
ABSTRACT
The emergence of SARS-CoV-2 has resulted in an ongoing global pandemic with significant morbidity, mortality, and economic consequences. The susceptibility of different animal species to SARS-CoV-2 is of concern due to the potential for interspecies transmission, and the requirement for pre-clinical animal models to develop effective countermeasures. In the current study, we determined the ability of SARS-CoV-2 to (i) replicate in porcine cell lines, (ii) establish infection in domestic pigs via experimental oral/intranasal/intratracheal inoculation, and (iii) transmit to co-housed naïve sentinel pigs. SARS-CoV-2 was able to replicate in two different porcine cell lines with cytopathic effects. Interestingly, none of the SARS-CoV-2-inoculated pigs showed evidence of clinical signs, viral replication or SARS-CoV-2-specific antibody responses. Moreover, none of the sentinel pigs displayed markers of SARS-CoV-2 infection. These data indicate that although different porcine cell lines are permissive to SARS-CoV-2, five-week old pigs are not susceptible to infection via oral/intranasal/intratracheal challenge. Pigs are therefore unlikely to be significant carriers of SARS-CoV-2 and are not a suitable pre-clinical animal model to study SARS-CoV-2 pathogenesis or efficacy of respective vaccines or therapeutics.
Introduction
The emergence of the SARS-CoV-2 virus, the causative agent of COVID-19, has resulted in a global pandemic with over 20 million cases and 740,000 deaths as of August 13, 2020 [Citation1,Citation2]. SARS-CoV-2 causes a respiratory disease in humans with a broad clinical presentation, ranging from asymptomatic or mild illness to severe fatal disease with multi-organ failure [Citation3–6]. SARS-CoV-2 is rapidly transmissible via contact with infected respiratory droplets and can also be transmitted by asymptomatic carriers [Citation6–8]. To curb viral spread, countries have instituted varying levels of social distancing policies, which have significant negative economic and social impacts [Citation9]. Mitigating the effects of this unprecedented pandemic will necessitate the development of effective vaccines and therapeutics, which will require well-characterized and standardized pre-clinical animal models.
SARS-CoV-2 is a member of the Betacoronavirus genus that includes the pathogenic human viruses SARS-CoV and MERS-CoV [Citation2,Citation10–12]. While details of the origin of SARS-CoV-2 are unknown, evidence indicates it emerged from a zoonotic spillover event, with bats and perhaps pangolins as probable origin species [Citation2,Citation13–15]. The potential for a reverse zoonotic event, i.e. human-to-animal transmission, is possible and of significant concern to animal and public health [Citation16–18]. Instances of natural human-to-animal transmission of SARS-CoV-2 have been reported with COVID-19 patients in domestic settings (dogs and cats), zoos (lions and tigers), and farms (mink) [Citation18–20]. Therefore, investigations into the infectivity of SARS-CoV-2 in various animal species with human contact are essential to assess and control the risk of a spillover event and to establish the role these animals may play in the ecology of the virus.
Several studies have determined the susceptibility of different animal species to SARS-CoV-2 via experimental infection [Citation20,Citation21]. Cats, hamsters, and ferrets are highly susceptible to SARS-CoV-2 infection, demonstrate varying clinical and pathological disease manifestations, readily transmit the virus to naïve animals, and mount a virus-specific immune response [Citation22–28]. Dogs are mildly susceptible to experimental SARS-CoV-2 infection, with limited viral replication but with clear evidence of seroconversion in some animals [Citation22]. Poultry species seem to be resistant to SARS-CoV-2 infection [Citation22,Citation26]. These findings establish the respective utility of different animal species as pre-clinical models to study SARS-CoV-2.
Several lines of evidence suggest that pigs could be susceptible to SARS-CoV-2 infection. Pigs are susceptible to both experimental and natural infection with the related betacoronavirus, SARS-CoV, and demonstrate seroconversion [Citation29,Citation30]. Structure-based analyses predict that the SARS-CoV-2 Spike (S) protein receptor binding domain (RBD) binds the pig angiotensin-converting enzyme 2 (ACE2) entry receptor with similar efficiency compared to human ACE2 [Citation31]. Single-cell screening also indicates that pigs co-express ACE2 and the protease TMPRSS2 (viral activating factor) in a variety of different cell lines, and SARS-CoV-2 replicates in various pig cell lines [Citation2,Citation26,Citation32,Citation33]. Despite these preliminary data indicating that pigs could be susceptible to SARS-CoV-2 infection, two recent studies revealed that intranasal inoculation of three and twelve pigs, respectively, with 105 pfu or TCID50 of SARS-CoV-2 did not lead to any detectable viral replication or seroconversion [Citation22,Citation26]. However, the single route of intranasal inoculation used in these studies suggests that additional investigations are necessary before definitive conclusions can be made regarding susceptibility of pigs to SARS-CoV-2.
In the present study, we determined the susceptibility of swine cell lines and domestic pigs to SARS-CoV-2 infection. Two different porcine cell lines were found to be permissive to SARS-CoV-2 infection showing cytopathic effects (CPE). Domestic pigs were challenged via simultaneous oral/intranasal/intratracheal inoculation with a 106 TCID50 dose of SARS-CoV-2. SARS-CoV-2 did not replicate in pigs and none of them seroconverted. Furthermore, the virus was not transmitted from SARS-CoV-2 inoculated animals to sentinels. The present findings, combined with the other studies [Citation22,Citation26], confirm that pigs seem resistant to SARS-CoV-2 infection despite clear susceptibility of porcine cell lines. Pigs are therefore unlikely to play an important role in the COVID-19 pandemic as a virus reservoir or as a pre-clinical animal model to study SARS-CoV-2 pathogenesis or develop novel countermeasures.
Materials and Methods
Virus and cells
SARS-CoV-2 USA-WA1/2020 isolate (GenBank accession # MN985325) [Citation34] was obtained from BEI resources (catalog # NR-52281, American Type Culture Collection [ATCC®]. Manassas, VA, USA). The virus was passaged three times in VeroE6 cells (ATCC® CRL-1586™), before being passaged two times in swine testicle (ST; ATCC CRL-1746™) and four times in porcine kidney (PK-15; ATCC® CCL-33™) cell lines to investigate suitability of these swine cell lines for propagation of SARS-CoV-2. The first passage of the SARS-CoV-2 virus from ST cells was used to prepare the challenge material. The titre of the virus inoculum stock was 2.5 × 105 TCID50/mL All experiments involving the SARS-CoV-2 virus were performed under Biosafety Level (BSL) 3+ conditions at the Biosecurity Research Institute (BRI) at Kansas State University (KSU), Manhattan, KS, USA.
Outline of animal experiments
Animal infection experiments using swine were performed under BSL-3Ag conditions at the BRI at KSU. Animal research was conducted in compliance with the Animal Welfare Act and other federal statutes and regulations relating to animal care and experimentation under protocol #4390, approved by the Institutional Animal Care and Use Committee (IACUC) at Kansas State University on April 8, 2020.
Eighteen pigs (mix of males and females, five weeks of age) were used in the study. Pigs were acquired from a source guaranteed free of swine influenza virus (SIV), porcine circovirus-2 (PCV-2), and porcine reproductive and respiratory syndrome virus (PRRSV) infection. The study outline is illustrated in . Upon arrival, pigs were acclimated for 3 days prior to SARS-CoV-2 inoculation. Nine pigs were designated as uninfected negative controls and housed in separate BSL-2 facilities. Three of these uninfected negative control pigs were humanely euthanized at 3 days post challenge (DPC) to provide negative control clinical and tissue samples. Nine principal infected pigs were housed in the same room in two separate groups (4 or 5 pigs each; ) and were infected with 1 x106 TCID50 of SARS-CoV-2 orally (1 mL), intranasally (1 ml; 0.5 ml each nostril) and intratracheally (2 mL), after being anesthetized with a mixture of telazol/xylazine. Three sentinel contact pigs were added to each group on 1 DPC (). Rectal temperature and signs of clinical disease for each pig were determined daily throughout the study; clinical signs include overall activity/attitude (signs of depression, decreased alertness or unresponsiveness), appetite (based on interest in treats), respiratory signs (sneezing, coughing, laboured breathing, nasal discharge), and digestive signs (diarrohea or vomiting). Blood samples and nasal, oropharyngeal, and rectal swabs were collected in virus transport medium (VTM; DMEM plus antibiotic/antimycotic) at 0, 1, 3, 5, 7, 10, 14, and 21 DPC. As summarized in and , pigs were humanely euthanized for scheduled post mortem examinations on 4 DPC (3 principal infected pigs), 8 DPC (3 principal infected pigs), and 21 DPC (remaining 3 principal infected and 6 sentinel pigs) to collect respiratory tissue samples, with blood and swab samples collected on pigs prior to euthanasia. Gross pathological examinations on major organs were performed and respiratory tissue samples were collected and either stored in 10% neutral-buffered formalin or stored as fresh samples at −80°C. Blood and swab samples were all filtered using a 0.2 µm filter prior to storage at −80°C.
Figure 1. Study Design. Eighteen pigs were placed into three groups. Group 1 (principal infected animals) consisted of nine pigs (four and five in each pen) and was inoculated via intranasal (IN), oral (PO), and intratracheal (IT) routes simultaneously with a total dose of 1 × 106 TCID50 of SARS-CoV-2 in 4 mL DMEM. The pigs in Group 2 (n = 6; sentinel contact animals) and Group 3 (n = 3; mock control animals) were housed in a separate room. At 1-day post challenge (DPC), the six pigs in Group 2 were co-mingled with the principal infected animals in Group 1 (three pigs per pen) and served as sentinel contact controls. The remaining three pigs in Group 3 remained in separate housing and served as mock-infected negative controls and were euthanized and necropsied on 3 DPC. Principal infected animals were euthanized and necropsied at 4 (n = 3), 8 (n = 3), and 21 (n = 3) DPC to determine the course of infection. All six sentinel pigs were also euthanized on 21 DPC.
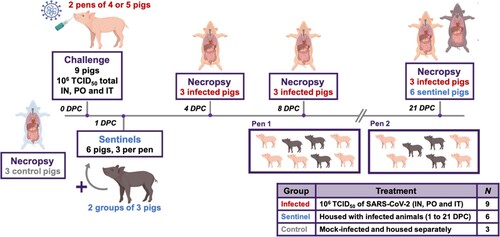
Table 1. Animal groups.
RNA extraction and reverse transcription quantitative PCR (RT-qPCR)
RNA was isolated from blood, swabs, and tissue samples using a magnetic bead-based protocol in a BSL-3+ laboratory at the BRI at KSU. Lung tissue homogenates (200 mg per 1 mL DMEM; 20% w/v) were prepared by thawing tissue, mincing it into 1 mm sections, followed by lysis in a 2 mL sure-lock tube containing 5 mm stainless steel homogenization beads using the TissueLyser LT (Qiagen, Germantown, MD, USA) for 30 s at 30 hz followed by 1 min of 30 hz while keeping the sample cold. Following clarification via a 3-minute centrifugation (3,000xg; room temperature), supernatants were mixed with an equal volume of RLT lysis buffer. Blood and clinical swabs were directly mixed with an equal volume of RLT lysis buffer. Two hundred microliters of each sample lysate were used to extract RNA using a magnetic bead-based nucleic acid extraction kit (GeneReach USA, Lexington, MA) on an automated TacoMiniTM nucleic acid extraction system (GeneReach USA, Lexington, MA) according to the manufacturer’s protocol with the following modifications: beads were added to the sample well, followed by the sample lysate, then 200 µL molecular grade isopropanol (ThermoFisher Scientific, Waltham, MA, USA). The last wash buffer was replaced by molecular grade 200 proof ethanol (ThermoFisher Scientific, Waltham, MA, USA). Extraction positive controls (IDT, IA, USA; 2019-nCoV_N_Positive Control, diluted 1:100 in RLT buffer) and negative controls were employed. Extracted RNA was eluted in 100 µL buffer.
Reverse transcription quantitative PCR (RT-qPCR) was performed to detect viral RNA using the CDC standard N2-based SARS-CoV-2 detection assay [Citation35] that was validated for use with the qScript XLT 1-Step RT-qPCR ToughMix (Quantabio, Beverly, MA, USA) on a CFX96 real-time thermocycler (Bio-Rad, Hercules, CA, USA) using a 20 min RT step and 45 cycle qPCR in a 20 µL reaction volume. RT-qPCR on each sample was performed in duplicate wells with a quantitated PCR positive control (IDT 2019-nCoV–N-Positive Control, diluted 1:100) and four non-template negative controls on every plate. A positive Ct cut-off of <37 cycles was used. A plasmid template including the SARS-CoV-2 N gene was used as a positive PCR amplification control. A 10-point standard curve using quantitated stock viral RNA (USA-WA1/2020 isolate) was used to quantify RNA copy number.
Gross pathology and histopathology
During post mortem examinations, the upper and lower respiratory tract, central nervous system, lymphatic and cardiovascular systems, gastrointestinal and urogenital systems, and integument were evaluated. Lungs were removed in toto and the percentage of the lung surface that was affected by macroscopic lesions was estimated by a single veterinarian experienced in evaluating gross porcine lung pathology as previously described [Citation36,Citation37]. Lungs were evaluated for gross pathology such as edema, congestion, discolouration, atelectasis, and consolidation. Lung tissue samples from right cranial, middle, and caudal lobes were collected and either fixed in 10% neutral-buffered formalin for histopathological examination or frozen at −80°C for RT-qPCR testing. Tissues were fixed in formalin for 7 days, then transferred to 70% ethanol (ThermoFisher Scientific, Waltham, MA, USA) prior to trimming and paraffin embedding following standard automated protocols used in the histology section of the Kansas State Veterinary Diagnostic Laboratory. Following embedding, tissue sections were cut and stained with hematoxylin and eosin and evaluated by a board-certified veterinary pathologist who was blinded to the treatment groups.
For RNAscope® in situ hybridization (ISH), an anti-sense probe targeting the nucleocapsid protein (N; nucleotide sequence 28,274-29,533) of SARS-CoV-2 USA-WA1/2020 isolate (GenBank accession # MN985325) was designed (Advanced Cell Diagnostics (ACD), Newark, CA, USA) and used as previously described [Citation38]. Four micron thick sections of formalin-fixed paraffin-embedded tissues were mounted on Superfrost® Plus slides (VWR, Radnor, PA). The RNAscope® ISH assay was performed using the RNAscope 2.5 LSx Reagent Kit (ACD) on the automated BOND RXm platform (Leica Biosystems, Buffalo Grove, IL, USA). Briefly, sections were subjected to automated baking and deparaffinization followed by heat-induced epitope retrieval (HIER) using an EDTA-based solution (pH 9.0; Leica Biosystems) at 100°C for 15 min. Tissue sections were then protease treated with RNAscope® 2.5 LSx Protease for 15 min at 40 °C followed by hydrogen peroxide treatment for 10 min at room temperature. Slides were incubated with the probe mixture for 2 h at 40 °C, and the signal was amplified using a specific set of amplifiers (AMP1 through AMP6) as recommended by the manufacturer. The signal was detected using a Fast-Red solution for 10 min at room temperature. Slides were counterstained with a ready-to-use hematoxylin for 5 min, followed by five washes with 1X BOND Wash Solution (Leica Biosystems). Slides were finally rinsed in deionized water, dried in a 60 °C oven for 30 min, and mounted with Ecomount® (Biocare, Concord, CA, USA). Lung sections from a SARS-CoV-2-infected hamster were used as positive assay controls.
For immunohistochemistry (IHC), four micron thick sections of formalin-fixed paraffin-embedded tissue were mounted on Superfrost® Plus slides and subjected to IHC using a SARS-CoV-2-specific anti-nucleocapsid rabbit polyclonal antibody (rabbit antibody #3A, developed by our laboratory) with the method previously described [Citation38]. IHC was performed using the automated BOND-RXm platform and the Polymer Refine Red Detection kit (Leica Biosystems). Following automated deparaffinization, HIER was performed using a citrate-based solution (pH 6.0; Leica Biosystems) at 100 °C for 20 min. Sections were then incubated with the primary antibody (1:5,000 in primary antibody diluent [Leica Biosystems]) for 30 min at room temperature, followed by a polymer-labeled goat anti-rabbit IgG coupled with alkaline phosphatase (30 min). Fast Red was used as the chromogen (15 min), and counterstaining was performed with hematoxylin for 5 min. Slides were dried in a 60 °C oven for 30 min and mounted with a permanent mounting medium (Micromount®, Leica Biosystems). Lung sections from a SARS-CoV-2-infected hamster were used as positive assay controls.
Serological testing
To detect SARS-CoV-2 antibodies in sera, indirect ELISAs were performed using recombinant SARS-CoV-2 Receptor Binding Domain (RBD) expressed in HEK cells with a C-terminal Strep-tag and the Nucleocapsid (N) protein expressed in E.coli with a C-terminal His-tag. Briefly, the recombinant proteins were produced in their respective expression systems according to standard procedures and purified using either Ni-NTA (ThermoFisher Scientific, Waltham, MA, USA) or Strep-Tactin (IBA Lifesciences, Goettingen, Germany) columns according to manufacturer’s instructions. For testing, 96-well plates were coated with 100 ng of the recombinant protein in 100 µL of sodium carbonate/bicarbonate buffer and incubated overnight at 4°C. The wells were then washed and blocked with casein blocking buffer. Sera were diluted 1:200 in 100 µL of casein blocking buffer, incubated at room temperature for 1 h, then washed with washing solution. One hundred microliters of anti-pig-IgG or IgM secondary antibodies, conjugated with horseradish peroxidase, and diluted 1:2,500 in blocking buffer were then incubated at room temperature for 1 h, protected from light. TMB colorimetric substrate was added and incubated at room temperature for 5 min, then the reaction was stopped with a solution of 0.2 sulfuric acid. The optical density (OD) value was measured at 450 nm within 5 min of adding the stop solution to quantify the amount of antigen-binding antibody present in the sample. Sera from mock-infected pigs were used as negative controls. Sera collected from SARS-CoV-2 infected cats, from a different study [Citation39], were used as positive controls. Serum from pigs infected with African Swine Fever Virus (ASFV) and the baculovirus-expressed ASFV-p54 antigen were used as positive control for anti-pig-IgG or IgM antibodies. The cutoff for a sample being called positive was defined by the average OD at 0 DPC +3x standard deviation.
The presence of virus-neutralizing antibodies in sera was determined via microneutralization assay. Serum samples were diluted 1:10 and heat-inactivated at 56°C for 30 min while shaking. Subsequently, 100 µL of serum samples per well in duplicate were subjected to 2-fold serial dilutions starting at 1:20 through 1:2560 in 100 µL culture media. One hundred microliters of 100 TCID50 of SARS-CoV-2 was then added to 100 µL of the sera dilutions and incubated for 1 h at 37°C, followed by culture of the mixture on VeroE6 cells in 96-well plates. Results of the virus neutralization were determined by the appearance of CPE, which was observed under a microscope at 96 h post inoculation. The neutralizing antibody titre is determined as the reciprocal of the average serum dilution at which no CPE breakthrough in any of the testing wells is observed. Neutralizing sera from SARS-CoV-2-infected cats from a separate study [Citation39] were used as positive controls.
Next generation sequencing
To determine the consensus sequence of the USA-WA/1/2020 virus and to analyse if there were any nucleic acid substitutions in the SARS-CoV-2 virus consensus sequence after passage in porcine cell lines, RNA was extracted from cell culture supernatant as described above. The RNA was then subjected to RT–PCR amplification using a tiled-primer approach to amplify the entire SARS-CoV-2 genome as described previously [Citation40]. Briefly, the PCR amplicons were pooled and subjected to library preparation for Next Generation Sequencing using the Nextera XT library prep kit (Illumina, San Diego, CA, USA). The library was normalized and sequenced using a MiSeq nano v2 2 × 250 sequencing kit. The sequence was then analysed by mapping reads to the parent sequence (Genbank accession # MN985325) [Citation34] to generate a consensus sequence.
Results
Propagation of SARS-CoV-2 in swine cells
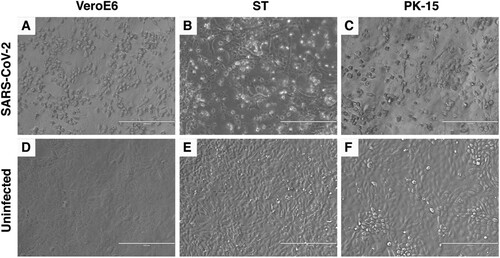
The SARS-CoV-2 USA-WA1/2020 isolate, which was isolated from a human patient in Washington State, USA, was used as the parent stock for the study [Citation34]. The virus stock was passaged 3 times at a multiplicity of infection (MOI) between 0.001 and 0.01 in VeroE6 cells and NGS sequenced before being inoculated onto porcine cells. The consensus sequence of the VeroE6-passaged virus was 100% identical to the GenBank reference sequence (GenBank accession # MN985325). To determine the susceptibility of porcine cell lines to SARS-CoV-2 infection, swine testicle (ST) and porcine kidney (PK-15) cell lines were inoculated with approximately 0.05 MOI of passage 3 of the VeroE6-passaged SARS-CoV-2 USA-WA/1/2020 isolate. No obvious cytopathic effect (CPE) was observed in either cell line during the first passage, however clear CPE was observed in passage two in ST cells and passage four in PK-15 cells (). Cell culture supernatant from SARS-CoV-2 passage one and two on ST cells, and passage three on PK-15 cells was collected, and RNA extracted for sequencing. Next generation sequencing was performed to generate a consensus genomic sequence for the ST- and PK-15-passaged SARS-CoV-2 virus. No nucleotide mutations or amino acid substitutions within the consensus sequence were observed upon passage in the two porcine cell lines. These results indicate that SARS-CoV-2 is able to infect porcine kidney and testicle cells without the requirement of major genetic adaptations in the consensus sequence. SARS-CoV-2 from passage 1 in ST cells was used as challenge material for the pig inoculation since a sufficient volume and titre was available only from passage 1 at the time of challenge.
Figure 2. Cytopathic effect (CPE) of SARS-CoV-2 in Swine Testicle (ST) and Porcine Kidney (PK-15) cells. SARS-CoV-2 USA-WA1/2020 isolate was passaged in ST and PK-15 porcine cells. Passage two in ST cells (B) and passage four in PK-15 cells (C) resulted in clear CPE, similar to that observed in permissive VeroE6 cells (A). No CPE is observed in the uninfected VeroE6 or porcine cells lines (D, E, F).
Oral/intranasal/intratracheal inoculation of pigs with SARS-CoV-2
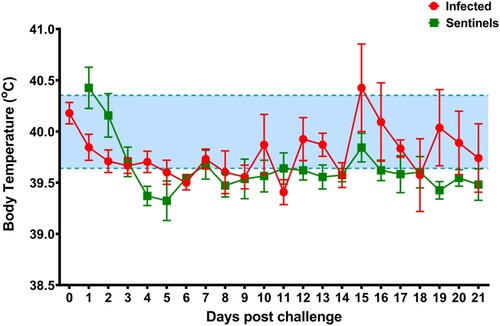
To determine the effect of SARS-CoV-2 infection in domestic pigs, nine five-week-old SARS-CoV-2 seronegative piglets were inoculated with a total of 1 × 106 TCID50 of the USA-WA1/2020 isolate, which was passaged once in swine ST cells (). The challenge material (total 4 mL) was administered orally (1 mL), intranasally (1 mL; 0.5 mL each nostril) and intratracheally (2 mL) after sedation of the animals. At 1-day post challenge (DPC), six uninoculated sentinel contact pigs were co-mingled with the principal inoculated animals (3 animals per pen). Daily rectal temperatures were recorded for each pig and clinical signs were monitored daily, including observations for signs of lethargy, hyporexia, respiratory distress (coughing, laboured breathing, nasal discharge), and digestive issues (diarrohea or vomiting). No significant change in rectal temperature was observed in the principal inoculated nor sentinel contact pigs throughout the study (). Moreover, no obvious clinical signs were observed for any of the principal inoculated nor sentinel pigs throughout the 21-day observation period.
Figure 3. Average daily rectal temperatures of SARS-CoV-2 inoculated and sentinel pigs. Daily average rectal temperatures of pigs inoculated orally, intranasally, and intratracheally with SARS-CoV-2 (red) and co-mingled sentinel pigs (green) showed no significant change over the course of the experiment. The baseline temperature (blue; 39.6°C to 40.4°C) was determined from all pigs before infection.
To detect viral replication in the principal and sentinel pigs, clinical samples were subjected to RT-qPCR to detect the SARS-CoV-2 N gene () [Citation35]. Blood, as well as oropharyngeal, nasal, and rectal swabs, were collected at various time points throughout the study and upper and lower respiratory tract tissues were collected at post-mortem examinations on 4, 8 and 21 DPC. RT-qPCR failed to detect any viral RNA in any swab or blood sample for the duration of the study (). The only exception was a nasal swab sample at 1 DPC in the principal inoculated pig #161, for which one of two RT-qPCR replicates yielded a low fluorescent amplification curve with a Ct of 37.72, which is slightly above the Ct cut-off of >37 and thus considered a negative sample (). Moreover, viral RNA was not detected in any lung sample collected at post-mortem examination on 4, 8 and 21 DPC (). In addition, gross and histopathological analysis of lung from the principal challenged pigs, including immunohistochemical and in situ hybridization analyses specific for the SARS-CoV-2 nucleocapsid (N) protein, did not reveal the presence of any obvious pathological lesions or the presence of SARS-CoV-2 antigen or RNA (, ). These results indicate that SARS-CoV-2 failed to replicate in the respiratory and digestive tract as well as the blood in orally/intranasally/intratracheally inoculated pigs throughout an observation period of 21 days. This is confirmed by the fact that the principal infected pigs failed to transmit SARS-CoV-2 to co-mingled sentinel animals.
Figure 4. Histopathological analysis of pig lung tissue. Lung tissue sections were stained with hematoxylin and eosin for histopathological evaluation (H&E, left panels). Immunohistochemistry (IHC, middle panels) analysis was done using a rabbit anti-SARS-CoV-2 nucleocapsid polyclonal antibody and in situ hybridization (ISH, right panels) analysis using an anti-sense probe to detect nucleocapsid-specific RNA. (A) Uninoculated negative control pig #210, (B) SARS-CoV-2-inoculated pig #161, 4 DPC, (C) SARS-CoV-2-inoculated pig #803, 8 DPC, (D) SARS-CoV-2-inoculated pig #193, 21 DPC. No significant histopathology and no detection of SARS-CoV-2 antigen or RNA were observed by IHC or ISH. Lung sections from a SARS-CoV-2-infected hamster were used as positive assay controls for ISH and IHC (data not shown). Magnification is 10x for all images.
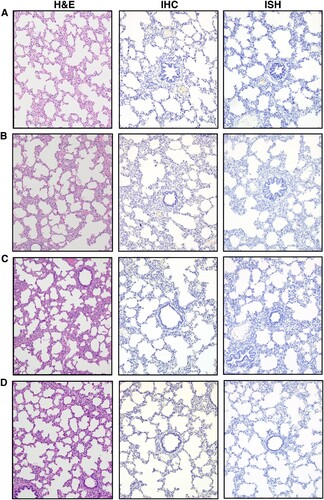
Table 2. Summary of RT-qPCR results.
Table 3. Macroscopic lesions of total lung (%).
Absence of immune response in SARS-CoV-2-inoculated pigs
To determine whether the orally/intranasally/intratracheally inoculated pigs developed an immune response against SARS-CoV-2 antigens, sera collected from infected pigs at various time points post infection were subjected to indirect ELISAs to detect IgG and IgM antibodies reactive against the receptor-binding domain (RBD) of the SARS-CoV-2 spike protein and the SARS-CoV-2 nucleocapsid (N) protein. Cumulatively, the results from these assays indicated that the infected pigs did not develop an IgG or IgM immune response against either of the SARS-CoV-2 antigens at any point throughout the study (). The sera from one sentinel pig (#894) showed transient IgM reactivity on 3 and 5 DPC against N and RBD and an isolated IgG reactivity against N and RBD on 3 DPC. Importantly, neutralizing antibody experiments performed with sera collected from principal inoculated and sentinel contact pigs necropsied at 14 and 21 DPC, revealed that none of the pigs developed neutralizing antibodies against SARS-CoV-2. Overall, these results indicate that SARS-CoV-2 is unable to replicate and generate an immune response in pigs upon oral/intranasal/intratracheal inoculation. In addition, transmission to sentinel contact animals was not possible.
Figure 5. Serological response in pigs infected orally/intranasally/intratracheally with SARS-CoV-2 and sentinel contact pigs. Indirect ELISAs were performed against the SARS-CoV-2 antigens N (nucleocapsid protein [A, B]) and RBD (Spike protein receptor binding domain [C, D]) to detect antigen-specific IgG (A, C) or IgM (B, D) antibodies. Sera reactivity was determined for three principal infected pigs (#893 (red), #193 (dark red), #219 (orange)) and three sentinel pigs (#848 (blue), #172 (cyan), #894 dark blue)). The cutoff for a positive sample was determined by +3 standard deviations of 0 DPC samples (dotted line). Feline SARS-CoV-2-specific antibodies were used as positive controls from a separate study (left bar, [Citation38]). Porcine positive control sera for IgG/IgM-specific antibodies were ASFV-infected pig sera detecting the ASFV-p54 antigen (right bar). Uninfected pigs were used as negative controls. O.D. – optical density.
![Figure 5. Serological response in pigs infected orally/intranasally/intratracheally with SARS-CoV-2 and sentinel contact pigs. Indirect ELISAs were performed against the SARS-CoV-2 antigens N (nucleocapsid protein [A, B]) and RBD (Spike protein receptor binding domain [C, D]) to detect antigen-specific IgG (A, C) or IgM (B, D) antibodies. Sera reactivity was determined for three principal infected pigs (#893 (red), #193 (dark red), #219 (orange)) and three sentinel pigs (#848 (blue), #172 (cyan), #894 dark blue)). The cutoff for a positive sample was determined by +3 standard deviations of 0 DPC samples (dotted line). Feline SARS-CoV-2-specific antibodies were used as positive controls from a separate study (left bar, [Citation38]). Porcine positive control sera for IgG/IgM-specific antibodies were ASFV-infected pig sera detecting the ASFV-p54 antigen (right bar). Uninfected pigs were used as negative controls. O.D. – optical density.](/cms/asset/818390ff-8b74-420b-a21d-c77b92937046/temi_a_1831405_f0005_oc.jpg)
Discussion
SARS-CoV-2 is a zoonotic agent, and a detailed understanding of the susceptibility of various animal species to SARS-CoV-2 is central to controlling its spread [Citation16,Citation17]. In addition, the development of animal models that emulate COVID-19 in humans is essential for pre-clinical testing of novel vaccines and therapeutics [Citation20]. In this study, we inoculated nine pigs with a high dose of SARS-CoV-2 that was passaged once in porcine cells. Simultaneous oral/intranasal/intratracheal inoculation did not result in any detectable viral RNA in the blood, the oral/nasal/rectal cavities, or the lungs. Also, none of the co-mingled, sentinel contact pigs shed viral RNA. Moreover, a virus-specific immune response characteristic of SARS-CoV-2 infection was not observed within the 21-day study period in the principal infected or sentinel pigs. The transient nature of the IgM and IgG response observed in pig #848, particularly against the N protein, could indicate cross-reactivity of antibodies directed against another porcine coronavirus such as porcine epidemic diarrohea virus (PEDV), transmissible gastroenteritis virus (TGEV), or porcine respiratory coronavirus (PRCV) [Citation41–43]. A two-way antigenic cross-reactivity between SARS-CoV and TGEV/PRCV has been documented previously and attributed specifically to the N protein, which is 90% identical between SARS-CoV and SARS-CoV-2 [Citation44–46]. Such antibodies could be maternally derived and therefore transient as the lack of SARS-CoV-2 specific reactivity by the end of the study might suggest. In contrast to previous SARS-CoV-2 swine studies [Citation22,Citation26], the present study used a more stringent inoculation procedure (intratracheal and oral, in addition to intranasal) and a 1 log higher titre of virus inoculum (106 vs 105). In addition, the inoculum in the present study was passaged once in porcine ST cells. These results, combined with previous intranasal pig inoculation studies [Citation22,Citation26], indicate that young pigs seem to be resistant to SARS-CoV-2 infection, are unlikely to be a SARS-CoV-2 carrier animal species, and are also not suitable as an animal model for COVID-19 research.
The results of the present and previous SARS-CoV-2 inoculation studies in pigs are intriguing in light of the findings that the porcine ACE2 receptor seems highly compatible with the SARS-CoV-2 RBD, suggesting that pigs could be susceptible to SARS-CoV-2 infection [Citation2,Citation31]. Pigs are susceptible to both experimental and natural infection with SARS-CoV [Citation29,Citation30]. However, the experimental SARS-CoV infection was via simultaneous intranasal/oral/intraocular/intravenous inoculation [Citation29], thus the actual route(s) of SARS-CoV infection cannot be determined. Recently, several porcine cell lines have been shown to be permissive to SARS-CoV-2 infection [Citation26,Citation33]; in addition, single-cell screening studies showed that porcine ACE2/TMPRSS2 expression are compatible with infection [Citation32]. In contrast to previous reports that some porcine cell lines are susceptible to SARS-CoV-2 infection, but show no CPE [Citation26,Citation33], we found that both ST and PK-15 cell lines are susceptible to infection and observed CPE after two or four passages, respectively. The delay in observed CPE until after several passages is likely due to the lower susceptibility of swine cells to the virus compared to VeroE6, combined with the low MOI used for the first blind passage (0.05 MOI). The absence of SARS-CoV-2 replication and transmission in the present and two previous pig studies [Citation22,Citation26] seems to lessen the need to monitor pig populations for SARS-CoV-2 during the ongoing pandemic. However, the evidence described above suggesting pig susceptibility should not be disregarded, because all pig studies to date have used rather young and healthy pigs, and commercially available pig breeds/genetics; increased age, different breeds, or co-morbidities could make pigs more susceptible to infection. We also have to be aware that unforeseen genetic changes in the SARS-CoV-2 genome may result in a better compatibility of the virus for pigs in the future.
Pigs are considered to be an excellent model for studying human infectious diseases based on their relatedness to humans in terms of anatomy and immune responses and they have been found to be much more predictive for the efficacy of therapeutics when compared to rodent models [Citation47]. However, the results presented here indicate that pigs are not a suitable preclinical model for SARS-CoV-2 pathogenesis studies and the development and efficacy testing of therapeutics and/or vaccines. A recently available article indicates that while pigs are not susceptible to SARS-CoV-2 infection, neutralizing antibody responses were detected in pigs infected via intramuscular or intravenous inoculation routes [Citation48]; this indicates that pigs could be used for immunogenicity studies related to SARS-CoV-2. However, the use of pigs to monitor for SARS-CoV-2 immune responses must be carefully planned to avoid detection of cross-reactive antibodies specific for porcine coronaviruses [Citation43]. Alternate pre-clinical animal models, namely non-human primates, Syrian hamsters, transgenic or transduced mice expressing human ACE2, ferrets, or even cats need to be considered to gain additional insights into SARS-CoV-2 pathogenesis and virulence. Comprehensive characterization of SARS-CoV-2 pathogenesis in pre-clinical animal models and the establishment of standardized infection and testing protocols will be crucial for the development of much-need countermeasures to combat COVID-19.
Meekins_et_al_Supplemental_Materials-clean.docx
Download ()Acknowledgements
We gratefully thank the staff of KSU Biosecurity Research Institute, the histological laboratory at the Kansas State Veterinary Diagnostic Laboratory (KSVDL), the CMG staff and Gleyder Roman-Sosa, Yonghai Li, Konner Cool, Emily Gilbert-Esparza, and Chester McDowell at KSU. We also gratefully acknowledge the members of the Histology and Immunohistochemistry section at the Louisiana Animal Disease Diagnostic Laboratory (LADDL) for their assistance on slide preparation and staining. The following reagent was obtained through BEI Resources, National Institute of Allergy and Infectious Diseases (NIAID), National Institutes of Health (NIH): SARS-CoV-2 Virus strain USA-WA1/2020 (catalogue # NR-52281).
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Organization WH. (2020). Coronavirus disease (COVID-19) Situation report - 206.
- Zhou P, Yang XL, Wang XG, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020 Mar;579(7798):270–273.
- Macera M, De Angelis G, Sagnelli C, et al. Clinical presentation of COVID-19: Case Series and Review of the Literature. Int J Environ Res Public Health. 2020 Jul 14;17(14):5062.
- Chen N, Zhou M, Dong X, et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. Lancet. 2020 Feb 15;395(10223):507–513.
- Guan WJ, Ni ZY, Hu Y, et al. Clinical Characteristics of coronavirus disease 2019 in China. N Engl J Med. 2020 Apr 30;382(18):1708–1720.
- Wiersinga WJ, Rhodes A, Cheng AC, et al. Pathophysiology, transmission, Diagnosis, and treatment of coronavirus disease 2019 (COVID-19): A Review. JAMA. 2020 Aug 25;324(8):782–793.
- Li Q, Guan X, Wu P, et al. Early transmission Dynamics in Wuhan, China, of novel coronavirus-infected Pneumonia. N Engl J Med. 2020 Mar 26;382(13):1199–1207.
- He X, Lau EHY, Wu P, et al. Temporal dynamics in viral shedding and transmissibility of COVID-19. Nat Med. 2020 May;26(5):672–675.
- Nicola M, Alsafi Z, Sohrabi C, et al. The socio-economic implications of the coronavirus pandemic (COVID-19): A review. Int J Surg. 2020 Jun;78:185–193.
- Coronaviridae Study Group of the International Committee on Taxonomy of V. The species severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Microbiol. 2020 Apr;5(4):536–544.
- Vijayanand P, Wilkins E, Woodhead M. Severe acute respiratory syndrome (SARS): a review. Clin Med (Lond). 2004 Mar-Apr;4(2):152–160.
- Ramadan N, Shaib H. Middle East respiratory syndrome coronavirus (MERS-CoV): A review. Germs. 2019 Mar;9(1):35–42.
- Andersen KG, Rambaut A, Lipkin WI, et al. The proximal origin of SARS-CoV-2. Nat Med. 2020 Apr;26(4):450–452.
- Zhang T, Wu Q, Zhang Z. Probable Pangolin origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Curr Biol. 2020 Apr 20;30(8):1578.
- Lau SKP, Luk HKH, Wong ACP, et al. Possible Bat origin of severe Acute respiratory syndrome coronavirus 2. Emerg Infect Dis. 2020 Jul;26(7):1542–1547.
- Gollakner R, Capua I. Is COVID-19 the first pandemic that evolves into a panzootic? Vet Ital. 2020 Apr 24;56(1):7–8.
- Yoo HS, Yoo D. COVID-19 and veterinarians for one health, zoonotic- and reverse-zoonotic transmissions. J Vet Sci. 2020 May;21(3):e51.
- McNamara T, Richt JA, Glickman L. A Critical Needs Assessment for Research in Companion animals and Livestock following the pandemic of COVID-19 in humans. Vector Borne Zoonotic Dis. 2020 Jun;20(6):393–405.
- Hernandez M, Abad D, Eiros JM, et al. Are animals a Neglected transmission route of SARS-CoV-2? Pathogens. 2020 Jun 18;9(6):480.
- Abdel-Moneim AS, Abdelwhab EM. Evidence for SARS-CoV-2 infection of animal Hosts. Pathogens. 2020 Jun 30;9(7):529.
- Sarkar J, Guha R. Infectivity, virulence, pathogenicity, host-pathogen interactions of SARS and SARS-CoV-2 in experimental animals: a systematic review. Vet Res Commun. 2020 Jul 10;1–10.
- Shi J, Wen Z, Zhong G, et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science. 2020 May 29;368(6494):1016–1020.
- Halfmann PJ, Hatta M, Chiba S, et al. Transmission of SARS-CoV-2 in domestic cats. N Engl J Med. 2020 Aug 6;383(6):592–594.
- Kim YI, Kim SG, Kim SM, et al. Infection and Rapid transmission of SARS-CoV-2 in ferrets. Cell Host Microbe. 2020 May 13;27(5):704–709 e2.
- Richard M, Kok A, de Meulder D, et al. SARS-CoV-2 is transmitted via contact and via the air between ferrets. Nat Commun. 2020 Jul 8;11(1):3496.
- Schlottau K, Rissmann M, Graaf A, et al. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: an experimental transmission study. Lancet Microbe. 2020 Sep;1(5):e218–e225.
- Chan JF, Zhang AJ, Yuan S, et al. Simulation of the clinical and pathological manifestations of coronavirus disease 2019 (COVID-19) in golden Syrian hamster model: implications for disease pathogenesis and transmissibility. Clin Infect Dis. 2020 Mar 26;ciaa325.
- Sia SF, Yan LM, Chin AWH, et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature. 2020 Jul;583(7818)0:834–838.
- Weingartl HM, Copps J, Drebot MA, et al. Susceptibility of pigs and chickens to SARS coronavirus. Emerg Infect Dis. 2004 Feb;10(2):179–184.
- Chen W, Yan M, Yang L, et al. SARS-associated coronavirus transmitted from human to pig. Emerg Infect Dis. 2005 Mar;11(3):446–448.
- Wan Y, Shang J, Graham R, et al. Receptor Recognition by the novel coronavirus from Wuhan: an analysis based on Decade-Long Structural studies of SARS coronavirus. J Virol. 2020 Mar 17;94(7).
- Chen D, Sun J, Zhu J, et al. Single-cell screening of SARS-CoV-2 target cells in pets, livestock, poultry and wildlife. Biorxiv (Preliminary). 2020.
- Chu H, Fuk-Woo Chan J, Tsz-Tai Yuen T, et al. Comparative tropism, replication kinetics, and cell damage profiling of SARS-CoV-2 and SARs-CoV with implications for clinical manifestations, transmissibility, and laboratory studies of COVID-19: an observational study. Lancet Microbe. 2020 May;1(1):e14–e23.
- Harcourt J, Tamin A, Lu X, et al. Severe Acute respiratory syndrome coronavirus 2 from patient with coronavirus disease, United States. Emerg Infect Dis. 2020 Jun;26(6):1266–1273.
- Centers for Disease Control and Prevention RVB, Division of Viral Diseases. (2020). Real-Time RT-PCR Panel for Detection - 2019-Novel Coronavirus.
- Richt JA, Lager KM, Janke BH, et al. Pathogenic and antigenic properties of phylogenetically distinct reassortant H3N2 swine influenza viruses cocirculating in the United States. J Clin Microbiol. 2003 Jul;41(7):3198–3205.
- Lee J, Wang L, Palinski R, et al. Comparison of Pathogenicity and Transmissibility of influenza B and D viruses in pigs. Viruses. 2019 Sep 27;11(10):905.
- Carossino M, Ip HS, Richt JA, et al. Detection of SARS-CoV-2 by RNAscope((R)) in situ hybridization and immunohistochemistry techniques. Arch Virol. 2020 Oct;165(10):2373–2377.
- Gaudreault NN, Trujillo JD, Carossino M, et al. SARS-CoV-2 infection, disease and transmission in domestic cats. Emerg Microbes Infect. 2020 Oct 8;1–36.
- Quick J. (2020). nCov-2019 sequencing protocol v2 V.2. protocols.io https://dx.doi.org/10.17504/protocols.io.bdp7i5rn
- Langel SN, Paim FC, Lager KM, et al. Lactogenic immunity and vaccines for porcine epidemic diarrhea virus (PEDV): Historical and current concepts. Virus Res. 2016 Dec 2;226:93–107.
- Sestak K, Lanza I, Park SK, et al. Contribution of passive immunity to porcine respiratory coronavirus to protection against transmissible gastroenteritis virus challenge exposure in suckling pigs. Am J Vet Res. 1996 May;57(5):664–671.
- Wang Q, Vlasova AN, Kenney SP, et al. Emerging and re-emerging coronaviruses in pigs. Curr Opin Virol. 2019 Feb;34:39–49.
- Vlasova AN, Zhang X, Hasoksuz M, et al. Two-way antigenic cross-reactivity between severe acute respiratory syndrome coronavirus (SARS-CoV) and group 1 animal CoVs is mediated through an antigenic site in the N-terminal region of the SARS-CoV nucleoprotein. J Virol. 2007 Dec;81(24):13365–13377.
- Sun ZF, Meng XJ. Antigenic cross-reactivity between the nucleocapsid protein of severe acute respiratory syndrome (SARS) coronavirus and polyclonal antisera of antigenic group I animal coronaviruses: implications for SARS diagnosis. J Clin Microbiol. 2004;42:2351–2352.
- Okba NMA, Muller MA, Li W, et al. Severe Acute respiratory syndrome coronavirus 2-specific antibody responses in coronavirus disease patients. Emerg Infect Dis. 2020 Jul;26(7):1478–1488.
- Meurens F, Summerfield A, Nauwynck H, et al. The pig: a model for human infectious diseases. Trends Microbiol. 2012 Jan;20(1):50–57.
- Vergara-Alert J, Rodon J, Carrillo J, et al. (2020). Pigs are not susceptible to SARS-CoV-2 infection but are a model for viral immunogenicity studies.