
ABSTRACT
This study deals with biology and life table parameters of the predatory mite, Cheyletus malaccensis Oudemans were evaluated on three astigmatid mites as prey (i.e. Acarus siro (Linnaeus), Caloglyphus berlesei (Michael) and Tyrophagus putrescentiae (Schrank)) under laboratory conditions. Three constant temperatures (22, 27 and 32 ± 2°C) and constant relative humidity 80 ± 5% were used. Statistical analysis indicated significant difference between prey types and the three different temperatures. The shortest life cycle was recorded on A. siro (11.60 and 8.0 days) at 32°C, while the longest was on C. berlesei (29.5 and 21.2) at 22°C for females and males, respectively. Longest female longevity was on A. siro (43.6 days) at 22°C and shortest was 20.65 days on T. putrescentiae at 32°C. Highest fecundity was found on A. siro (196.50 eggs/female) at 27°C and lowest one was 69.10 eggs/female on C. berlesei at 22°C. The highest net reproduction rate of increase (R0) was 113.2 ♀/♀/generation at 27°C on A. siro. Obtained results suggested that C. malaccensis could develop and reproduce within a wide range of temperatures and prey types. It can also be used as a biological control agent to reduce the amount of acarid mite pests that infest stored commodities.
Introduction
Stored product mites pollute stored grain products used for human consumption, cheeses, animal feed, and dried fruit [Citation1]. The stored product mites change the quality of infected food by the production of secrets and feces [Citation2]. The mites become an important pest of stored products when populations reach high densities at humid climate. The level of damage caused by mites is related to the size of the population that, in turn, depends on how rapidly the population is able to increase [Citation3].
The acarid mite, Tyrophagus putrescentiae (Schrank) inhabits different natural habitats, infesting many commodities, such as wheat, dried fruits, mushrooms, oil seeds, cheese, dried ham and grain debris [Citation1]. Acarus siro (Linnaeus), also known as the flour mite is an economically important pest of stored food products (e.g. farm products, cheese, harvested cereals, hay, grass, soil, and the nests of birds and mammals) worldwide. This is partly due to its effect on germination and the nutritive value of grain (e.g [Citation1,Citation4].). The two acarid mites, A. siro and T. putrescentiae decreased germination form 20 to 70% for cereals and from 4 to 100% for vegetables [Citation5]. The effect of C. berlesei on some stored products (soybean, wheat, maize and fishmeal) on some chemicals change was studied by Gamal Eldin et al. [Citation6]. This mite decreased protein content, but fiber were increased.
Cheyletus malaccensis Oudemans was recorded in Egypt firstly associated with stored grain and seed pests [Citation7]. Also, this mite was recorded associated with stored products and house dust in the Philippines and in India [Citation8,Citation9].
The predatory mite, C. malaccensis commonly found in grain storage, is a potential natural enemy of stored-product pests [Citation10]. It was generalist predator fed on stored-product pests, such as A. siro, Aleuroglyphus ovatus (Troupeau), Caloglyphus redickorzevi (Zachvatkin), Caloglyphus rodriguezi Samsinak, Lepidoglyphus destructor (Schrank), Rhizoglyphus echinopus Fumouze and Robin, T. putrescentiae, and small insects such as eggs, young larvae or nymphs of moths and beetles [Citation1,Citation11,Citation12].
Cheyletus malaccensis has arrhenotokous reproduction; fertilized females produce offspring of both male and female, while the unfertilized females produce only males. The temperature has an impact on its survival, lifespan, fertility, and activity in biological control [Citation13,Citation14].
Initial studies on the mite life history were carried out by Zaher and Soleman [Citation15]. Various biological aspects were followed by several authors (e.g. [Citation10–12,Citation14,Citation16–19]).
Several researchers used various cheyletid mites in biological control strategies which resulted in significant reduction percentages of insect and mite pests [Citation13,Citation20].
The present study aims to elucidating the influence of three constant temperatures (22, 27 and 32°C) on biology and life table parameters of C. malaccensis feed on three astigmatid mites: A. siro, C. berlesei and T. putrescentiae.
Materials and methods
Prey mite culture
Three astigmatid mites Acarus siro (Linnaeus), Caloglyphus berlesei (Michael) and Tyrophagus putrescentiae (Schrank) cultures were established from samples pre-collected from stored grains at Qaha city in Qalyubia Governorate. The mites were mass-reared on wheat bran and dried yeast media with ratio of (4:1 wt) in plastic containers (11 X 8 × 3 cm) soaked with distilled water on another plastic containers. The rearing containers were covered by muslin to allow ventilation and prevent the mites escaping. The container’s bottoms were covered with a layer of mixture of cement and charcoal with ratio of (9: 1) and a depth of one cm. The cells were kept in a climatic chamber at 28°C, 80% RH., for six months before the study started at Qaha Agriculture Research Station, Qalyubia governorate, Plant Protection Research Institute, Agricultural Research Center, Egypt.
Predator mite culture
The predatory mite, Cheyletus malaccensis Oudemans was collected from wheat grain at Qaha city, Qalyubia governorate, Egypt. Cultures were maintained in the same containers filled with mixed of three acarid mite diets and wheat bran in the same condition. Mite identification was conducted according to the world references keys [Citation1].
Experimental unites
The experimental unites were black plastic cells each was (5 cm diameter and 2 mm in height), covered with glass cover slip. A circular hole was drilled in the center of each plate (1.5 cm in diameter and 1.5 mm in height). The study was conducted in an incubator at three constant temperatures (22, 27 and 32 ± 1°C) and 80 ± 5% RH. The three prey species: A. siro, C. berlesei and T. putrescentiae (immature stages) were used to feed C. malaccensis predator.
Biological aspects of C. malaccensis
To study the effect of prey species on biology and life table parameters of C. malaccensis, three groups of 40 newly deposited eggs were singly transferred with a fine brush to plastic plate and subjected to different temperatures. After egg hatching, 20 immature stages of the three prey diets were added daily to each C. malaccensis until reaching adulthood. The egg incubation period, immature stages, survival rates and fecundity of females were recorded daily. A male was introduced to each plastic plate for mating and removed from the plate after the deposition of the first egg. Every plastic plate was examined daily to record the number of eggs laid until the female died.
Life-table parameters
Life-table parameters as defined by Birch [Citation21] were calculated using a BASIC computer program [Citation22]. Sex ratio from each experiment was determined by visual observation and life tables were constructed from the data obtained for developmental time of immature stages and adult characteristics. Whereas: The net reproductive rate is the mean number of female offspring produced per female (R0) = Σ (lx × mx), where ‘mx’ is female progeny per female; ‘lx’ is the rate of females survival; the mean length of generation period, expressed in days (T) = Σ (x × lx × mx)/Σ(lx × mx); intrinsic rate of natural increase is a natural logarithm of the intrinsic rate of increase and indicates the number of times of population multiplication in a time unit (rm) = ln (R0)/T; means time of population to double (DT) = ln (2)/ rm and the finite rate of increase is the multiplication per female in unit time of a population with a stable age distribution (λ) = exp (rm).
Statistical analysis
The influence of prey species, developmental time, fecundity and duration of adult female reproductive stages were analyzed using One Way ANOVA and mean compared was conducted using Tukey’s HSD. Significance level was P > 0.05. Analysis was conducted using SAS program [Citation23]. The relationship between the rate of development (Y) and temperature (X) (at a specific range) can be represented by a straight line resembled with the linear equation (Y = a + bX °C), a (intercept), b (slope of temperature) and X is the tested temperature °C. The threshold temperature for development (t0) can be estimated using the equation, as – a/b (i.e. when Y = 0). The reciprocal of the slope (b) of the straight line (i.e. 1/b) is (K), which is the number of degree-days (DDUs) above (t0) required by an animal to complete its development as physiological time [Citation24,Citation25].
Results
The cheyletid predatory mite Cheyletus malaccensis Oudemans males passed through four stages (egg, larvae, protonymph, and adult) while female’s development passed through two protonymph stages before reaching adult.
Developmental time and longevity of C. malaccensis male reared on different prey species at different temperatures
The predatory mite C. malaccensis males completed its development on the astigmatid mites Acarus siro (Linnaeus), Caloglyphus berlesei (Michael) and Tyrophagus putrescentiae (Schrank). The results showed that all of the deposited eggs were hatched (100%). Developmental period (from egg to adult) of males was significantly affected by prey type and temperature (). Highly significant differences were obtained between the incubation periods of eggs of C. malaccensis males, the shortest period was 2.0 days at 32°C on A. siro, while the longest one was 5.30 days at 22°C on C. berlesei. The A. siro shortened the duration of male immature stages to 7.15 and 12.85 days at 27 and 22°C, but the longer immature stages was respectively 12.10 and 15.90 days when fed C. berlesei. The short life cycle of C. malaccensis male was 8.0 days on A. siro at 32°C, while the longest period was 21.20 days on C. berlesei at 22°C.
Table 1. Mean durations in days of Cheyletus malaccensis male reared on different prey diets at different temperatures and 80 ± 5% R.H.
The longevity of C. malaccensis male was affected by prey diets and temperature, the longest period was 22.10 days when fed on A. siro, while the shortest was 15.80 days when fed on C. berlesei ().
Effect of prey diets on C. malaccensis male
The durations of all developmental stages were longer on C. berlesei followed by T. putrescentiae, whereas the shorter periods were on A. siro. The finding in showed that the average immature stages of C. malaccensis were affected by prey diets; it was the shortest on A. siro (8.66 days) and the longest on C. berlesei (11.83 days). While longevity was longest when fed on A. siro (17.33 days) and shortest respectively on C. berlesei and T. putrescentiae (14.16 and 15.03 days). These results indicated that the acarid mite, A. siro is more favored prey for C. malaccensis than other diets.
Table 2. Effect of prey diets on durations of Cheyletus malaccensis male at different constant temperatures.
Effect of temperature on C. malaccensis male
Results presented in clarified that the optimum temperature for development of C. malaccensis was 32°C as the predator developed faster than other temperatures. At all three constant temperatures, there were highly significant variations between immature and adult stages. These results indicated that the temperature affected on duration of predator male than prey diets.
Developmental time and longevity of C. malaccensis females reared on different prey species at different temperatures
The results in indicated that developmental period (from egg to adult) of females was significantly affected by both prey types. Highly significant differences between different incubation periods of eggs of C. malaccensis; the shortest period was 2.20 days at 32°C on A. siro, while the longest one was extended to 5.55 days at 22°C when fed C. berlesei. The shortest protonymph and deutonymph periods of predatory mite female was 2.50 and 3.20 days at 32°C when fed A. siro, while, the longer period obtained was 7.65 and 7.85 days at 22°C, respectively when fed on C. berlesei. The short life span and generation period of C. malaccensis female was 11.60 and 13.40 days when prey on A. siro at 32°C, while the longest one was 29.50 and 36.50 days when prey on C. berlesei at 22°C, respectively.
Table 3. Mean durations in days of Cheyletus malaccensis females reared with different prey diets at different temperatures and 80 ± 5% R.H.
Significant differences were also found between adult female longevities and oviposition periods. The A. siro was more favored to the predatory mite followed by T. putrescentiae and C. berlesei. In addition, significant differences occurred between the three prey diets on female fecundity, the highest one was observed on A. siro was (196.5 eggs/ ♀) with daily rate of 10.15 eggs/ ♀/day at 27°C and the lowest rate was on C. berlesei was (69.10 eggs/ ♀) with daily rate of 4.72 eggs/ ♀/day at 22°C. Also the life span of C. malaccensis female was affected by temperature, the longest and shortest periods when fed on A. siro (68.05 and 33.80 days) at respectively 22 and 32°C ().
Effect of prey diets on C. malaccensis female
The obtained results indicated that biological aspects of a predator could be affected by the nutritional history of its prey, for example, changes in the nutritional components of prey can affect the predator’s growth and reproduction.
Similar results were obtained with males; the durations of all developmental stages were longer on C. berlesei followed by T. putrescentiae however the shorter periods was when fed on A. siro. The finding in showed that the average life cycle of C. malaccensis was affected by prey diets; it was the shortest on A. siro (16.93 days) and longer on C. berlesei (22.20 days). Despite female longevity was longeer on A. siro (31.56 days) and shorter (27.63 days) on C. berlesei. The oviposition period was longer for females fed on A. siro, with a daily and cumulative fecundity being two times higher than females fed on C. berlesei. These results indicated that the acarid mite, A. siro is favored prey for the C. malaccensis females more than other prey diets.
Table 4. Effect prey diets on durations of Cheyletus malaccensis females at different temperatures.
Effect of temperature on biological aspects of C. malaccensis female
The developmental time of C. malaccensis was significantly decreased as the temperature increase. Results obtained in explained that the optimum temperature for development was 32°C as the predator developed faster than other temperatures. The results also showed that developmental time, lifespan, oviposition period, and fecundity of C. malaccensis were all affected by temperature.
The mean total and daily fecundity of C. malaccensis fed on various diets are given in . The highest total egg production was at 27°C (139.2 eggs/♀) with a daily rate of (8.04 eggs/♀/day), followed by (121.9 eggs/♀/day) at 22°C and the lowest at 32°C (93.46 eggs/ ♀). These results indicated that the temperature affected the duration of predator females than prey diets.
Thermal requirements of C. malaccensis
The thermal factor had a negative relationship with duration of each stage as increasing temperature rapped development and shortened, duration from egg to adult. The lower threshold temperature (t0) for different stages of the predatory mite, C. malaccensis ranged between 6.32 and 15.15 form eggs to life cycle (). The accumulated day degrees (K) ranged between 37.37 and 301.06 DDUs from egg to adult. The values of coefficient of determination (the proportion of variation in the dependent variable that can be explained by the independent variable) R2 of C. malaccensis ranged between 0.92 and 1.0.
Table 5. Parameters of linear regression model for temperature-dependent developmental rates of immature stages of Cheyletus malaccensis females reared on different prey species.
Effect of prey types on life table parameters
The evaluation of C. malaccensis life table parameters at various temperatures are shown in . The shortest mean generation time (Tc) was observed when fed on A. siro was (18.6 days) at 32°C, while the longest (36.26 days) was recorded when fed on C. berlesei at 22°C. Whereas, the shortest time for population density doubling (DT) was 2.70 days at 27°C when fed on T. putrescentiae, while the longest period (7.70 days) was at 22°C when fed on C. berlesei. The maximum net reproductive rate (R0) (113.2 ♀/♀/generation) was occurred on A. siro at 27°C recorded, while the lowest value (26.94 ♀/♀/generation) when fed on C. berlesei at 22°C.
Table 6. Life-table parameters of Cheyletus malaccensis reared on different prey species and different temperatures.
The maximum intrinsic rate of natural increase (rm) when the difference between birth rate and death rate was obtained at 27°C when fed on T. putrescentiae was (0.256 individuals/♀/day) followed by (0.233 individuals/♀/day) when fed on A. siro at 32°C, whereas the lowest (0.090 individuals/♀/day) was recorded when fed on C. berlesei at 22°C. The finite rate of increase (λ) ranged from 1.09 offspring/individual/day at 22°C when fed on C. berlesei to 1.29 offspring/individual/day at 27°C when fed on T. putrescentiae. Gross reproduction rate (GRR) recorded the highest value (117.4 eggs/ female/generation) when fed on A. siro at 32°C, the lowest value (43.82 eggs/ female/generation) when fed on C. berlesei at 22°C .
The population of C. malaccensis is reduced by half (50% mortality) in 26.0 days at 27°C when fed on T. putrescentiae and 49.0 days at 22°C when fed on A. siro. Daily age-specific survival rate ranged from 60.0 to 80.0%. The sex ratio ranged from 0.60 to 0.72 female/total not affected by temperature and prey diet ().
Age specific survival rate of C. malaccensis reared on different prey and temperature
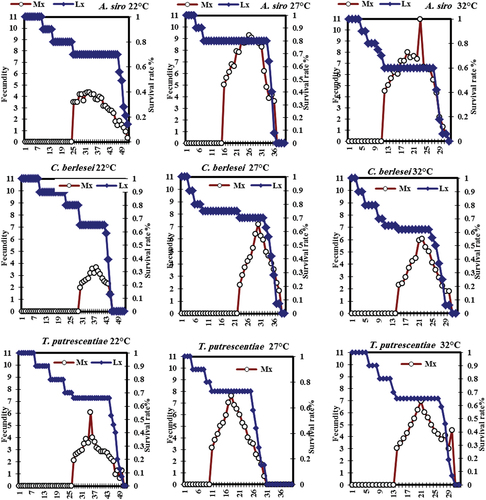
Age specific survival rate (lx) and fecundity (mx) curves for C. malaccensis are shown in . The daily age-specific survival rate was highest at 27°C and decreased as the temperature increased on the three prey species, all of the deposited eggs hatched (100%). The maximum number of eggs produced (day 23: 10.99 egg/♀/day) when fed on A. siro at 32°C, the lowest (day 38: 3.66 egg/♀/day) when fed on C. berlesei at 22°C. The highest survival rate of females was 80% when fed on A. siro at 27°C, while lowest value was 0.60% when fed on C. berlesei at 22°C.
Figure 1. Age-stage specific survival rate (Lx) and fecundity (Mx) curves of Cheyletus malaccensis at different temperatures.
Discussion
The predatory mite C. malaccensis abled to survive and reproduce on A. siro, T. putrescentiae and C. berlesei. The findings agree with that of [Citation11] who found T. putrescentiae immatures are more preferable food for C. malaccensis. On the contrary, Aleuroglyphus ovatus (Troupeau) and Caloglyphus redickorzev Zach. are more preferable than T. putrescentiae [Citation16]. In the current study, C. malaccensis feed well on A. siro than other prey species. The optimum temperature for its development was 32°C as developed faster than other on other temperatures. Whereas, the survival rate was highest at 27°C then decreased as the temperature increased on all prey species. Similar results obtained by [Citation19] indicated the values of age-specific fecundity (high to low) were 28°C > 24°C > 30°C > 32°C > 22°C. Previously published works on C. malaccensis [Citation12] indicated a significant difference in the duration of the egg–adult period which was shorter when feeding on T.putrescentiae than Caloglyphus rodrigeuzi Samsinak. The shortest life cycle averaged 16.3 days at 24°C and 85%RH, while the longest was 18.6 days at 24°C and 65% RH. The female fecundity, oviposition period and daily fecundity were respectively 493.0, 46.2 d, and 10.3 at 85% RH [Citation10]. The shortest incubation period was at 32°C and the longest was at 22°C in both females and males. The life cycle ranged from 11.10 to 27.50 days for females and 8.80 to 22.71 days for males at 22°C [Citation19].
In this study the thermal factor had a negative relationship with duration of each stage as increasing temperature rapped development and shortened egg-adult duration. The present results are almost agree with the findings of [Citation17] who indicated that lower and upper developmental thresholds ranged between 11.6–12.0 and 37.4–37.8°C, respectively. The thermal constant ranged between 238.1 and 312.5 degree-days. Based on the data of the total pre-imaginal period, immatures survival peaked at 25°C and R2 ranged between 0.94 to 0.99.
The relationship between temperature and rate of development in insects and mites is typically estimated as linear, whereas it is really curved [Citation26].
According to effect of temperature and prey diet on life table parameters of C. malaccensis in the current study, the highest (rm) and finite rate of increase (λ) was obtained at 27°C when fed on T. putrescentiae. These results are in coincidence with that of [Citation13] who found that rising temperature faster the natural increase of C. malaccensis feed acarid mites and helped to double its numbers in a shorter time. The intrinsic rate of population increase (rm) of C. malaccensis was strongly affected by temperature increase from 17.5°C to 32.5°C, while the (rm) was 0.213 individuals/♀/day for C. malaccensis at 32.5°C [Citation14]. The prey species obviously affected rm value of C. malaccensis (0.059 individuals/♀/day) when feed T. putrescentiae. The highest finite rate of increase (λ) was (1.062) when C. malaccensis individuals feed T. putrescentiae [Citation12]. The mean generation time was 30.3 and the finite rate of increase was 1.09 when feed T. putrescentiae and A. siro [Citation18]. The highest net reproductive rate (R0 = 290.25) and highest fecundity (544.52) occurred at 28°C, as temperature significantly affected the intrinsic rate of increase (rm) and fecundity [Citation19]. The sex ratio of C. malaccensis offspring and mortality were not significantly influenced by temperature [Citation17].
Conclusion
From the obtained results it can be concluded that 27°C is satisfactory temperature than 22 and 32°C for population increased of C. malaccensis when reared on A. siro. These findings can be used to forecast population dynamics, as well as to guide mass rearing and use of C. malaccensis predator to reduce stored-product pests. It can also be used in Egypt as a biological control agent to reduce the amount of acarid mite pests that infest stored commodities.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Hughes AM. The mites of stored food and houses. Tech Bull Ministry Agric Fish Food. 1976;9:1–400.
- Taha HA.Morphological and biological studies on some mites associated with stored products.Ph.D Thesis Agric Zool Nematol Depart Fac Agric Al-Azher Uni.1985:159.
- Athanassiou CG, Kavallieratos NG, and Palyvos NE, et al. Spatio-temporal distribution of insects and mites in horizontally-stored wheat. J Econ Entomol. 2005;98(3):1058–1069. doi:10.1603/0022-0493-98.3.1058
- Van Hage-Hamsten M, and Johansson SG. Storage mites. Exp Appl Acarol. 1992;l(16):117–128. doi:10.1007/BF01201495
- Zdarkova E.The effect of mites on germination of seed.Ochrana Rostlin- Plant Protection Science.1996;32:175–179.
- El-Din HM G, Metwally AM, AbdAllah A, and El-Bltagy, HM. A study on some stored products infected by Caloglyphus berlesei. Egypt J Nutr Feeds. 2017;20:77–84. doi:10.21608/EJNF.2017.103927
- Wafa AK, Zaher MA, El-Kifl AH, et al. Survey of stored grain and seed mites (Acarina). Bull Soc Entomol d’Egypte. 1966;50:225–232.
- Corpuz-Raros LA, Sabio G, and Velasco-Soriano M. Mites associated with stored products, poultry houses and house dust in the Philippines. Philipp Entomol. 1988;7:311–321.
- Putatunda B. Mites (Acarina) associated with stored food products in Himachal Pradesh, India: a taxonomic study. J Entomol Res. 2005;29:79–82.
- Liu L, Cao Y, and He P, et al. A preliminary study of growth and development of Cheyletus malaccensis (Oudemans) under different humidity conditions. J Henan Univ Technol Nat Sci Educ. 2018;5:102–107. doi:10.5073/jka.2018.463.117
- Yousef AA, Zaher MA, Kandil MM. Effect of prey and temperature on the development and biology of Cheyletus malaccensis Oudemans (Acari: Cheyletidae). Z Ang Entomol. 1982;93:39–42.
- Al-Shammery KA. Influence Cheyletus malaccensis (Acari: Cheyletidae) in Hail, Saudi Arabia. Life Sci. 2014;11(5):260–266. http://www.lifesciencesite.com
- Pekár S, and Hubert J. Assessing biological control of Acarus siro by Cheyletus malaccensis under laboratory conditions: effect of temperatures and prey density. J Stored Prod Res. 2008;44:335–340. doi:10.1016/j.jspr.2008.02.011
- Palyvos NE, and Emmanouel NG. Reproduction, survival, and life table parameters of the predatory mite Cheyletus malaccensis (Acari: Cheyletidae) at various constant temperatures. Exp Appl Acarol. 2011;54:139–150. doi:10.1007/s10493-011-9427-7
- Zaher MA, Soliman ZR. Life-history of the predatory mite Cheyletus malaccensis Oudemans. Bull Soc Entomol d’Egypte. 1971;55:49–53.
- Cebolla R, Pekár S, and Hubert J. Prey range of the predatory mite Cheyletus malaccensis (Acari: cheyletidae) and its efficacy in the control of seven stored-product pests. Biol Control. 2009;50:1–6. doi:10.1016/j.biocontrol.2009.03.008
- Palyvos NE, and Emmanouel NG.Temperature-dependent development of the predatory mite Cheyletus malaccensis (Acari: cheyletidae).Exp Appl Acarol.2009;47:147–158. doi:10.1007/s10493-008-9200-8
- Granich J, Horn T, Körbes J, Toldi, M, Da Silva, G, and Ferla, N . Development of Cheyletus malaccensis (Acari: cheyletidae) feeding on mite species found in commercial poultry systems: megniniag inglymura (Acari: analgidae) and Tyrophagus putrescentiae (Acari: acaridae). Syst Appl Acarol.2016;21:1604–1613. doi:10.11158/saa.21.12.2
- Sun W, Cui M, Xia L, Yu, Q, Cao, Y, and Wu, Y . Age-stage, two-sex life table’s of the predatory mite Cheyletus malaccensis Oudemans at different temperatures. Insects. 2020;11(181):11–13. doi:10.3390/insects11030181
- Mesbah AE. Efficiency of two predatory mites Amblyseius swirskii and Cheletogenes ornatus early release in controlling the two spotted spider mite, Tetranychus urticae Koch on Soybean plants in Sharkia governorate. J Plant Prot Path Mansoura Univ 2016;7(12):837–843. doi:10.21608/JPPP.2016.52467
- Birch LC. The intrinsic rate of natural increase of an insect population. J Anim Ecol. 1948;17:15–26.
- Abou-Setta MM, Sorrell RW, Childers CC. Life 48: a BASIC computer program to calculate life table parameters of an insect or mite species. Flor Entomol. 1986;69:690–697.
- Anonymous. SAS statistics and graphics guide, Release 9.1. Cary, North Carolina, 275l3, USA; SAS Institute; 2003.
- Elhalawany AS, and Abdel-Wahed N. Effect of temperature and host plant on developmental times and life table parameters of Tetranychus urticae Koch on Persimmon trees (Acari: tetranychidae). Egypt J Agric Res. 2013;91(2):595–607. doi:10.21608/EJAR.2013.163532
- Abou-Setta MM. Nutritional ecology bridges the gap between mites’ biological and ecological research results under Mediterranean environment. Acarines. 2020;14:45–52.
- Sharpe J, De Michele D. Reaction kinetics of poikilotherm development. J Theor Biol. 1977;64:649–670.