
ABSTRACT
This paper provides a conceptual framework for analyzing the dynamics of tree cover transformations in the cattle pastures across the Neotropics. It proposes that tree cover variability across cattle pastures can be envisioned as a set of ‘syndromes of production,’ among which transformations may be linear, non-linear, or multivalued. Our framework is informed by a review of the literature from which we define the components that make up tree cover and the socioecological drivers of tree cover in cattle pastures. We propose that the drivers of tree cover are the continuum between two economies: the peasant and the capitalist.
Introduction
Conversion of forest into pastures in the Neotropics has been of great concern for conservation since the large cattle ranching expansion of the 1960s (Grandia Citation2009; Hecht Citation1985; Zahawi, Duran, and Kormann Citation2015). Less concern has materialized with respect to the nature of those pastures, especially regarding the incorporation of trees into the production system. It is evident from casual observation that Neotropical farms with cattle vary dramatically in tree densities (Harvey et al. Citation2011), yet efforts to document and understand this variation have not been common. It wasn’t until 2009 that the World Agroforestry Center published the first global scale report on agroforestry land, and only in 2016 did we receive a full accounting of the significant carbon biomass contained therein (Zomer et al. Citation2016). Nevertheless, silvo-pastoral techniques with an agroecological vision have long been promoted as a sustainable alternative to the treeless cattle pastures so evident across the Neotropics (Broom, Galindo, and Murgueitio Citation2013; Murgueitio et al. Citation2011; Pagiola et al. Citation2007; Payne Citation1985). Although such tree management techniques are commonly encountered among small-scale producers throughout the Neotropics, and government and non-profits promote agroforestry, there is scant evidence of large-scale adoption (Dagang and Nair Citation2003; Pattanayak 2003; Mercer Citation2004; Calle, Montagnini, and Zuluaga Citation2009; Calle et al. Citation2013; Lerner et al. Citation2017).
Observed variability in silvo-pastoral systems () can be envisioned as a series of ‘syndromes of production’ (Andow and Hidaka Citation1989; Vandermeer Citation1997). A syndrome of production (see ) is a set of management practices that function and adapt in conjunction (Andow and Hidaka Citation1989). The silvo-pastoral syndrome is characterized by the tree densities, which work in conjunction with specific grazing management techniques and cultural repertoires that are tied to the values that are given to trees. A different set of management techniques and cultural repertoires characterize the treeless pasture syndrome. As a starting point, we simplify the production syndrome as the observed tree density (high and low), which is easy to conceptualize and measure (). We propose that the transformations from a treeless pasture to a silvo-pastoral syndrome may be studied through a dynamics framework (), as commonly applied in other ecosystems (see, e.g., Cale and Allen-Diaz Citation2013; Scheffer Citation2009; Staver, Archibald, and Levin Citation2011). We also propose that moving from a treeless cattle pasture to a silvo-pastoral system entails complex interactions among socioecological variables, ultimately leading to non-linearities and multivalued dynamics (Vandermeer and Perfecto Citation2012), characterized by multiple values of the dependent variable for one value of the independent variable. This dynamics framework may provide unique insights into how silvo-pastoral systems emerge and change.
Table 1. Concepts of the dynamics framework.
Figure 1. Illustrates the syndromes of production, formulated as a linear relationship between cattle density (or other disturbance) as the driving variable and tree density as the response variable. Adapted from Moguel and Toledo (Citation1999) depiction of five coffeegrowing systems.
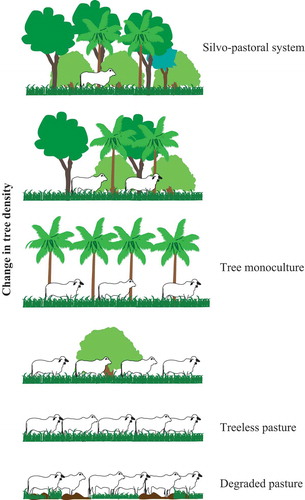
Figure 2. Dynamics between disturbance and tree density (a) Quasi- linear dynamics in which a large change in the disturbance results in a large change in tree density. (b) A strong but monotonic non-linear dynamic in which a small change in the disturbance leads to a large change in the tree density. (c) A multivalued pattern in which different equilibria may result from the same disturbance value.
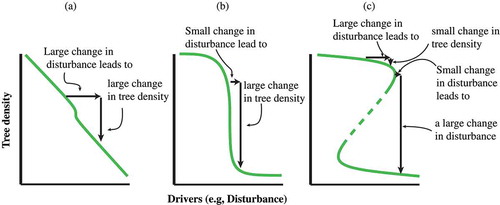
Models that propose linear dynamics between population and land use are widely used to explain tree density change across rural landscapes in the Neotropics. One example is Forest Transition Theory (FTT), which proposes that rural to urban population transitions will result in agricultural abandonment and forest recovery (Mather and Needle Citation1998; Rudel Citation1998). FTT assumes that forest cover can be easily reversed through agricultural abandonment and depopulation (Perz Citation2007), but it’s not difficult to imagine that many (perhaps most?) historical agricultural transformations are not easily reversible (Lockwood and Lockwood Citation1993). Hysteresis occurs when the dependent (or syndrome) variable responds slowly (if at all) to change in the independent (or socioecological driver) variables. Once a threshold is reached a small change in the driver condition will lead to a large change in the syndrome. When the response variable does change, generally the change is rapid (). For example, in San Andres Lagunas, in Oaxaca, Mexico, emigration led to transforming sustainable irrigation systems into slash-and-burn agriculture (García-Barrios and García-Barrios 1990). Emigration resulted in the breakdown of centralized institutions that were the core of these terrace/irrigation systems. Complex ecological and economic factors would have to be re-organized in order for the terrace/irrigation systems to return. Simply reversing the population decline will not automatically lead to recovery.
Figure 3. Depicts the proposed relationship between cattle density (a particular ‘disturbance’) and tree density as a multivalued dynamic. The rectangle indicates the zone of hysteresis. In the zone of hysteresis multiple equilibriums (or values for tree density) are possible: 1) stable equilibrium at high tree density, 2) stable equilibrium at low tree density, and 3) unstable equilibrium at intermediate tree densities. When critical tree densities are reached, the system rapidly transitions into a new stable tree density state.
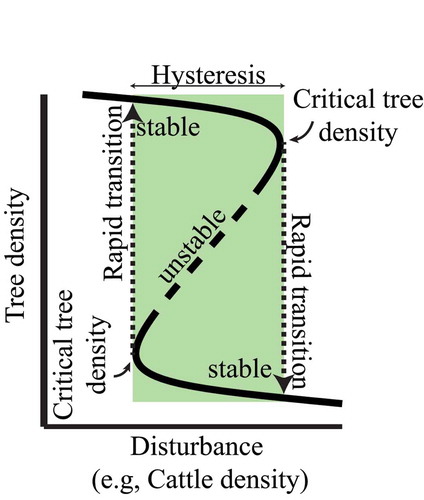
The present review places silvo pastoralism in the Neotropics within this particular framing. It seems obvious that the quantitative and qualitative features of the trees that ranchers and farmers conserve in pastures result from complex socioecological drivers that govern the farm. Frequently, there is a tacit assumption that those socioecological drivers, while complicated, can be mapped into a one-to-one relationship with trees (); thus a graph of tree quantity (or some other measure of tree prevalence) as a function of socioeconomic force will appear as a simple monotonic function ()). Monotonic functions are presented in ,), where there is one value on the dependent variable (Y axis) for every value on the independent variable (X axis). This relationship, seemingly evident, may be erroneous given the complexity of factors involved. We propose that the dynamics of this relationship are more likely to be multivalued ()), including the important condition of hysteresis ().
In this initial formulation, the independent variable is some measure of socioecological ‘disturbance’ (). Disturbance in our usage refers to any socioecological process that reduces trees directly or restricts future establishment of trees. We may assume that the relationship between tree density and disturbance is quasi-linear, which is to say, a small change in disturbance frequency will lead to a small change in tree density, while a large change in disturbance frequency will lead to a larger change ()). Yet, such a quasilinear assumption does not necessarily accord well with field observations, and frequently small changes in management (disturbance) can lead to much larger changes in tree density (,)). Another potential pattern, the multivalued pattern mentioned above that involves hysteresis ()), seems to pass with little notice.
Thus we propose that the relationship between disturbance frequency and tree density falls into one of three qualitatively distinct models: quasilinear ()), strongly nonlinear ()), or multivalued ()). In the third case, there are two values for tree density (large or small) for any disturbance value within the hysteretic zone (), a fact that carries with it important lessons for ecosystem management. Hysteretic systems resist change until the driving force reaches a threshold, or tipping point resulting in dramatic and sudden shifts in the response variable ().
The agroforestry literature emphasizes adoption of silvo-pastoral systems that involve intentional tree planting as part of the initial agronomic designs (Broom, Galindo, and Murgueitio Citation2013; Calle, Montagnini, and Zuluaga Citation2009; Garen et al. Citation2011). However, with the exception of live fences in some areas, most trees in Neotropical pastures exist either because they survived initial pasture establishment or as a result of natural regeneration that ranchers and small farmers foster or tolerate as they make farm management decisions (Harvey et al. Citation2011; Sanfiorenzo-Barnhard et al. Citation2009; Somarriba et al. Citation2012). Ranchers and farmers may perceive trees as integral to pasture management or as negligible. These perceptions will depend on a complex set of factors that shape their farming goals. We propose that the drivers of tree densities on pastures can be understood by employing Chayanov’s Theory of Peasant Economy (Chayanov Citation1986) and its recent resurrection by van der Ploeg (Citation2013) in the Art of Peasant Farming. Chayanov (Citation1986) proposes that farmers make farming decisions by balancing various aspects of their livelihood. The most basic balance that farmers seek is that between the household consumption (or mouths to feed) and the amount of labor put into farming. A related balance is the utility of what they produce versus the drudgery involved (van der Ploeg Citation2013). We apply the ‘balances’ framework to understand the drivers of tree cover on the farm. Ultimately, we combine two epistemologies, the dynamics framework so commonly employed in the natural sciences, and the peasant balances framework drawn from Chayanov (Citation1986).
The main objective of this review is to use the dynamics framework summarized in to explore how linear, non-linear, and multivalued dynamics can be applied to understand the extant tree cover densities and their potential for change across silvo-pastoral landscapes. Our focus is on the possibility that multivalued dynamics may be operative, thus identifying high- and low-tree-density farms as syndromes of production. Our first specific objective is to describe the dependent and independent variables that give rise to tree-density syndromes. Our second specific objective is to discuss how the balances described by Chayanov (Citation1986) and van der Ploeg (Citation2013) may be envisioned as drivers of tree cover across the cattle farms and ranches of the Neotropics. We begin with discussions of the general conceptual framework, followed by a discussion of the syndromes that seem to emerge in the examples of cattle ranching across Central America and Brazil. We also discuss the ecological and social drivers (or dependent variables) of the tree syndromes. Finally, we discuss the implications of this framework for future research on silvo-pastoral systems.
The multivalued framework
The multivalued framework we employ is motivated in part by the recent discoveries that forest and savanna may be alternative states along a range of precipitation in various continents (Hirota et al. Citation2011; Staver, Archibald, and Levin Citation2011). The gradient from a high-density silvo-pastoral system to a treeless pasture can be conceptualized by comparison to the similar natural systems of tropical vegetation (Hirota et al. Citation2011; Staver, Archibald, and Levin Citation2011). Under common tropical conditions, relatively low precipitation will inevitably produce a savanna, and high precipitation will inevitably produce a forest. At intermediate levels of precipitation, historical evidence suggests that two alternative tree density values (savanna state and forest state) exist for any precipitation value. Over that range of annual precipitation, beyond a critical density, trees create locally more humid conditions, thus resisting fire and rapidly transitioning from savanna to forest. Yet, at that same level of annual precipitation, if the tree density is below the critical threshold, the microclimate becomes drier and the system will move rapidly to the periodic fire and savanna state. The zone in which such critical tree densities mark the dividing line between forest and savannah is a hysteretic zone. There is also strong evidence for a third state, the treeless state (Hirota et al. Citation2011). Trees can contribute to the local climate affecting the levels of precipitation. Precipitation decreases tree density that decreases the level of precipitation. Models that include the climate-tree feedback can produce hysteresis zones with three syndromes. These models also indicate that direct shifts between forest and the treeless syndromes are possible (Van Nes et al. Citation2014). In light of this forest-savanna dynamic, a simple monotonic model for tree density in Neotropical pastures may be unrealistic (Westoby, Walker, and Noy-Meir Citation1989). Rather, a hysteretic model may be more probable.
The first challenge in constructing this frame of reference is objectively defining the variables involved, which is to say, identifying the relevant drivers and the measure of arborization of the system. At the most simple level we can begin with the variables ‘density of trees’ as the response variable, and cattle density (as a depiction of disturbance) as the driving variables. As shown in , as cattle density increases, it is more likely that trees will become less desirable in the system, as represented by a slight decline in tree cover. However, the high-tree-density state system is largely resilient to perturbations due to positive feedback loops among the components of the system. For example, trees create a humid microclimate through evapotranspiration that allows more trees to thrive. As trees are eliminated (by introducing cattle), the humid conditions may persist until a new positive feedback loop between tree loss and fire (or some form of ecological erosion) leads to a new tipping point followed by a rapid transition to the treeless syndrome ().
To identify the variables involved in our framework, we review the literature on tree management on Neotropical cattle ranches. We draw from the ecological literature describing tree densities and natural regeneration processes on the cattle ranches. Social drivers of tree management in the Neotropical agricultural land have been documented from the perspectives of conservation biology (see, e.g., Lerner et al. Citation2017; Pagiola et al. Citation2007), agroforestry systems (see, e.g., Mercer Citation2004; Pattanayak et al. Citation2003), and land-use change science (see, e.g., Perz and Walker Citation2002). While we draw from these disciplines, we find them limited in scope, and ultimately draw from Chayanov’s Theory of Peasant Economy to characterize the socioeconomic drivers. Chayanov’s (Citation1986) perspective is focused on farmers’ goals of prioritizing households’ needs and incrementing production accordingly (Steckley and Weis Citation2016). Chayanov’s approach helps understand farmers’ and ranchers’ responses to capitalist relations in agriculture (Steckly and Weis 2016). We propose that these relations drive the tree densities found in the farm and ranches of the Neotropics.
In the next sections we will describe the variables that compose the low- and high-tree-density syndromes and drivers. We will also discuss the positive feedback loops that may keep the syndrome from changing.
The Syndrome
Tree density is the most obvious variable associated with our ability to identify one or another syndrome. But it is not the only one. How trees are arranged spatially, and the diversity of trees are also characteristics of the syndromes. Tree spatial arrangements can be categorized into seven distinct patterns: dispersed trees in pasture or agricultural plots, live fences, riparian vegetation, forest fragments, fallow vegetation, horticulture area, and forestry plantations (Caughlin et al. Citation2016; Chazdon Citation2003; Harvey and Haber Citation1998). Dispersed trees are either planted, the result of natural regeneration, or remnant trees from a previous forest. A live fence is a row of trees connected by barbed wire employed to separate properties and paddocks within properties. Live fences are mostly planted trees, and require a lot of care (Harvey and Haber Citation1998). Riparian vegetation includes tree groupings found along streams and rivers that aid in water conservation. Forest fragments are remnants of larger tracts of forests. Fallow vegetation is vegetation that farmers allow to grow temporarily to renew the soil. Sometimes ranchers and farmers have clusters of trees in gardens (generally close to the house), and tree plantations for timber (for example, teak, mahogany), or for commercial fruits (for example, Brazil nut) (Simmons, Walker, and Wood Citation2002). Each of the seven categories has different purposes and together they comprise the tree density of the farm and ranch.
Trees in agro-silvo-pastoral systems are managed by the farmer who consequently directly determines the diversity by: (1) allowing certain species to remain standing after forest clearing (Harvey and Haber Citation1998); (2) managing regeneration of certain species (Lerner et al. Citation2015; Love and Spaner Citation2005; Ramírez-Marcial et al. Citation2012; Somarriba et al. Citation2012); or (3) directly planting trees (Love and Spaner Citation2005; Simmons, Walker, and Wood Citation2002). Forest clearing and managing regeneration seem to be more important methods for maintaining dispersed trees in pastures than direct planting (Lerner et al. Citation2015; Sanfiorenzo-Barnhard et al. Citation2009). Farmers select those tree species allowed to regenerate (Harvey et al. Citation2011; Lerner et al. Citation2015; Ramírez-Marcial et al. Citation2012). Living fences and forestry plantations are commonly planted (Love and Spaner Citation2005), whereas natural regeneration is more common for maintaining riparian vegetation and forest fragments.
Shade for cattle is one of the main reasons for retaining trees in pastures (Harvey and Haber Citation1998; Lerner et al. Citation2015; Love and Spaner Citation2005; Sanfiorenzo-Barnhard et al. Citation2009), but trees in the farms and ranches have multiple uses, such as timber, fruit crops (Garen et al. Citation2011; Harvey et al. Citation2011; Lerner et al. Citation2015), and fodder (Love and Spaner Citation2005). Ecological, aesthetic (Garen et al. Citation2011; Lerner et al. Citation2015) and other values have also been attached to these trees, and farmers’ selectivity for particular tree species may be reflected in the diversity of trees found in the farm.
Ecological and management drivers
Tree regeneration and the tree structural diversity
The structural diversity of trees refers to the tree density, diversity, and canopy structure. In Neotropical silvo-pastoral systems it includes live fences, dispersed trees on pastures, riparian vegetation, forest fragments, and forestry plantations. The level of tree structural diversity is the most important factor allowing tree community regeneration to continue after disturbance. For example, areas of pasture that are closer to a forest, riparian areas, or that have a lot of dispersed trees in pastures will regenerate a lot faster than areas far away from any tree presence (Chazdon Citation2003; Griscom et al. Citation2011). Tree structural diversity has a very important role in succession as demonstrated in a 20-year post-agricultural succession plot, where remnant trees had the largest effect on the species composition of the surrounding forest (Sandor and Chazdon Citation2014).
If a silvo-pastoral system has high tree density and diversity it will more easily recover from disturbance both through resprouting, and seed dispersal (Boucher et al. Citation2001; Ferguson et al. Citation2003) processes. Seed banks appear to not contribute much to tree regeneration in the cattle pastures because they are nearly absent from the pasture soils that have been so heavily eroded. Consequently, resprouting can be a more common process of regeneration than seed germination after disturbance both in forested and pasture habitats (Griscom, Griscom, and Ashton Citation2009; Vieira and Scariot Citation2006). Both below-ground and above-ground sprout tissue can be important for regeneration depending on the habitat and level of disturbance (Kammesheidt Citation1999).
Animals are critical for tree regeneration because they disperse seeds to open habitats. Open habitats rarely attract dispersing animals unless there is structural diversity in the pastures (Guevara, Purata, and Van Der Maarel Citation1986; Vieira, Uhl, and Nepstad Citation1994; Galindo‐González, Guevara, and Sosa 2000; Slocum and Horvitz Citation2000; Chazdon Citation2003; Ferguson et al. Citation2003). Insects, mammals, and birds have been found to correlate with structural diversity in agroforestry systems (Harvey and Haber Citation1998; Harvey et al. Citation2006; Perfecto and Vandermeer Citation2002; Philpott and Dietsch Citation2003; Van Bael et al. Citation2007). For example, dispersed trees and live fences provide the structure for birds and mammals to move within agricultural landscapes (Estrada and Coates-Estrada Citation2001; Perfecto and Vandermeer Citation2008; Stoner Citation2001).
Finally, deep and extended tree roots reduce the runoff of surface water and allow better absorption of humidity and nutrients into the soil and they reduce the effect of compaction that comes with raising cattle (Joffre, Rambal, and Ratte Citation1999; Ogden et al. Citation2013). Long exposure of the land to the disturbance factors that eliminates trees leads to soil degradation by elimination of top soil layers with the majority of nutrients accessible to plants. This long exposure also reduces the ability of the land to conserve water. These degraded microclimatic conditions limit the ability of seeds to thrive once they’ve been dispersed to a particular pasture site (Vieira, Holl, and Peneireiro Citation2009). Ultimately, there are feedbacks among the ecological components of the agroecosystem that help maintain a high tree density state stable.
Disturbance caused by cattle pasture management
Disturbance during farm management is a negative driver of tree density (Muchagata and Brown Citation2003). Disturbance, in the context of pasture management, maintains low vegetation, mainly grasses. We can summarize disturbance into three processes: cattle herbivory and trampling of seedlings, competition between introduced grasses and tree seedlings, and the various forms of weeding.
Cattle trampling by browsing and stomping can negatively affect tree seedling growth rate and survival (Griscom, Griscom, and Ashton Citation2009; Guevara, Purata, and Van Der Maarel Citation1986; Harvey and Haber Citation1998). Cattle roaming the land also compact the soil, increasing runoff during the rainy periods and creating poor soil conditions for seedling growth (Chazdon Citation2003). But cattle’s effect on seedling growth and survival is not all negative. For example, cattle can facilitate the regeneration of some trees by dispersing seeds into areas that are unreachable without the cattle (Miceli-Méndez, Ferguson, and Ramírez-Marcial Citation2008). Cattle also eat grasses that can fuel fire (Janzen Citation1988). For the purpose of maintaining a healthy grass, some farmers manage the stocking rate employing paddock rotation (Broom, Galindo, and Murgueitio Citation2013). The effect of cattle on the vegetation depends on the stocking rate, which ultimately depends on the farmer or rancher. Rangeland research has focused on the effect of cattle on grass species (Westoby, Walker, and Noy-Meir Citation1989), but less on the effect of cattle on tree seedling growth and survival in active pastures.
Competition with grasses and forbs also limits tree establishment in pastures. The dense root systems of grasses compete for moisture and nutrients with the trees and thick grass canopies limit light availability for seedlings (Nepstad et al. Citation1996). Grasses are so aggressive that some suggest it is necessary for shrub species to colonize pastures before trees. Shrubs can shade out grasses and create microhabitats that facilitate the establishment of tree species (Aide et al. Citation1995). However, grasses can also have positive effects, protecting seedlings from desiccation during the dry season (Gerhardt Citation1993; Holl Citation1999; Knoop and Walker Citation1985). Thus, the effect of grasses on tree seedling establishment is dynamic.
Farmers use herbicide, manual or mechanical weeding, and fire to manage grasses and control weeds, many of which are trees in the early stages of growth. The goal is to maintain pasture grasses and eliminate woody vegetation and other weeds. Continuous application of herbicide has dire consequences for tree growth as it leads to soil moisture loss and eliminates vegetation capable of resprouting (Griscom, Ashton, and Berlyn Citation2005). Mechanical or manual weeding directly eliminates trees, seedlings, and saplings. However, mechanical weeding can be selective, favoring some tree species and leaving roots capable of resprouting. Fire is also used to create and maintain pastures and savannas (Murphy and Lugo Citation1986). Fires favor fire-dependent grasses and plants with the capacity to resprout. Pastures severely exposed to fires can lose their capacity to regenerate tree species that can’t easily resprout after the fires (Cavelier et al. Citation1998; Griscom et al. Citation2011).
Ecological positive feedback loops
Nonetheless, natural regeneration in these open habitats plays a more important role than direct tree planting by farmers (Harvey et al. Citation2011; Lerner et al. Citation2015; Ramírez-Marcial et al. Citation2012), often because tree planting efforts are too labor intensive and require technical knowledge (Holl Citation1999).
Similar to the forest/savanna example presented previously, the structural diversity is a component of the silvo-pastoral system that leads to positive feedbacks that maintain stable tree communities by attracting seed dispersers and maintaining soil and microclimate conditions. The more structural diversity, the easier it will be for the tree community to recover after disturbance. However, with more and more disturbance, a critical tree density may be reached and the high tree density silvo-pastoral system will no longer be stable. At this critical tree density, the system rapidly transitions to a treeless pasture ().
Farmers may find the task of increasing the tree density on land that has been heavily disturbed through weeding, cattle trampling and the introduction of exotic grasses to be difficult because the land is no longer suited for tree establishment. Positive feedbacks between soil degradation and lack of seed dispersal events keep treeless pasture treeless. Thus a lot of effort may be necessary to reach the critical threshold necessary to transition towards the high tree density silvo-pastoral system ().
Factors maintaining a stable equilibrium in the treeless pastures are as follows: the dry microclimate, grass dominance, soil degradation, fire, and lack of seed dispersal. The inverse of these factors maintains the silvo-pastoral syndrome.
Socioeconomic drivers
Chayanovian economics
At the beginning of the 19th century the majority of the Russian population was peasant, or subsistence farmers. Lenin theorized that class differentiation was unavoidable with industrialization, and argued that the countryside would be split into large capitalist farms driven by surplus value and farm workers (or labor) (Bernstein Citation2009). Chayanov, a Russian rural economist in the early 1900s, developed a different perspective from his extensive study of the Russian peasantry, which he developed into the Theory of Peasant Economy (Chayanov Citation1986). Chayanov (Citation1986), having spent most of his career working with the peasantry, was concerned with the actual process of decision-making within the peasant families (Bernstein Citation2009; van der Ploeg Citation2013). He claimed that the peasantry (or family farm) was not necessarily going to become a capitalist farm, considering that the peasant farm works very differently than the capitalist farm.
Chayanov (Citation1986) argued that capitalist farms aim to produce a surplus, while peasant or family farms produce enough to ensure survival. The capitalist farm is governed by maximizing profits, while the peasant farm is governed by a balance between the consumption or satisfaction pattern of the family, and the labor necessary to supply that consumption or satisfaction (see ). In the peasant farm, the amount of labor that goes into production is determined by the size of the family, and the consumption patterns of the family. In a capitalist farm, labor is determined by the market price and the amount of capital available, making labor indistinguishable from other commodities.
Table 2. Concepts of the Chayanovian balances framework.
The balance between consumption (or family satisfaction) and labor leads to another balance, the balance between drudgery and utility (Chayanov Citation1986). Utility refers to the ‘possibility to satisfy needs and aspirations’ (van der Ploeg Citation2014, 1007) and drudgery refers to ‘the energy and hardship needed to realize a particular level of production’ (van der Ploeg Citation2014, 1007). The marginal rate of utility decreases ‘per unit of production as the total level of production grows’ (van der Ploeg Citation2014, 1007), while drudgery ‘increases with the further growth of total production’ (van der Ploeg Citation2014, 1007). The main take away from Chayanov is the qualitative idea that the peasants can redefine drudgery and utility. For example, a farmer that wants to increase production because it will open possibilities, may perceive a task less drudgerous, and work harder (van der Ploeg Citation2014). Chayanov wrote about many other balances that were later conceptualized by van der Ploeg (Citation2013), including the balance of autonomy with dependency, production with reproduction, and external with internal resources.
Lenin thought that differences in incomes across peasant households were due to a transition from peasant agriculture to capitalism. Chayanov said that the income differences were a reflection of household demography, with variations in the consumer-to-laborer ratio at different stages of the household life cycle (see, e.g., Perz ad Walker 2002). The Russian peasants that Chayanov described did not pay for wage labor, but did participate in the market, own property, and had other jobs besides farming (Perz and Walker Citation2002). Clearly, this is not the case of all peasant communities – for example there are farmers who worked very close to cities, or did not own land. Nevertheless, the value of Chayanov’s model is in the observation that there is a qualitative difference among farmers who are peasant-like and those that are capitalist (van der Ploeg Citation2014).
Continuum from peasant balances to capitalism
van der Ploeg (Citation2013) conceptualizes many other balances described by Chayanov, emphasizing the importance of the balance between autonomy and dependency. Peasant farmers balance autonomy with dependency as they resist the forces that strip away their autonomy or ability to balance utility-drudgery, and capitalism is one of those forces. Since capitalist farms are highly specialized to produce commodity crops, they are dependent on the market for inputs (fertilizers, pesticides, etc.), wage labor, and even land. Peasant farms are grounded on social and natural resources, and don’t rely on the commodity market for land, external inputs or wage labor. Peasant farmers’ autonomy from the market is what allows them to balance utility-drudgery. Oostindie, van der Ploeg, and Broekhuizen (2013) found that among 1,000 Dutch dairy farmers, the peasant-like farms showed greater resilience to reduction in the market price of milk than the capitalist farms. The resilience of peasant farms to market fluctuations stems from their ability to balance utility with drudgery (van der Ploeg Citation2014). If there is a drop in price of a crop, a capitalist farm may operate at a loss and go out of business. A peasant farm under the same situation can re-define drudgery and produce more. Of course, peasant farmers can only increase production, and therefore improve their livelihood, if they have the space and means to balance utility-drudgery. For example, if a farm is being stripped of its water resources or land, the farmer can no longer balance the utility of the water with the drudgery of conserving that water (van der Ploeg Citation2014).
Similar to the Dutch dairy farmers, we expect that farmers and ranchers that manage cattle in the Neotropics vary depending on how reliant they are on the market for inputs, wage labor, and others. We propose that the utility-drudgery balance drives the tree densities across the farms. In peasant farms trees provide various utilities that are necessary to reproduce the farm, and the utility of trees (soil fertilization, cattle fodder, etc.) is balanced with drudgery of managing trees. The capitalist that specializes in raising cattle is not concerned with the utilities of trees that are necessary to reproduce the farm. Instead, tree functions are replaced with artificial items that are often commodities themselves (van der Ploeg 2009). From soil to labor, everything is bought in the capitalist farm and the Chayanovian balances are irrelevant. We propose the X axis on and , can be envisioned as a spectrum of farms according to the degree to which they balance utility-drudgery.
Tree utility and the Chayanovian balances
In addition to the utility – drudgery and autonomy – dependency balances, farmers balance internal with external farm resources, and production with reproduction. The incorporation of trees in the farm and ranch reflects farm diversification because farmers have trees for various uses in addition to crop or livestock management (Garen et al. Citation2011; Harvey et al. Citation2011; Lerner et al. Citation2015). Incorporating these trees can require drudgery to plant, eliminate weeds, and prune (Holl Citation1999). Farmers exert this effort because the trees will eventually provide resources that are necessary to reproduce the farm operation (see, e.g., Garen et al. Citation2011; Harvey et al. Citation2011, Citation2005; Lerner et al. Citation2015; Love and Spaner Citation2005). In addition, when farmers make the decision to manage trees, they are balancing production with reproduction because trees take a long time to produce utility. Often, farmers don’t think they will profit from tree planting but see this activity as a way to improve the farm for their children (Simmons, Walker, and Wood Citation2002; Pattanayak 2003; Lerner et al. Citation2015).
Chayanovian balances are reflected in farmers’ management of live fences. In Nicaragua and Costa Rica, farmers employ live fences to establish boundaries in their farm. These farmers choose the live fence over electric fences or dead wood posts because it is self-sustaining (Harvey et al. Citation2005). Therefore, farmers are balancing the live fence (or internal resource) with the electric fence (or external resource). Live fences are also multifunctional as they can be sold to neighbors, as well as provide forage, timber, and firewood (Harvey et al. Citation2005). As such, live fences are not just an internal resource to the farm, they are also necessary for the reproduction of the farm operation, and to provide some autonomy from markets for wood and other resources.
Through the Chayanovian balances, the peasant farmer can redefine the utility of trees and the drudgery of tree management (van der Ploeg Citation2013). Of course some capitalist ranchers and farmers may have trees in their farms but only to the extent that they don’t obstruct profit maximization. Unlike the peasant farmer, the capitalist farmer can’t redefine the utility of trees and if the trees obstruct profit maximization than they will be eliminated. Through these different economic logics we expect to see differences in tree densities between peasant and capitalist farms. Vennet, Schneider, and Dessein (Citation2016) demonstrated that among soy producers in Brazil, perceptions about nature can vary. Different farming styles emerged from farmers’ perceptions of nature either as a functional or a non-functional element of the farm. Such differences in perceptions are likely to be found among cattle ranchers and can be assessed employing the Chayanovian balances framework.
The peasant and capitalist farmers and rancher of the Neotropics
Based on their economy and history of settlement, distinctions among cattle ranchers have been made. For example, in the Amazon region of Brazil ranchers are often differentiated into large cattle ranchers (fazendeiros), medium (fazendeirinhos), and small (colonos) (Hecht Citation1985; Hoelle Citation2015; Muchagata and Brown Citation2003; Perz and Walker Citation2002; Taravella and De Sartre Citation2012; Walker et al. Citation2009). The large rancher has been depicted as having the power to increase production through access to technology, capital, and political influence. They are considered entrepreneurial (Taravella and De Sartre Citation2012). Chayanovian theory was applied to the case of small farm families in the Brazilian Amazon by Walker and Homma (Citation1996). Depending on the amount of household labor farmers may grow more trees vs. raising cattle (Perz and Walker Citation2002; Walker and Homma Citation1996). The small farm families may hire labor, have access to credit, and participate in the market, but cattle is typically used as a savings bank and only sold in times of need (Muchagata and Brown Citation2003; Hoelle 2011). Similar distinctions have been made in other regions of Latin America (see, e.g., Coomes et al. Citation2008; Grandia Citation2009; Heckadon 2009). Based on our literature review there is a range of economic logics across the cattle ranchers and farmers of the Neotropics.
Social positive feedback loops
Peasant farms are more likely to favor tree cover because they are more interested in diversifying production for income and subsistence as opposed to maximizing profit through commodity production. In the process of diversifying, farmers and ranchers are more likely to find utility in trees, and to explore and experiment with that utility. Feedback between farm diversification and tree utility maintains the high tree densities or silvo-pastoral syndrome stable. When the goal of the farmer is to maximize profit through cattle rearing, the farm goes from diversification to specialization. The farmer eliminates trees as they become negligible in the process of farm management. A positive feedback between farm specialization and tree elimination maintains the farm as a treeless pasture.
Unstable equilibria are possible when the peasant farmer becomes more ‘capitalist’, taking more financial risk with the aim of generating profit. Examples include mechanization of dairy production, or mortgaging their farm to obtain bank loans. In the process the peasant farmer may begin to specialize, moving along the continuum from governance by Chayanovian balances toward capitalism and profit maximizing. The farmer may eliminate trees, but still able to bounce back to the high tree density silvo-pastoral syndrome as long as the ecological conditions hold. The problem is that as the farmer specializes they lose their ability to balance utility-drudgery as they become economically trapped and dependent on the market. The farmer may continue to eliminate trees until reaching a critical tree density and rapidly transitioning into the treeless pasture syndrome.
Conclusion: a new integrative silvo-pastoral research framework and its implications
Across the Neotropics managers, academics, and farmers engage in research and practice that aims at transforming treeless cattle pastures (Lerner et al. Citation2017; Soler et al. 2018). A transition from treeless pastures to silvo-pastoral systems may be a major agricultural transformation that requires change at multiple human and environmental scales. To know where managers should intervene it’s imperative to ask first what we know about these socioecological systems (Hobbs et al. Citation2011; Suding and Hobbs Citation2009). We propose that the Chayanovian balances drive silvo-pastoral management and that multivalued dynamics ( and ) are the most likely dynamic behind the silvo-pastoral transformation. Silvo-pastoral transformation requires research that integrates many kinds of academic disciplines and participants (Sherren and Darnhofer Citation2017). We hope to advance this discussion by combining social and ecological approaches in our conceptual model of tree dynamics.
The first specific objective was to identify the dependent and independent variables involved in the dynamics. This review is focused on variables related to tree cover, and not much attention has been given to livestock management, the main source of income for cattle ranchers (Soler et al. Citation2018). In addition, the focus has been on individually held land, but land held in commons should also be included in this research program (Sherren and Darnhofer Citation2017).
Farming style research frameworks can be implemented to identify the variables that make up the silvo-pastoral and treeless pasture syndrome (Vennet, Schneider, and Dessein Citation2016). Grazing management techniques and cultural repertoires (among others) that may act in conjunction with tree densities must be identified and measured. Qualitative research that includes farmers can be implemented to identify the other syndrome variables. Farmers can be recruited as knowledge co-creators and in-depth interviews can be employed to identify the most relevant syndrome variables (Sherren and Darnhofer Citation2017).
The second specific objective was to conceptualize the Chayanovian balances as drivers of tree cover. This idea can be tested but first it’s necessary to measure the extent of the Chayanovian balances (or independent variable). Qualitative analytical methods can be used to develop a list of farmer perceptions and how these relate to the concept of the Chayanovian balances. These initial results can be discussed with farmers, allowing researchers to assess initial assumptions about the Chayanovian balances (Sherren and Darnhofer Citation2017). Qualitative research can inform the design of large-scale social surveys (Miles, Huberman, and Saldana Citation2013). Constructing a quantitative scale of the drivers is also necessary to identify the dynamic model that best describes the system.
The main objective of this review was to explore how the dynamics framework shown in can help understand the extent and emergence of the silvo-pastoral and treeless pasture syndrome. Detecting thresholds and hysteresis requires long term data sets that show abrupt changes across a time series, but these data sets are often hard to obtain. An alternative is to use spatial patterns (Suding and Hobbs Citation2009). Threshold dynamics in may be detected by testing for nonlinear spatial patterns of tree densities across farms that vary in how farmers balance utility-drudgery, production-reproduction, autonomy-dependency, and internal–external resources. The extent of the syndrome can be assessed through the use of remote sensing data that captures the tree cover of farms (Caughlin et al. Citation2016) in combination with field observations (Lerner et al. Citation2015; Sanfiorenzo-Barnhard et al. Citation2009) across farm properties and large landscapes. Tree cover can be classified further according to spatial arrangements (Caughlin et al. Citation2016; Harvey et al. Citation2011). Ideally, this would be done across the entirety of the Neotropics in areas where rainforest once stood but that are currently dominated by cattle ranching. An analysis of the distribution of the tree cover across farms is likely to show multimodal distributions coinciding with different styles of tree management across the landscape (Hirota et al. Citation2011; Staver, Archibald, and Levin Citation2011). Statistical analysis can be used to define tree cover modes and regression models can test the probability of being in one or another tree cover style as a function of the Chayanovian balances (Hirota et al. Citation2011). Hysteretic dynamics can be inferred if multimodality is present at an intermediate range of the Chayanovian balances scale, with unimodal distribution at the extremes of the Chayanovian balances scale (Hirota et al. Citation2011).
The presence or absence of tipping points and hysteresis requires different kinds of management actions. If the dynamics are monotonic (,)), a simple change in the driver variable may lead to a desired outcome. If the dynamics have tipping points and hysteresis, leverage points related to the positive feedback loops may be the best place for managers to exert effort (Kéfi, Holgrem, and Scheffer Citation2016; Suding and Hobbs Citation2009). In this review, we’ve identified some of the most important ecological and farm management variables and processes that directly shape tree densities in the farm. Possible ecological and social positive feedback loops at the agroecosystem level have also been described. Understanding the interactions among farm management and/or ecological variables can help uncover the presence or absence of positive feedback loops. Identifying the positive interactions, measured as positive correlations among management and ecological variables, can begin to uncover the mechanism by which high or low tree densities are maintained (Kéfi, Holgrem, and Scheffer Citation2016). Understanding these mechanisms can help managers and stakeholders know where to exert management efforts by identifying leverage points (Suding and Hobbs Citation2009).
A socioecological research program that focuses on understanding existing silvo-pastoral systems instead of introducing new agroforestry practices can help initiate a conversation among farmers and scientists. The idea is not to discourage agroforestry research, but to enrich our understanding of farmers’ knowledge and experiences so that research can engage with their priorities. If the Chayanovian balances drive tree densities, programs that call for silvo-pastoral systems must take into account the economic logics that promote the spontaneous silvo-pastoral systems, including farmer’s autonomy (van der Ploeg Citation2014). For example, if a farmer’s economic logic is to be independent of market inputs, bank credits or just one commodity, intensive silvo-pastoral techniques that promote these kinds of dependencies are likely to be rejected (Steckley and Weis Citation2016) or have negative consequences. Implementation programs that are built on extentionists and commodity markets as key actors create more dependencies (Mier Y Terán Giménez Cacho et al. Citation2018). A better leverage point for policy makers would be to create markets that support farmers that already manage silvo-pastoral systems and to strengthen their existing efforts (Mier Y Terán Giménez Cacho et al. Citation2018). A deeper understanding of the system dynamics, including their interacting social and ecological drivers, will play a key role in scaling up silvo-pastoral systems.
Acknowledgements
We are grateful to the University of Michigan Department of Ecology and Evolutionary Biology for helping accomplish this research. We thank Catalina Esguerra, Shannon Dowd, Elise Hernandez, Paul Glaum, Trevor Caughlin, Catherine Badgley, Lesli Hoey, Ivette Perfecto, Maria Carmen Lemos and two anonymous reviewers for their comments on previous versions of this manuscript.
References
- Aide, T. M., J. K. Zimmerman, L. Herrera, M. Rosario, and M. Serrano. 1995. Forest recovery in abandoned tropical pastures in Puerto Rico. Forest Ecology and Management 77(1–3):77–86. doi:10.1016/0378-1127(95)03576-V.
- Andow, D. A., and K. Hidaka. 1989. Experimental natural history of sustainable agriculture: Syndromes of production. Agriculture, Ecosystems & Environment 27(1–4):447–62. doi:10.1016/0167-8809(89)90105-9.
- Bernstein, H. 2009. VI Lenin and AV Chayanov: looking back, looking forward. The journal of peasant studies 36(1):55–81. doi:10.1080/03066150902820289.
- Boucher, D. H., J. H. Vandermeer, I. G. De La Cerda, M. A. Mallona, I. Perfecto, and N. Zamora. 2001. Post-agriculture versus post-hurricane succession in southeastern Nicaraguan rain forest. Plant Ecology 156(2):131–37. doi:10.1023/A:1012672005360.
- Broom, D. M., F. A. Galindo, and E. Murgueitio. 2013. Sustainable, efficient livestock production with high biodiversity and good welfare for animals. Proceedings of the Royal Society Biological Sciences 280 (1771):Article Number UNSP 20132025. doi:10.1098/rspb.2013.2025
- Cale, P., and B. H. Allen-Diaz. 2013. New models for ecosystem dynamics and restoration. Washington, DC: Island Press.
- Calle, A., F. Montagnini, and A. F. Zuluaga. 2009. Farmer’s perceptions of silvopastoral system promotion in Quindío, Colombia. Bois et forets des tropiques 300(2):79–94. doi:10.19182/bft2009.300.a20417.
- Calle, Z., E. Murgueitio, J. Chará, C. H. Molina, A. F. Zuluaga, and A. Calle. 2013. A strategy for scaling-up Intensive silvopastoral systems in Colombia. Journal of sustainable forestry 32(7):677–93. doi:10.1080/10549811.2013.817338.
- Caughlin, T. T., S. W. Rifai, S. J. Graves, G. P. Asner, and S. A. Bohlman. 2016. Integrating LiDAR‐derived tree height and Landsat satellite reflectance to estimate forest regrowth in a tropical agricultural landscape. Remote Sensing in Ecology and Conservation 2(4):190–203. doi:10.1002/rse2.33.
- Cavelier, J., T. Aide, C. Santos, A. Eusse, and J. Dupuy. 1998. The savannization of moist forests in the Sierra Nevada de Santa Marta, Colombia. Journal of Biogeography 25(5):901–12. doi:10.1046/j.1365-2699.1998.00222.x.
- Chayanov, A. V. 1986. A.V. Chayanov On the Theory of Peasant Economy. In Madison, Wis, ed. Kerblay, and R. E. F. Smith. Madison, WI: University of Wisconsin Press.
- Chazdon, R. L. 2003. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspectives in Plant Ecology, Evolution and Systematics 6(1):51–71. doi:10.1078/1433-8319-00042.
- Coomes, O. T., F. Grimard, C. Potvin, and P. Sima. 2008. The fate of the tropical forest: Carbon or cattle? Ecological Economics 65:207–12. doi:10.1016/j.ecolecon.2007.12.028.
- Dagang, A. B., and P. Nair. 2003. Silvopastoral research and adoption in Central America: Recent findings and recommendations for future directions. Agroforestry Systems 59(2):149–55. doi:10.1023/A:1026394019808.
- Estrada, A., and R. Coates-Estrada. 2001. Bat species richness in live fences and in corridors of residual rain forest vegetation at Los Tuxtlas, Mexico. Ecography 24(1):94–102. doi:10.1034/j.1600-0587.2001.240111.x.
- Ferguson, B. G., J. Vandermeer, H. Morales, and D. M. Griffith. 2003. Post‐agricultural succession in El Petén, Guatemala. Conservation Biology 17(3):818–28. doi:10.1046/j.1523-1739.2003.01265.x.
- Galindo-González, J., S. Guevara, and V. J. Sosa. 2000. Bat‐and bird‐generated seed rains at isolated trees in pastures in a tropical rainforest. Conservation biology 14(6):1693–703. doi:10.1046/j.1523-1739.2000.99072.x.
- García-Barrio, R., and L. García-Barrios. 1990. Environmental and technological degradation in peasant agriculture: a consequence of development in Mexico. World Development 18(11):1569–85. doi:10.1016/0305-750X(90)90044-X.
- Garen, E. J., K. Saltonstall, M. S. Ashton, J. L. Slusser, S. Mathias, and J. S. Hall. 2011. The tree planting and protecting culture of cattle ranchers and small-scale agriculturalists in rural Panama: Opportunities for reforestation and land restoration. Forest Ecology and Management 261(10):1684–95. doi:10.1016/j.foreco.2010.10.011.
- Gerhardt, K. 1993. Tree seedling development in tropical dry abandoned pasture and secondary forest in Costa Rica. Journal of Vegetation Science 4(1):95–102. doi:10.2307/3235736.
- Grandia, L. 2009. Raw hides: Hegemony and cattle in Guatemala’s northern lowlands. Geoforum 40(5):720–31. doi:10.1016/j.geoforum.2009.01.004.
- Griscom, H. P., P. M. Ashton, and G. P. Berlyn. 2005. Seedling survival and growth of native tree species in pastures: implications for dry tropical forest rehabilitation in Central Panama. Forest Ecology and Management 218(1–3):306–18. doi:10.1016/j.foreco.2005.08.026.
- Griscom, H. P., A. B. Connelly, M. S. Ashton, M. H. Wishnie, and J. Deago. 2011. The structure and composition of a tropical dry forest landscape after land clearance; Azuero Peninsula, Panama. Journal of Sustainable Forestry 30(8):756–74. doi:10.1080/10549811.2011.571589.
- Griscom, H. P., B. W. Griscom, and M. Ashton. 2009. Forest regeneration from pasture in the dry tropics of Panama: effects of cattle, exotic grass, and forested riparia. Restoration Ecology 17(1):117–26. doi:10.1111/rec.2009.17.issue-1.
- Guevara, S., S. E. Purata, and E. Van Der Maarel. 1986. The role of remnant forest trees in tropical secondary succession. Vegetatio 66(2):77–84.
- Harvey, C. A., and W. A. Haber. 1998. Remnant trees and the conservation of biodiversity in Costa Rican pastures. Agroforestry Systems 44(1):37–68. doi:10.1023/A:1006122211692.
- Harvey, C. A., A. Medina, D. M. Sánchez, S. Vílchez, B. Hernández, J. C. Saenz, J. M. Maes, F. Casanoves, and F. L. Sinclair. 2006. Patterns of animal diversity in different forms of tree cover in agricultural landscapes. Ecological applications 16(5):1986–99. doi:10.1890/1051-0761(2006)016[1986:POADID]2.0.CO;2.
- Harvey, C. A., C. Villanueva, H. Esquivel, R. Gómez, M. Ibrahim, M. Lopez, J. Martinez, et al. 2011. Conservation value of dispersed tree cover threatened by pasture management. Forest Ecology and Management. 261(10):1664–74. doi:10.1016/j.foreco.2010.11.004.
- Harvey, C. A., C. Villanueva, J. Villacís, M. Chacón, D. Muñoz, M. López, M. Ibrahim, et al. 2005. Contribution of live fences to the ecological integrity of agricultural landscapes. Agriculture, Ecosystem and Environment 111:200–30. doi:10.1016/j.agee.2005.06.011.
- Hecht, S. B. 1985. Environment, Development and Politics: Capital Accumulation and the Livestock Sector in Eastern Amazonia. World Development 13(6):663–84. doi:10.1016/0305-750X(85)90114-7.
- Hirota, M., M. Holmgren, E. H. Van Nes, and M. Scheffer. 2011. Global resilience of tropical forest and avanna to critical transitions. Science 334(6053):232–35. doi:10.1126/science.1210657.
- Hobbs, R. J., L. M. Hallett, P. R. Ehrlich, and H. A. Mooney. 2011. Intervention Ecology: Applying ecological science in the Twenty-first Century. BioScience 61(6):442–50. doi:10.1525/bio.2011.61.6.6.
- Hoelle, J. 2015. Rainforest Cowboys: the rise of ranching and cattle culture in Western Amazonia. Austin, TX University of Texas Press.
- Holl, K. D. 1999. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate, and soil1. Biotropica 31(2):229–42. doi:10.1111/btp.1999.31.issue-2.
- Janzen, D. H. 1988. Tropical ecological and biocultural restoration. Science 239(4837):243–44. doi:10.1126/science.239.4837.243.
- Joffre, R., S. Rambal, and J. P. Ratte. 1999. The dehesa system of southern Spain and Portugal as a natural ecosystem mimic. Agroforestry systems 45(1/3):57–79. doi:10.1023/A:1006259402496.
- Kammesheidt, L. 1999. Forest recovery by root suckers and above-ground sprouts after slash-and- burn agriculture, fire and logging in Paraguay and Venezuela. Journal of Tropical Ecology 15(2):143–57. doi:10.1017/S0266467499000723.
- Kéfi, S., M. Holgrem, and M. Scheffer. 2016. When can positive interactions cause alternative stable states in ecosystems? Functional Ecology 30:88–97. doi:10.1111/1365-2435.12601.
- Knoop, W., and B. Walker. 1985. Interactions of woody and herbaceous vegetation in a Southern African savanna. The Journal of Ecology 235–53. doi:10.2307/2259780.
- Lerner, A. M., T. K. Rudel, L. C. Schneider, M. McGroddy, D. V. Burbano, and C. F. Mena. 2015. The spontaneous emergence of silvo-pastoral landscapes in the Ecuadorian Amazon: Patterns and processes. Regional Environmental Change 15(7):1421–143. doi:10.1007/s10113-014-0699-4.
- Lerner, A. M., A. F. Zuluaga, J. Chará, A. Etter, and T. Searchinger. 2017. Sustainable cattle ranching in practice Moving from theory to planning in Colombia´s livestock sector. Ecological Management 60:176–84.
- Lockwood, J. A., and D. R. Lockwood. 1993. Catastrophe theory: a unified paradigm for rangeland ecosystem dynamics. Journal of Range Management 282–88. doi:10.2307/4002459.
- Love, B., and D. Spaner. 2005. A survey of small-scale farmers using trees in pastures in Herrera Province, Panama. Journal of Sustainable Forestry 20(3):37–65. doi:10.1300/J091v20n03_03.
- Mather, A. S., and C. L. Needle. 1998. The forest transition: a theoretical basis. Area 30(2):117–24. doi:10.1111/area.1998.30.issue-2.
- Mercer, D. E. 2004. Adoption of agroforestry innovations in the tropics: a review. Agroforestry Systems 61(1–3):311–28.
- Miceli-Méndez, C. L., B. G. Ferguson, and N. Ramírez-Marcial. 2008. Seed dispersal by cattle: Natural history and applications to Neotropical forest restoration and agroforestry. Chapter 7 in Post-Agricultural succession in the Neotropics. New York: Springer.
- Mier Y Terán Giménez Cacho, M., O. F. Giraldo, M. Aldasoro, H. Morales, B. G. Ferguson, P. Rosset, A. Khadse, et al. 2018. Bringing agroecology to scale: key drivers and emblematic cases. Agroecology and Sustainable Food Systems. doi:10.1080/21683565.2018.1443313.
- Miles, M. B., A. M. Huberman, and J. Saldana. 2013. Qualitative data analysis. Thousand Oaks, CA: Sage.
- Moguel, P., and V. M. Toledo. 1999. Biodiversity conservation in traditional coffee systems of Mexico. Conservation Biology 13:11–21. doi:10.1046/j.1523-1739.1999.97153.x.
- Muchagata, M., and K. Brown. 2003. Cows, colonists and trees: rethinking cattle and environmental degradation in Brazilian Amazonia. Agricultural systems 76(3):797–816. doi:10.1016/S0308-521X(02)00015-X.
- Murgueitio, E., Z. Calle, F. Uribe, A. Calle, and B. Solorio. 2011. Native trees and shrubs for the productive rehabilitation of tropical cattle ranching lands. Forest Ecology and Management 261(10):1654–63. doi:10.1016/j.foreco.2010.09.027.
- Murphy, P. G., and A. E. Lugo. 1986. Ecology of tropical dry forest. Annual review of ecology and systematics 67–88. doi. org/10.1146/annurev.es.17.110186.000435
- Nepstad, D. C., C. Uhl, C. A. Pereira, and J. M. C. Da Silva. 1996. A comparative study of tree establishment in abandoned pasture and mature forest of eastern Amazonia, 25–39. Oikos.
- Ogden, F. L., T. D. Crouch, R. F. Stallard, and J. S. Hall. 2013. Effect of land cover and use on dry season river runoff, runoff efficiency, and peak storm runoff in the seasonal tropics of Central Panama. Water Resources Research 49(12):8443–62. doi:10.1002/2013WR013956.
- Oostindie, H., J. D. van der Ploeg, and R. van Broekhuizen. 2013. Buffercapaciteit: Bedrijfsstijlen in de melkveehouderij, volatiele markten en kengetallen. Wageningen: WUR/ALFA.
- Pagiola, S., E. Ramírez, J. Gobbi, C. De Haan, M. Ibrahim, E. Murgueitio, and J. P. Ruíz. 2007. Paying for the environmental services of silvopastoral practices in Nicaragua. Ecological Economics 64(2):374–85. doi:10.1016/j.ecolecon.2007.04.014.
- Pattanayak, S. K., D. E. Mercer, E. Sills, and J. C. Yang. 2003. Taking stock of agroforestry adoption studies. Agroforestry systems 57(3):173–86. doi:10.1023/A:1024809108210.
- Payne, W. 1985. A review of the possibilities for integrating cattle and tree crop production systems in the tropics. Forest ecology and management 12(1):1–36. doi:10.1016/0378-1127(85)90133-1.
- Perfecto, I., and J. Vandermeer. 2002. Quality of agroecological matrix in a tropical Montane landscape: Ants in coffee plantations in Southern Mexico. Conservation Biology 16(1):174–82. doi:10.1046/j.1523-1739.2002.99536.x.
- Perfecto, I., and J. Vandermeer. 2008. Biodiversity conservation in tropical agroecosystems. Annals of the New York Academy of Sciences 1134(1):173–200. doi:10.1196/nyas.2008.1134.issue-1.
- Perz, S. G. 2007. Grand theory and context-specificity in the study of forest dynamics: Forest transition theory and other directions. The Professional Geographer 59(1):105–14. doi:10.1111/j.1467-9272.2007.00594.x.
- Perz, S. G., and R. T. Walker. 2002. Household life cycles and secondary forest cover among small farm colonists in the Amazon. World Development 30(6):1009–27. doi:10.1016/S0305-750X(02)00024-4.
- Philpott, S. M., and T. Dietsch. 2003. Coffee and conservation: A global context and the value of farmer involvement. Conservation Biology 17(6):1844–46. doi:10.1111/cbi.2003.17.issue-6.
- Ramírez-Marcial, N., M. L. Rueda-Pérez, B. Ferguson, and G. Jiménez-Ferrer. 2012. Caracterización del sistema agrosilvopastoril en la depresión central de Chiapas. Avances en Investigación Agropecuaria 16(2):7–22.
- Rudel, T. K. 1998. Is there a forest transition? Deforestation, reforestation and development. Rural Sociology 63:533–52. doi:10.1111/j.1549-0831.1998.tb00691.x.
- Sandor, M. E., and R. L. Chazdon. 2014. Remnant trees affect species composition but not structure of tropical second-growth forest. PloS one 9(1):e83284. doi:10.1371/journal.pone.0083284.
- Sanfiorenzo-Barnhard, C., L. García-Barrios, E. Meléndez-Ackerman, and R. Trujillo-Vásquez. 2009. Woody cover and local farmers’ perceptions of active pasturelands in La Sepultura Biosphere Reserve Buffer Zone, Mexico. Mountain Research and Development 29(4):320–27. doi:10.1659/mrd.00013.
- Scheffer, M. 2009. Critical transitions in nature and society. Princeton, NJ Princeton University Press.
- Sherren, K., and I. Darnhofer. 2017. Precondition for integration: In support of stand-alone social science in Rangeland and silvopastoral research. Rangeland Ecology & Management. doi:10.1016/j.rama.2017.08.003.
- Simmons, C. S., R. T. Walker, and C. H. Wood. 2002. Tree planting by small producers in the tropics: a comparative study of Brazil and Panama. Agroforestry systems 56(2):89–105. doi:10.1023/A:1021377231402.
- Slocum, M. G., and C. C. Horvitz. 2000. Seed arrival under different genera of trees in a neotropical pasture. Plant Ecology 149(1):51–62. doi:10.1023/A:1009892821864.
- Soler, R., P. L. Peri, H. Bahamonde, V. Gargaglione, S. Ormaechea, A. H. Herrera, L. S. Jardón, C. Lorenzo, and G. M. Pastur. 2018. Assessing knowledge production for agrosilvopastoral systems in South America. Rangeland Ecology and Management. doi:10.1016/j.rama.2017.12.006.
- Somarriba, E., J. Beer, J. Alegre-Orihuela, H. J. Andrade, R. Cerda, F. DeClerck, G. Detlefsen, et al. 2012. ‘Mainstreaming agroforestry. In In Agroforestry-The Future of Global Land Use, ed. L. America.’, 429–53. Dordrecht: Springer Netherlands.
- Staver, A. C., S. Archibald, and S. A. Levin. 2011. The global extent and determinants of savanna and forest as alternative biome states. Science 334(6053):230–32. doi:10.1126/science.1210465.
- Steckley, M., and T. Weis. 2016. Peasant balances, neoliberalism, and the stunted growth of non-traditional agro-exports in Haiti. Canadian Journal of Latin American and Caribbean Studies/Revue canadienne des études latino-américaines et caraïbes 41(1):1–22. doi:10.1080/08263663.2015.1130293.
- Stoner, K. E. 2001. Differential habitat use and reproductive patterns of frugivorous bats in tropical dry forest of northwestern Costa Rica. Canadian Journal of Zoology 79(9):1626–33. doi:10.1139/z01-105.
- Suding, K. N., and R. J. Hobbs. 2009. Threshold models in restoration and conservation: a developing framework. Trends in Ecology and Evolution 24(5):271–79. doi:10.1016/j.tree.2008.11.012.
- Taravella, R., and X. A. De Sartre. 2012. The symbolic and political appropriation of scales: A critical analysis of the Amazonian ranchers’ narrative. Geoforum 43(3):645–56. doi:10.1016/j.geoforum.2011.11.009.
- Van Bael, S. A., P. Bichier, I. Ochoa, and R. Greenberg. 2007. Bird diversity in cacao farms and forest fragments of western Panama. Biodiversity and Conservation 16(8):2245–56. doi:10.1007/s10531-007-9193-3.
- van der Ploeg, J. D. 2013. The Art of Peasant Farming: A Chayanovian Manifesto. No. 2. Winnipeg, Manitoba: Fernwood.
- van der Ploeg, J. D. 2014. Peasant-driven agricultural growth and food sovereignty. The Journal of Peasant Studies 41(6):999–1030. doi:10.1080/03066150.2013.876997.
- Van Nes, E. H., B. M. S. Arani, A. Staal, B. Van Der Bolt, B. M. Flores, S. Bathiany, and M. Scheffer. 2016. What do you mean, ‘Tipping point’? Trends in Ecology and Evolution 31(12):902–04. doi:10.1016/j.tree.2016.09.011.
- Van Nes, E. H., M. Hirota, M. Holgrem, and M. Scheffer. 2014. Tipping points in tropical tree cover: linking the theory to data. Global Change Biology 20:1016–21. doi:10.1111/gcb.12398.
- Vandermeer, J. 1997. Syndromes of production: An emergent property of simple agroecosystem dynamics. Journal of Environmental Management 51(1):59–72. doi:10.1006/jema.1997.0128.
- Vandermeer, J., and I. Perfecto. 1997. The agroecosystem: a need for the conservation biologist’s lens. Conservation Biology 11(3):591–92. doi:10.1046/j.1523-1739.1997.07043.x.
- Vandermeer, J. H., and I. Perfecto. 2012. Syndromes of production in agriculture: Prospects for social-ecological regime change. Ecology and Society 17(4):39. doi:10.5751/ES-04813-170439.
- Vennet, B. V., S. Schneider, and J. Dessein. 2016. Different farming styles behind the homogenous soy production in southern Brazil. The Journal of Peasant Studies 43(2):396–418. doi:10.1080/03066150.2014.993319.
- Vieira, D. L., K. D. Holl, and F. M. Peneireiro. 2009. Agro‐successional restoration as a strategy to facilitate tropical forest recovery. Restoration Ecology 17(4):451–59. doi:10.1111/j.1526-100X.2009.00570.x.
- Vieira, D. L., and A. Scariot. 2006. Principles of natural regeneration of tropical dry forests for restoration. Restoration Ecology 14(1):11–20. doi:10.1111/j.1526-100X.2006.00100.x.
- Vieira, I. C. G., C. Uhl, and D. Nepstad. 1994. The role of the shrub Cordia multispicata Cham. as a ‘succession facilitator’ in an abandoned pasture, Paragominas, Amazonia. Vegetatio 115(2):91–99.
- Walker, R., J. Browder, E. Arima, C. Simmons, R. Pereira, M. Caldas, R. Shirota, and S. De Zen. 2009. Ranching and the new global range: Amazônia in the 21st century. Geoforum 40(5):732–45. doi:10.1016/j.geoforum.2008.10.009.
- Walker, R., and A. K. O. Homma. 1996. Land use and land cover dynamics in the Brazilian Amazon: an overview. Ecological Economics 67–80. doi:10.1016/0921-8009(96)00033-X.
- Westoby, M., B. Walker, and I. Noy-Meir. 1989. Opportunistic management for rangelands not at equilibrium. Journal of Range Management 42(4):266–74. doi:10.2307/3899492.
- Zahawi, R. A., G. Duran, and U. Kormann. 2015. Sixty-seven years of land-use change in southern Costa Rica. PloS one 10(11):e0143554. doi:10.1371/journal.pone.0143554.
- Zomer, R. J., H. Neufeldt, J. Xu, A. Ahrends, D. Bossio, A. Trabucco, M. Van Noordwijk, and M. Wang. 2016. Global tree cover and biomass carbon on agricultural land: The contribution of agroforestry to global and national carbon budgets. Scientific reports 6. doi:10.1038/srep29987