhibits the proliferation of gastric cancer cells by targeting HMGB1. Hum Cell. 2019;32(3):352–359.
- Li WJ, Xie XX, Bai J, et al. Increased expression of miR-1179 inhibits breast cancer cell metastasis by modulating Notch signaling pathway and correlates with favorable prognosis. Eur Rev Med Pharmacol Sci. 2018;22(23):8374–8382.
- Yang Y, Ding L, Li Y, et al. Hsa_circ_0039411 promotes tumorigenesis and progression of papillary thyroid cancer by miR-1179/ABCA9 and miR-1205/MTA1 signaling pathways. J Cell Physiol. 2020;235(2):1321–1329.
- Ye M, Hou H, Shen M, et al. RNA circFOXM1 plays a role in papillary thyroid carcinoma by sponging miR-1179 and regulating HMGB1 expression. Mol Ther Nucleic Acids. 2020;19:741–750.
- Wang S, Zhang Y. HMGB1 in inflammation and cancer. J Hematol Oncol. 2020;13(1):116.
- Zhang HD, Jiang LH, Sun DW, et al. CircRNA: a novel type of biomarker for cancer. Breast Cancer. 2018;25(1):1–7.
- Zhang W, Liu T, Li T, et al. Hsa_circRNA_102002 facilitates metastasis of papillary thyroid cancer through regulating miR-488-3p/HAS2 axis. Cancer Gene Ther. 2020. DOI:10.1038/s41417-020-00218-z
- Gui X, Li Y, Zhang X, et al. Circ_LDLR promoted the development of papillary thyroid carcinoma via regulating miR-195-5p/LIPH axis. Cancer Cell Int. 2020;20(1):241.
- Chan JJ, Noncoding TY. RNA:RNA regulatory networks in cancer. Int J Mol Sci. 2018;19(5):1310.
- Qu X, Zhu L, Song L, et al. circ_0084927 promotes cervical carcinogenesis by sponging miR-1179 that suppresses CDK2, a cell cycle-related gene. Cancer Cell Int. 2020;20(1):333.
- He SJ, Cheng J, Feng X, et al. The dual role and therapeutic potential of high-mobility group box 1 in cancer. Oncotarget. 2017;8(38):64534–64550.
- Liu PL, Tsai JR, Hwang JJ, et al. High-mobility group box 1-mediated matrix metalloproteinase-9 expression in non-small cell lung cancer contributes to tumor cell invasiveness. Am J Respir Cell Mol Biol. 2010;43(5):530–538. .
- Cebrian MJ, Bauden M, Andersson R, et al. Paradoxical role of HMGB1 in pancreatic cancer: tumor suppressor or tumor promoter? Anticancer Res. 2016;36(9):4381–4389.