
ABSTRACT
Island ecosystems are highly threatened by invasive rats and mice. Currently, the only effective technology for eradicating rodents from islands is toxicants. Though effective, they are expensive and have high failure rates. They are not species-specific and are potentially dangerous to humans. Gene drive technology is one alternative to toxicants for rodent eradication. Gene drive methods of rodent eradication offer an alternative to killing that has the potential to be more species-specific, more humane, and more biologically safe for use around humans. Technologies in development aim to apply either natural meiotic drive or clustered regularly interspersed short palindromic repeats to influence offspring development so that all offspring are phenotypically male, eventually creating a population that is not reproductively viable. Implementing this technology would involve releasing laboratory-developed engineered mice into wild populations. Some areas for further research include assessing the ecological effects of releasing engineered mice, the potential risks for the accidental or deliberate release of genetically modified organisms into mainland mouse populations, and the social, ethical, and regulatory acceptability of the technology.
1. Introduction
Invasive species are responsible for over 50% of animal extinctions worldwide (Clavero and García-Berthou Citation2005; Doherty et al. Citation2016). Island ecosystems are at a greater risk from invasive species than mainland ecosystems (Sax, Gaines, and Brown Citation2002). While islands compose only 5% of Earth’s landmass, they disproportionately contain over 20% of terrestrial animal species (Howald et al. Citation2007). One of the greatest threats to island plant and animal species today are invasive rats (Rattus rattus, R. norvegicus, R. exulans) and mice (Mus musculus) (Campbell et al. Citation2015). Invasive rodents have been implicated in many island extinctions – they are present on over 80% of islands worldwide due to human introduction, and are associated with the extinction of over 60 vertebrate species alone (Towns, Atkinson, and Daugherty Citation2006; Howald et al. Citation2007).
The significant biodiversity threat posed by rats and mice makes their removal a critical conservation goal, but both species are difficult to eradicate from many localities using current methods, primarily toxicants (Howald et al. Citation2007; Campbell et al. Citation2015). We and other researchers are studying gene drive technologies as another method of controlling or eliminating invasive rodent populations from islands: genetic biocontrol of rodents. Our goals in this paper are to summarize the threats rats and mice pose to biodiversity on islands, to compare toxicants and the genetic biocontrol of rodents for eradicating invasive rodent populations, and to contribute to discussions about ecological risk and social acceptability.
2. Threats posed by invasive rats and mice to biodiversity on islands
Rodents travel commensally with humans and can adapt to a broad range of natural habitats and environments (Singleton and Krebs Citation2007). Both rats and mice are omnivorous and opportunistic feeders, making them particularly damaging to native island species, especially those that evolved without mammalian predators (Towns, Atkinson, and Daugherty Citation2006; Howald et al. Citation2007; Mackay, Russell, and Murphy Citation2007).
On islands without humans, mouse diets typically range from grains and grasses to invertebrates, depending on availability. On islands, food availability is often seasonal – a study of the feeding habits of invasive mice on a subantarctic island showed their diet consisted of high percentages of invertebrates when grasses and seeds were not available (Le Roux et al. Citation2002). Population declines and extirpations of both plants and invertebrates increase on islands with invasive mice (Crafford and Scholtz Citation1987; Angel, Wanless, and Cooper Citation2009). Mice will also feed on vertebrates, most notably the eggs and nestlings of nesting birds. The starkest example is on Gough Island – lacking sufficient plant and invertebrate food, invasive mice primarily consume bird eggs and chicks (Parkes Citation2008). Two seabird species, the Tristan albatross and Atlantic petrel, and a native terrestrial bird, the Gough bunting, are listed by the International Union for the Conservation of Nature as endangered or critically endangered, and invasive mice have been implicated as the cause of the declining populations (Parkes Citation2008).
Invasive mice also induce secondary ecological effects, including decreasing endemic species reproduction. While the adult animals or plants may not be affected by mice, eggs, young, or seeds may be consumed in such abundance that the species is effectively unable to reproduce (Le Roux et al. Citation2002; Parkes Citation2008). The presence of mice may also affect the food web through hyperpredation or increasing the threat to native species by attracting non-indigenous predators (Courchamp, Langlais, and Sugihara Citation2000). For example, on the Farallon Islands (CA, USA), the presence of mice creates an abundant food source for migrating owls and is believed to cause some owls overwinter on the islands instead of continuing migrating as they would do otherwise (South East Farallon Islands EIS Citation2013). Without sufficient food resources over the winter, the mouse population on the Farallon Islands crashes, and the owls consume ashy storm petrels, a threatened seabird that breeds on the island (South East Farallon Islands EIS Citation2013).
Rats’ effects on islands are similar, if not greater, compared to mice (Harper and Bunbury Citation2015). The damages rats can cause are well documented in New Zealand, where they have been present for hundreds of years. On a single island in New Zealand, Little Barrier Island, rats were documented to threaten a number of native plants, seabirds, invertebrates, and reptiles before being eradicated (Towns, Atkinson, and Daugherty Citation2006). On the Lord Howe Island Group, off the coast of Sydney, Australia, rats have brought on the extinction of many endemic species, including 5 species of birds, 2 species of plants, and 13 species of invertebrates (Wilkinson and Priddel Citation2011). Some species of invertebrates and birds now only exist in small populations of islands where rats are not present, and the population of many species of plants are in decline where rats are present (Wilkinson and Priddel Citation2011). Due to their size, rats are more threatening than mice to vertebrates and able to prey on adult birds, eggs, and chicks, as well as on other mammals (Towns, Atkinson, and Daugherty Citation2006; Angel, Wanless, and Cooper Citation2009; Harper and Bunbury Citation2015).
While the focus of this assessment is on the conservation threats posed by invasive rodents, it should be noted that rodents also represent threats to health and livelihoods on islands with human populations. Though these threats are not conservation-oriented, it is worth mentioning the potential future benefits of improved rodent pest management for humans living in areas impacted by rodents, as pressing human health and agriculture concerns could help drive the development of the technology. Rodents can consume vast quantities of grain and rice crops, causing loss of both a food source and a source of income. Rice farmers in Asia experience a 20–30% loss of crops yearly on average, with some farmers losing 50–100% of their crops to rodent pests (Singleton Citation2003). The potential damage from rodents also prevents many farmers from planting more rice crops per year. Taken together, these losses amount to enough rice to have fed 180 million people for a year (Singleton Citation2003). Similar effects are seen in Australia. One study found that damage from mice to cereal, rice, soybean, and maize crops in one month caused losses ranging from 14% to 66% (Brown and Singleton Citation2000). Additionally, mouse irruptions lead to damage to electrical equipment, animal housing, and human dwellings and food supplies (Brown and Singleton Citation2000). Rodent pests in agricultural areas also increase the potential for disease – over 60 zoonotic diseases are carried by rodents, some of which can cause death (Singleton Citation2003). Considering the threats to human health and agriculture, the potential benefits of improved rodent pest management to humans is significant.
3. Methods of rodent eradication
Currently, toxicants are the only effective technology for eradicating rodents from islands greater than 5 ha (Campbell et al. Citation2015). Second-generation anticoagulants, also known as ‘superwarfarins,’ are the toxicant of choice (Ishizuka et al. Citation2008; Campbell et al. Citation2015). Developed after rodents displayed resistance to the similar but less potent toxicant warfarin, these anticoagulants cause death through internal hemorrhaging (Ishizuka et al. Citation2008). Brodifacoum, the most widely used superwarfarin for rodent eradications, is palatable to rodents and is easily dispensed through bait stations or aerial broadcasting (Howald et al. Citation2007). It is lethal with the consumption of only a few grams and has a delayed effect, discouraging invasive rodents from associating the toxicant bait with illness and death (Howald et al. Citation2007).
Genetic pest management offers a potentially attractive alternative to using toxicants for rodent eradication, especially because the methods being proposed are non-lethal. The most promising of these new technologies are gene drive systems. Gene drives are selfish genetic elements that can replicate their genetic sequence and insert it into the genetic sequence of the gametes of a sexually reproducing species. By manipulating a gene drive and adding in a desired trait, humans could theoretically spread the desired trait and, in the case of invasive rodents, suppress wild populations. Most genes have normal Mendelian inheritance, or a 50% chance for each of two alleles for a gene being passed on to the next generation. Gene drive systems have super-Mendelian inheritance, which is broadly defined as an allele having greater than the 50% inheritance pattern expected. Accordingly, they have the potential to spread quickly through a population (Lyttle Citation1991).
Current research on genetic biocontrol of rodents is confined to mice due to the relative ease in manipulating the mouse genome in comparison to rats. Mice have a naturally occurring gene drive on Chromosome 17 called the t-haplotype (Willison and Lyon Citation2000), which is being developed for the genetic biocontrol of mice. Male mice heterozygous for the t-haplotype pass it on to greater than half of their offspring, with some variants of the t-haplotype having transmission rates of over 90% (Bauer et al. Citation2005). Sperm containing the t-haplotype are more successful than sperm without the t-haplotype, leading to an increased proportion of eggs being fertilized by sperm with the t-haplotype (Bauer et al. Citation2005). Mice homozygous for the t-haplotype usually die before birth from accumulated mutations in the t-haplotype. The tw2 variant of the t-haplotype is being used in developing genetic biocontrol for mice since males homozygous for the t-haplotype are sterile instead of dying before birth. Female mice with the tw2 variant have normal fertility (Lyon Citation2003).
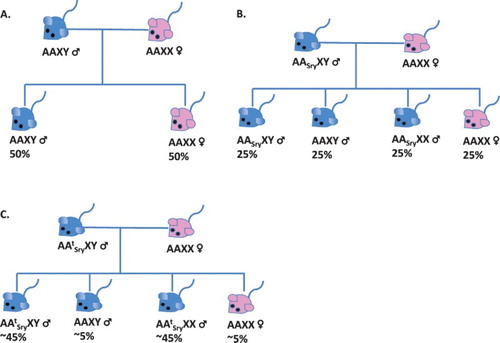
Gene editing tools are being used at Texas A&M University to create ‘daughterless’ mice – in other words, mice that are unable to bear female offspring – by inserting the Sry gene sequence into the t-haplotype. The Sry gene is normally located on the Y-chromosome and controls the development of male characteristics. Because the t-haplotype has meiotic drive and is located on an autosome, in order to spread the Sry gene to both XX and XY offspring, the Sry gene has to be copied and inserted into the t-haplotype. This ensures that all offspring receiving the t-allele with the Sry gene, regardless of chromosomal sex, would be phenotypically male (; Piaggio et al. Citation2017). Targeting a construct containing Sry to the t-haplotype should result in nearly all offspring inheriting the Sry gene. Releasing male genetically engineered (GE) mice with the Sry/t-haplotype into a population of wild mice, where they could breed with wild females, could be effective in spreading the Sry/t-haplotype through a population and increasing the relative proportion of male mice in a population.
Figure 1. Sry gene drive: to skew sex ratios in naturally breeding populations, the male determining gene (Sry), normally found on chromosome (Chr) Y, can be inserted into a naturally occurring gene drive element on Chr 17 called the t-complex. The t-complex is passed down to greater than 90% of the offspring through the paternal side. XX and XY indicate the sex chromosomes and A indicates any of the 22 autosomes. ASry is the Sry gene inserted into an autosome and AtSry is Sry inserted into the t complex. (A) In normal breeding scenarios, the Sry gene is only located on Chr Y and therefore only mice inheriting Chr Y are male, resulting in approximately 50% of the offspring are XY (male) and 50% are XX (female). (B) In a breeding scenario where the Sry gene has been added to any autosome, approximately 75% of the offspring will be male and 25% will be female. (C) In breeding scenarios where the male carries the Sry gene within the t-complex, over 90% of offspring will inherit the t-complex containing autosome. It is predicted that fewer than 10% of the offspring will be XX (female), with the remaining being phenotypically male, including either XY (male) or XX (sterile male).
Currently, the best gene drive alternative to the Sry/t-haplotype construct would be using a clustered regularly interspersed short palindromic repeats (CRISPR)/Cas9 gene drive system. Using CRISPR as a gene drive mechanism could enable the genetic pest management strategy to be applied to more species, as it does not rely on a native gene drive mechanism. The Sry/t-haplotype system is currently being explored because both components naturally exist in mice and there is a better understanding and ability to manipulate the mouse genome.
3.1. Drawbacks of eradication technologies
There are downsides to both types of technologies being examined for rodent eradications (). Many of the downsides are related to the removal of rodents from an ecosystem, regardless of the technology used to do so. Biomathematical modeling has been helpful in addressing these types of ecological questions. Whether using current methods or gene drive, the ecological ‘hole’ that is created when an entire species is removed has been addressed by modeling for invasive rodents (Zavaleta, Hobbs, and Mooney Citation2001). The presence of an invasive species, especially species that are food sources for predators, can attract other species in search of food, as seen on the Channel Islands and the Farallon Islands (Collins, Latta, and Roemer Citation2009; South East Farallon Islands EIS Citation2013). If the invasive food source is removed too quickly, the predator may turn to consuming endemic species rather than leave the island (Courchamp, Woodroffe, and Roemer Citation2003; Collins, Latta, and Roemer Citation2009). In the case of invasive rodents, if both mice and rats are present, they can have additional interactions that make removal difficult. When trying to suppress one or both populations, the overall reduction in individuals can cause a rapid population growth of one or both species (Caut et al. Citation2007). Fortunately, the use of modeling can help in planning species removal timing in order to more closely restore the food web to pre-invasive interactions (Courchamp, Woodroffe, and Roemer Citation2003; Caut et al. Citation2007; Caut, Angulo, and Courchamp Citation2009; Collins, Latta, and Roemer Citation2009).
Table 1. Comparing toxicants and gene drives.
There are also downsides specific to the technology being used. Brodifacoum is a biologically efficacious method for eradicating invasive rodents, but eradication campaigns using toxicants can still fail. Though reported instances of toxicant resistance are low, failed eradication campaigns increase the risk for mice and rats to develop resistance to currently used anticoagulants (Ishizuka et al. Citation2008). Mutations in the Vkorc1 protein can lead to resistance to anticoagulants and many eradications using toxicants now test for mutations in Vkorc1 before proceeding (Didion, Threadgill, and Pardo-Manuel de Villena Citation2012). Hybridization with native, closely related species resistant to Brodifacoum has also been observed in mice (Song et al. Citation2011), suggesting that there are multiple avenues for rodents to develop resistance to Brodifacoum. Resistance to Brodifacoum and other toxicants can make future eradication campaigns more difficult and can also cause problems in controlling rodents in urban areas if the resistant rodents spread. Although rat eradication campaigns using Brodifacoum and similar toxicants have been highly successful and have only a 5–10% failure rate, the same methods have a failure rate close to 40% when applied in the context of mouse eradications (Howald et al. Citation2007; Angel, Wanless, and Cooper Citation2009). Brodifacoum was developed after rodent resistance to Warfarin was widespread (Ishizuka et al. Citation2008), and if resistance to Brodifacoum reaches similar levels, a new toxicant would also have to be developed.
Explaining these higher failure rates for mice is difficult, but mice have smaller home ranges and different foraging behavior than rats, which may decrease mouse encounters with toxicants (Howald et al. Citation2007). Rodent eradications using toxicants typically involve a single release of toxicants across an island, which is then monitored in the following years for the presence of rodents (Howald et al. Citation2007). Failure of an eradication is often not detected until multiple years have passed. The toxicants do not persist long in the environment before degrading, and by the time failure is detected, a new campaign has to be developed (Howald et al. Citation2007).
Rodents can also be resistant to gene drives, both behaviorally and because the individuals carrying the gene drives often have lower fitness than the non-drive carriers. The t-haplotype is naturally occurring in Mus musculus, but only at low rates – only 6–25% of the wild mice have the t-haplotype (Carroll et al. Citation2004). Most strains have a recessive lethal allele associated with being homozygous for the t-haplotype. Females who carry the t-haplotype also tend to avoid males who are also heterozygous for the t-haplotype (Lindholm et al. Citation2013). It is not possible to contain the t-haplotype itself. However, it is likely that over time the autosomal Sry gene would be naturally selected against and only the natural t-haplotype without the Sry gene would remain in wild populations. Selection also naturally occurs against the t-haplotype, which should limit its drive (Carroll et al. Citation2004). Additionally, using an autosomal Sry involves inserting a large piece of DNA, which could decrease the fitness of the mouse, potentially inhibiting its ability to mate and thereby diminishing the effectiveness of the eradication campaign. It is possible that the translocated Sry gene will impair an important function of a gene or cause the mouse to not be able to compete as effectively as the native mice on the island. For these reasons, for the t-haplotype to be used as an effective drive it would likely need multiple releases (Backus and Gross Citation2016).
There is also a concern for secondary effects of these technologies. Second-generation anticoagulants have a greater impact on non-target species than warfarin, either through direct consumption or through consumption of toxicant-laced rodents or rodent bodies. While the biggest non-target effects are on birds, other native vertebrates are also impacted (Ishizuka et al. Citation2008). On islands with humans, pets, and livestock, toxicants pose an additional threat (Campbell et al. Citation2015). Not only do eradications on islands with humans require additional planning, adding to the lengthy years-long planning process, the financial cost also increases when incorporating protections for human populations (Campbell et al. Citation2015; Holmes et al. Citation2015). Eradication campaigns must also be timed to the cyclical patterns of rodent populations, particularly for islands in higher latitudes where food and water availability fluctuate (Mackay, Russell, and Murphy Citation2007). Eradication campaigns cost millions of dollars, and require a large number of people to implement them successfully (Howald et al. Citation2007; Holmes et al. Citation2015). If a campaign fails, financial and ecological costs are accrued again.
Gene drive technologies also have secondary effects, though many of them are still hypothetical while the technology is being developed in the lab. The primary secondary effect being examined is the temporary rodent population increase that happens when releasing the GE rodents. The use of a gene drive mechanism to eradicate rodent populations would initially involve increasing the overall population with the introduction of GE rodents. During this transitory period, ecological interactions can be intensified, potentially leading to permanent ecological consequences, such as further impacts to the ecosystem, potentially including some of the endemic species that are being protected (David et al. Citation2013; Esvelt et al. Citation2014; Backus and Gross Citation2016). These damages could potentially increase the overall cost of the eradication as well.
While neither technology being examined here is free from biological concerns, there are also social issues to consider. Eradicating invasive species can be a socially contentious issue that leads to a lack of support from local communities and may contribute to the failure of eradication campaigns (Howald et al. Citation2007; Campbell et al. Citation2015). Rather than causing a quick and painless death, anticoagulants like Brodifacoum kill over a period of days, and related animal welfare concerns can lead to public resistance (Howald et al. Citation2007; Campbell et al. Citation2015). The lack of social support, the risks toxicants pose to non-target species including humans and livestock, and the high financial and time costs provides strong impetus for consideration of newer, safer, and more humane technologies for removing invasive rodents from islands. The development of these new technologies also calls for an examination and integration of social opinion, which we discuss later on.
3.2. Containment and reversal of gene drive systems
Because gene drive systems, unlike toxicants, can spread on their own, biosecurity is a major concern (Esvelt et al. Citation2014). Containment and biosecurity must be addressed at various levels, from laboratory to wild settings. Many safe laboratory practices, such as secondary containment and security protocols, already exist for rodents, as mice and rats are frequently used for biomedical research. The need for proper containment of gene drive systems and proposed methods has also been addressed in laboratory settings by Akbari et al. (Citation2015) and Min, Smidler, and Esvelt (Forthcoming). Nevertheless, the deliberate release of gene drive biotechnologies subverts the very the concept of ‘containment,’ given that the technology is designed to spread (Min, Smidler, and Esvelt, Forthcoming). While the waters surrounding island ecosystems offer a geographic hurdle, rodents have a long history of uninvited travel on human sea vessels, and rats, in particular, are capable of swimming up to 1 km and for 3 days straight (Russell, Towns, and Clout Citation2008; Harper and Bunbury Citation2015). Additionally, both mice and rats are difficult to catch and can fit through holes smaller than 2.5 cm in diameter, which makes preventing escape (or reinvasion) difficult (Centers for Disease Control and Prevention Citation2016).
The 2016 National Academies of Sciences, Engineering, and Medicine (NASEM) report on gene drives suggests a detailed phased-testing pathway from conception of an idea to application in the field and post-application monitoring that incorporates both containment strategies as well as feedback from scientists, stakeholders, community members, and the general public (NASEM Citation2016). Modeled on the phased approach that the World Health Organization proposed for testing genetically modified mosquitoes (WHO Citation2014), this pathway would involve thoughtful evaluation at each step of the process, including checkpoints that determine whether research should move to the next phase. We recommend adopting this testing pathway (or something similar) for further development of a gene drive mouse because the proposed pathway is thorough in addressing containment issues and biological risk, transparent to the public and welcoming to their input (an issue that we discuss in more detail in Section 5), and goal-oriented, with a clear end-point.
Many questions of biological containment are already being considered in the development of a gene drive mouse. An ideal gene drive system would be able to spread through and eradicate a target island population but not spread through any mainland populations where they might unintentionally escape. One way to achieve this is to specifically design a threshold system where the gene of interest would only spread through population when the gene is above some sufficiently high threshold (Curtis Citation1968; Davis, Bax, and Grewe Citation2001; Akbari et al. Citation2013). Alternatively, a self-limiting gene drive like the Sry/t-haplotype mouse could behave like a threshold system if gene drive organisms are repeatedly released into the population above some minimum release rate (see Sections 3.1 and 4). In this situation, the Sry/t-haplotype construct would be forced to spread and eradicate the wild population even though the gene construct would be expected to be lost from the population when the release rate is not maintained (Backus and Gross Citation2016). If any mice carrying this Sry/t-haplotype construct were to escape the island, they would not be maintained at a high enough frequency to eradicate the mainland population (Backus and Gross Citation2016).
If CRISPR is used as an artificial gene drive, there are a number of ways to ensure that it would not spread to non-target species or to mainland rodent populations. It could be targeted to a very species-specific locus in the genome to ensure that it would not easily move laterally between species. Synthetic site targeting, or first inserting a non-coding sequence into a wild, invasive population and then targeting that sequence for Sry insertion would be possible using CRISPR, though not using the t-haplotype (Min, Smidler, and Esvelt, Forthcoming). This would help ensure that only the targeted population containing the specific non-coding sequence could receive the gene drive. A split drive system could also be used, where the construct of interest is broken up into multiple parts and inserted separately, which allows for monitoring at every stage and lessens the chance of accidental escape (Min, Smidler, and Esvelt, Forthcoming).
Regardless of the type of gene drive mechanism, an inducible system could be used to turn Sry function on or off, such as with the tet regulatory system controlled by doxycycline. The system can be designed such that when the mice are provided doxycycline in their water, the desired gene is turned either on or off through transcription regulation. This could be built into the Sry construct where the presence of doxycycline would turn on the Sry gene and lead to male development. If a mouse were to escape the island, then the Sry gene would be inactive. If that mouse were to reproduce, it would spread the inactive gene, and all of that mouse’s offspring would develop according to their inherited sex chromosomes. The challenge would be to have doxycycline available to the mice on the island of interest, as eradication could prove unsuccessful if not all mice consume it.
A way of reversing the production of male-only offspring if either the t-haplotype or CRISPR were used would be to introduce wild type males and females resistant to the drive system back into the population. Because success of the t-haplotype system is dependent on being above a certain frequency in the population, releasing wild type mice back into the system and not releasing any more GE mice should be enough to re-establish an invasive mouse colony. Though this has not been tested directly, there is ample evidence that female mice prefer mice without the t-haplotype (Carroll et al. Citation2004; Lindholm et al. Citation2013; Manser, König, and Lindholm Citation2015), and if there are enough male mice without the t-haplotype reintroduced, the wild females are likely to preferentially mate with those males. If using CRISPR as a gene drive system, the most effective way to reverse the system would be targeting an innocuous sequence found only in the engineered mice to override the current system, instead of targeting the Sry gene directly. Targeting the Sry gene would target both the autosomal Sry as well the Sry gene present on the Y-chromosome, which would also disrupt normal male-female ratios. While the resulting mice would still have autosomal Sry genes, their function would be disrupted and they would have normal male to female ratios in their offspring.
Regardless of the method used, proper monitoring and enforcing physical containment can help mitigate the risk of accidental escape and spread to the mainland population. Questions of monitoring and containment are addressed using the NASEM (Citation2016) phased-testing pathway. Included in the phased-testing pathway would be a field trial using just the gene drive mechanism with no linked gene. A trial of this nature is important to ascertain the effectiveness of the gene drive method, as well as to test out components of biosecurity and how quickly a gene would spread. We address additional research methods in the following section to predict how different drive systems will work and the risks they may pose to non-target populations.
4. Addressing ecological influence and risk
Addressing ecological risks of using genetic biocontrol of rodents is difficult, as the technology is still in development and not easily studied. However, the authors of this study believe that performing an ecological risk assessment is imperative to safe implementation in the field, a view that is in line with the recent recommendations of the NASEM (Citation2016) gene drive report. The NASEM report recommends both that risk assessments be performed prior to the implementation of gene drive technologies, and that field trials be conducted for ecological applications to ensure that there is enough information for a risk assessment. Given that there has not yet been an ecological risk assessment performed for a field trial of gene drive technologies, it is especially important that one be performed before (and after if necessary) a field trial of genetic biocontrol of rodents.
An important component of defining risk is acknowledging uncertainty. The NASEM (Citation2016) report on gene drives defines two types of uncertainty: linguistic, or normative, uncertainty, and epistemic, or fact-based, uncertainty. There are high levels of both types of uncertainty when examining the potential use of genetic biocontrol of rodents. Linguistic uncertainty relates to more general normative understandings of new genetic technologies and difficult-to-agree-upon terminology that exists in ecology as a discipline, while epistemic uncertainty stems from the technology still being in development (NASEM Citation2016). In seeking to restore ecosystems, a persistent normative question relates to how success should be evaluated. For example, there is room for disagreement in determining what would make a field trial sufficiently ‘adequate,’ depending on whether the trial’s goal is to test the ability of genetic biocontrol strategies to curb one versus many successive generations of rodents. It can also be difficult for field trials to speak to how successive releases of GE could affect other areas of a complex ecosystem over various lengths of time. More broadly, when eradication campaigns are applied, should success be determined when invasive species are eliminated, or merely curtailed, and if the latter, what would count as curtailed, and for how long would the curtailment need to last? Similarly, is the goal to restore the ecosystem to the state it was 5 years ago, 10 years ago, or at some other point in time? And again, for how long would the ecosystem need to stay that way?
Regarding epistemic uncertainty, much of the biological risk has to do with containment. There is a possibility, however slight, that an engineered rodent could mate with a closely related, non-invasive species and spread the gene drive system (Esvelt et al. Citation2014). Quantifying this possibility is difficult – even with present-day advanced genetic analysis tools, when and why species hybridize and the ability to hybridize is still not well understood (Harrison and Larson Citation2014). Engineered rodents on a contained island could also escape to mainland populations and potentially affect more than the intended target, a risk noted by Esvelt et al. (Citation2014), who also propose first using a CRISPR gene drive mechanism to introduce an ‘innocuous’ sequence to help mitigate the risk of unintended spread. This would be a genetically inert sequence that is not found in other organisms, which could be used as a target sequence for inserting the functional gene drive system (Esvelt et al. Citation2014). Targeting a specifically designed sequence could reduce the biological risk of the gene drive mechanism spreading to other populations of mice or related species of rodents. Especially in cases when an engineered gene is not self-limiting, the potential for global population suppression or extinction could become an issue of both conservation and international concern. To understand how a gene drive would behave in the field and develop proper containment measures, we, along with other researchers, are looking at gene drive systems that have already been implemented in insects, performing behavioral and genetic experiments on rodents in the lab, and using ecological modeling. However, it is impossible to eliminate epistemic uncertainty regarding how the technique would work in the field without field trials in a specific environment.
There is also epistemic uncertainty regarding how the fitness of the engineered rodents would compare to that of wild, invasive rodents. There are numerous genetic (individual and population level), neurological, and behavioral differences between laboratory and wild rodents, with laboratory rodents generally showing a narrower range of phenotypes than their wild counterparts (Koolhaas et al. Citation2010; Fonio, Benjamini, and Golani Citation2012; Chalfin et al. Citation2014). Invasive rodents can also undergo morphological changes while establishing a population on an island (Parkes Citation2008; Pergams et al. Citation2015), as they need to adapt to a variety of habitats quickly in order to survive (Berry Citation1996; Harper et al. Citation2015). Wild rodents also exhibit mating strategies such as polyandry and assortative mating, (Oakeshott Citation1974; Dean, Ardlie, and Nachman Citation2006; Manser, König, and Lindholm Citation2015) and have seasonal population fluctuations (Singleton et al. Citation2001), while laboratory rodents have very controlled reproductive environments. Thus, as rodent gene drive systems are being developed in laboratory strains, a primary research focus is whether laboratory rodents can survive in wild environments and compete reproductively with uniquely adapted wild rodents (also see Backus et al. Citation2016). Examining adaptations to different environments raises the question of how unique to the specific environment and context the engineered rodents would need to be to integrate into the invasive wild population. Would each island require backcrossing with unique wild stock rodents, or could a more general wild stock be backcrossed with engineered rodents to be used across islands? It is important to answer this question to understand the potential impact engineered rodents will have on the environments into which they are released.
Even once these questions are answered and an engineered rodent is shown to be able to survive and reproduce on an island, epistemic uncertainty would remain around how the spread of the gene drive construct itself would interact with a variety of genetic and ecological factors. Theoretical models suggest that the proposed Sry/t-haplotype system for mice would be unlikely to drive itself into a population with a single release (Backus and Gross Citation2016; or see the similar technique of autosomal X-chromosome shredders: Deredec, Burt, and Godfray Citation2008). As long as the genetic construct functions as expected and ecological conditions are not entirely unfavorable, the Sry/t-haplotype construct should be able to effectively suppress a population through multiple releases if the GE mice are released at a high enough rate relative to the wild population (Deredec, Burt, and Godfray Citation2008; Alphey Citation2014; Esvelt et al. Citation2014; Backus and Gross Citation2016). As long as the releases continue, the population should be suppressed unless resistance to the gene drive evolves. This resistance could take the form of mating behaviors that allow wild females to avoid producing offspring with gene drive males or mutations that disrupt the function of either the autosomal Sry or the t-haplotype (Burt Citation2003; Esvelt et al. Citation2014; Bull Citation2015; Lindholm et al. Citation2016). Other factors will come into play when using a system such as CRISPR/Cas9, which would likely result in a lower reduction in fitness (Esvelt et al. Citation2014; Min, Smidler, and Esvelt, Forthcoming). Testing these systems and rodents in the laboratory and the field is contingent on the development of the technology.
5. Social acceptability
As outlined above, genetic biocontrol of rodents would need to be sufficiently effective, minimize non-target impacts, eliminate or reduce animal welfare concerns, be affordable, and employ additional measures to maintain biosecurity before being considered a viable option. However, the potential application of genetic biocontrol of rodents will depend not only on technical assessment, but on the technology’s acceptability in specific socio-cultural contexts. Historically, the decision to apply a new technology has often relied on technical assessments that quantify and compare various options (e.g. cost–benefit analysis, traditional risk assessment) (Hill and Sendashonga Citation2003). More recent scholarship has emphasized the importance of more participatory methods that engage a wide variety of stakeholders (groups of people with direct professional or personal interest) and communities (e.g. groups of people who live on or near the island) in decision-making (NASEM Citation2016). In addition to advancing procedural justice (Besley and McComas Citation2005), deliberative engagement can allow researchers and developers to gain insights that make research – and the technologies it yields – more effective, producing knowledge that would not otherwise be gained (Bates et al. Citation2005; Sharpe Citation2014). When communication and public engagement are conducted in a flexible way that adapts to site- and audience-specific priorities, it is possible to uncover risks that would not be addressed by quantified technical assessments (Lavery et al. Citation2010), as well as potential areas for new research and development.
Acceptability regarding the application of a rodent gene drive may differ from previous biotechnology applications (Levidow, Carr, and Wield Citation2005). For example, because genetic biocontrol may be at odds with conventional moral norms that direct how humans should interact with the natural world, systems for controlling invasive rodent populations may be especially prone to criticism as proposed deployment sites are typically construed as wild places (NASEM Citation2016). The use of GE organisms may be viewed as unsuitable or incompatible with places viewed as wilderness and untouched by humans. In addition, some research suggests that cisgenic transformations (i.e. transformations using genetic material from the same rodent species) may be preferable to conventional transgenic transformations (i.e. inserting genes from a species of an unrelated taxa) among potential European consumers of agricultural genetically modified organisms (Delwaide et al. Citation2015). However, further investigation is warranted before assuming that cisgenic rodents would be favorably received.
More broadly, even if inclusive deliberations are undertaken to consider the potential application of these technologies, the scope of the questions considered will affect the outcome of any decision. For instance, the appeal of framing genetic biocontrol as an application specific to islands may stem in part from a desire for island applications to serve as proof of concept for genetic interventions in other locales, including mainland habitats. Accordingly, it would be valuable for further research to broaden the scope of this framing to consider issues such as containment, reversal, and ecological implications in non-island ecosystems, thereby offering points of comparison for the consideration of both scientific and regulatory audiences as well as stakeholders and communities. Given the uncertainties surrounding genetic biocontrol as well as the controversies that have been associated with past eradication campaigns, it may be equally valuable to expand the scope of public deliberations to examine what types of technology applications would be unacceptable, as well as acceptable, rather than focusing discussions on the narrower question of whether to pursue genetic biocontrol strategies. As gene drives and other emerging biotechnologies increasingly stretch the limits of current regulatory systems, opening up the scope of deliberation to include broad considerations, including questions about ethics and the human–nature relationship, could potentially help to establish a more adequate system of governance (Stirling Citation2008; Kuzma Citation2016).
6. Conclusions
Using gene drive technology to control or eradicate invasive rodent populations on islands holds promise. However, both the design of the engineered mouse and the research needed to assure its efficacy are only in the earliest stages. The limitations of the currently used toxicant approaches make development of gene drive technologies a potential alternative for conservation. Some of the advantages of using genetic biocontrol methods include that they may be considered more humane than toxicants, as the methods being proposed are non-lethal methods, and may require less time to implement and maintain than toxicants. At the same time, a number of areas of potential concern warrant consideration, including ecological risk, social acceptance, regulatory scrutiny, and ethical questions.
Acknowledgements
Valuable comments and support were given from participants of the following conferences: ‘Roadmap to Gene Drives: A Deliberative Workshop to Develop Frameworks for Research and Governance’, February 24–26, 2016, sponsored by North Carolina State University and the National Science Foundation Grant No. 1533990 and ‘SRY Mice: An International Workshop on Invasive Rodents’, October 13–16, 2015, sponsored by the United States Department of Agriculture Animal and Plant Health Inspection Service National Wildlife Research Center, North Carolina State University, Texas A&M University, Island Conservation, and the Commonwealth Scientific and Industrial Research Organisation.
Disclosure statement
No potential conflict of interest was reported by the authors.
Notes on contributors
Caroline Leitschuh is a doctoral candidate in Zoology, a member of the Genetic Engineering and Society Center and a member of the W.M. Keck Center for Behavioral Biology all at North Carolina State University. She studies how rearing environment affects mouse anxiety-related and exploratory behavior, in the context of eradication of invasive mice from islands. She also is interested in facilitating communication between disparate scientific disciplines and between researchers and the public.
Dona Kanavy earned her bachelors of science in clinical laboratory science at the University of North Carolina at Chapel Hill in 2009. She is currently a PhD student in genetics at North Carolina State University, and is finishing her research at Texas A&M. The primary focus of her research is using genetic technology to eradicate invasive rodents from islands.
Gregory Backus received his PhD in Biomathematics and Biological Sciences at North Carolina State University and is a member of the Genetic Engineering and Society Center. Using mathematical models, he studies the ecology and evolution eradicating invasive populations with gene drive technology.
Rene Xavier Valdez is a PhD student in the Fisheries, Wildlife, and Conservation Biology Program at North Carolina State University. His research focuses at the intersections of conservation and society, exploring how global drivers such as climate change, urbanization, and advancing biotechnology are shaping biodiversity conservation.
Megan Serr is a PhD candidate in Zoology and a member of the Genetic Engineering and Society Center at North Carolina State University. She received her BS and MS degrees in biology and is a former science educator. Her focus is on reproductive behavior and secondary invasions in house mice.
Dr Elizabeth Pitts is an assistant professor of English at the University of Pittsburgh. She studies how to better facilitate productive communication among citizens, scientists, policymakers, and other stakeholders when deliberating about the application of biotechnologies.
David Threadgill is a distinguished professor and Director of the Texas A&M Institute of Genome Sciences and Society. His laboratory uses mouse models to investigate genetic factors that contribute to inter-individual differences in health and disease. They currently focus on cancer susceptibility.
John Godwin is a professor of biological sciences and a member of the W.M. Keck Center for Behavioral Biology at North Carolina State University. Dr Godwin and his laboratory group study reproductive biology and the behavioral neuroendocrinology and neurogenomics of stress and anxiety in a number of models including the protogynous sex-changing bluehead wrasse, zebrafish, and wild-derived house mice.
ORCID
Dona Kanavy http://orcid.org/0000-0003-4770-485X
Gregory A. Backus http://orcid.org/0000-0003-3695-8168
Megan Serr http://orcid.org/0000-0001-7599-6605
Additional information
Funding
Related Research Data
References
- Akbari, O., H. Bellen, E. Bier, S. Bullock, A. Burt, G. Church, K. Cook, et al. 2015. “Safeguarding Gene Drive Experiments in the Laboratory.” Science 349: 927–929. doi:10.1126/science.aac7932.
- Akbari, O., K. Matzen, J. Marshall, H. Huang, C. Ward, and B. Hay. 2013. “A Synthetic Gene Drive System for Local, Reversible Modification and Suppression of Insect Populations.” Current Biology 23: 671–677. doi:10.1016/j.cub.2013.02.059.
- Alphey, L. 2014. “Genetic Control of Mosquitoes.” Annual Review of Entomology 59: 205–224. doi:10.1146/annurev-ento-011613-162002.
- Angel, A., R. Wanless, and J. Cooper. 2009. “Review of Impacts of the Introduced House Mouse on Islands in the Southern Ocean: Are Mice Equivalent to Rats?” Biological Invasions 11: 1743–1754. doi:10.1007/s10530-008-9401-4.
- Backus, G., and K. Gross. 2016. “Genetic Engineering to Eradicate Invasive Mice on Islands: Modeling the Efficiency and Ecological Impacts.” Ecosphere 7: e01589. doi:10.1002/ecs2.1589.
- Backus, G., C. Leitschuh, A. Ludvik, E. Pitts, M. Serr, and R. Valdez. “Island Mice: Conserving Island Biodiversity.” NC State Genetic Engineering and Society Center. Accessed March 20, 2016. https://research.ncsu.edu/ges/research/igert/student-research/island-mice-conserving-island-biodiversity/.
- Bates, B., J. Lynch, J. Bevan, and C. Condit. 2005. “Warranted Concerns, Warranted Outlooks: A Focus Group Study of Public Understandings of Genetic Research.” Social Science and Medicine 60: 331–344. doi:10.1016/j.socscimed.2004.05.012.
- Bauer, H., J. Willert, B. Koschorz, and B. Herrmann. 2005. “The t Complex-encoded GTPase-Activating Protein Tagap1 Acts as a Transmission Ratio Distorter in Mice.” Nature Genetics 37: 969–973. doi:10.1038/ng1617.
- Berry, R. 1996. “Small Mammal Differentiation on Islands.” Philosophical Transactions: Biological Sciences 351: 753–764. doi:10.1098/rstb.1996.0070.
- Besley, J. C., and K. A. McComas. 2005. “Framing Justice: Using the Concept of Procedural Justice to Advance Political Communication Research.” Communication Theory 15: 414–436. doi:10.1093/ct/15.4.414 doi: 10.1111/j.1468-2885.2005.tb00342.x
- Brown, P. R., and G. Singleton. 2000. “Impacts of House Mice on Crops in Australia – Costs and Damage.” Paper six in Natural Resources Management and Policy Commons: Human Conflicts with Wildlife: Economic Considerations.
- Bull, J. J. 2015. “Evolutionary Decay and the Prospects for Long-term Disease Intervention Using Engineered Insect Vectors.” Evolution, Medicine, and Public Health 2015: 152–166. doi:10.1093/emph/eov013.
- Burt, A. 2003. “Site-specific Selfish Genes as Tools for the Control and Genetic Engineering of Natural Populations.” Proceedings of the Royal Society of London B 270: 921–928. doi:10.1098/rspb.2002.2319.
- Campbell, K., J. Beek, C. Eason, A. Glen, J. Godwin, F. Gould, N. Holmes, et al. 2015. “The Next Generation of Rodent Eradications: Innovative Technologies and Tools to Improve Species Specificity and Increase Their Feasibility on Islands.” Biological Conservation 185: 47–58. doi:10.1016/j.biocon.2014.10.016.
- Carroll, L., S. Meagher, L. Morrision, D. Penn, and W. Potts. 2004. “Fitness Effects of a Selfish Gene (the Mus t Complex) Are Revealed in an Ecological Context.” Evolution 58: 1318–1328. doi:10.1554/03-544 doi: 10.1111/j.0014-3820.2004.tb01710.x
- Caut, S., E. Angulo, and F. Courchamp. 2009. “Avoiding Surprise Effects on Surprise Island: Alien Species Control in a Multitrophic Level Perspective.” Biological Invasions 11: 1689–1703. doi:10.1007/s10530-008-9397-9.
- Caut, S., J. Casanovas, E. Virgos, J. Lozano, G. Witmer, and F. Courchamp. 2007. “Rats Dying for Mice: Modelling the Competitor Release Effect.” Austral Ecology 32, 858–868. doi:10.1111/j.1442-9993.2007.01770.x.
- Centers for Disease Control and Prevention. “Rodents.” CDC.gov. Accessed March 20, 2016.
- Chalfin, L., M. Dayan, D. Levy, S. Austad, R. Miller, F. Iraqi, C. Dulac, and T. Kimchi. 2014. “Mapping Ecologically Relevant Social Behaviours by Gene Knockout in Wild Mice.” Nature Communications 5: 4569. doi:10.1038/ncomms5569.
- Clavero, M., and E. García-Berthou. 2005. “Invasive Species Are a Leading Cause of Animal Extinctions.” TRENDS in Ecology and Evolution 20: 110. doi:10.1016/j.tree.2005.01.003.
- Collins, P., B. Latta, and G. Roemer. 2009. “Does the Order of Invasive Species Removal Matter? The Case of the Eagle and the Pig.” PLoS ONE 4: e7005. doi:10.1371/journal.pone.0007005.
- Courchamp, F., M. Langlais, and G. Sugihara. 2000. “Rabbits Killing Birds: Modelling the Hyperpredation Process.” Journal of Animal Ecology 69: 154–164. doi:10.1046/j.1365-2656.2000.00383.x.
- Courchamp, F., R. Woodroffe, and G. Roemer. 2003. “Removing Protected Populations to Save Endangered Species.” Science 302: 1532. doi:10.1126/science.1089492.
- Crafford, J., and C. Scholtz. 1987. “Quantitative Differences between the Insect Faunas of Sub-Antarctic Marion and Prince Edward Islands: A Result of Human Intervention?” Biological Conservation 40: 255–262. doi:10.1016/0006-3207(87)90119-4.
- Curtis, C. 1968. “Possible Use of Translocations to Fix Desirable Genes in Insect Pest Populations.” Nature 218: 368–369. doi:10.1038/218368a0.
- David, A., J. Kaser, C. Morey, A. Roth, and D. Andow. 2013. “Release of Genetically Engineered Insects: A Framework to Identify Potential Ecological Effects.” Ecology and Evolution 3: 4000–4015. doi:10.1002/ece3.737.
- Davis, S., N. Bax, and P. Grewe. 2001. “Engineered Underdominance Allows Efficient and Economical Introgression of Traits into Pest Populations.” Journal of Theoretical Biology 212: 83–98. doi:10.1006/jtbi.2001.2357.
- Dean, M., K. Ardlie, and M. Nachman. 2006. “The Frequency of Multiple Paternity Suggests that Sperm Competition is Common in House Mice (Mus domesticus).” Molecular Ecology 15: 4141–4151. doi:10.1111/j.1365-294X.2006.03068.x.
- Delwaide, A.-C., L. Nalley, B. Dixon, D. Danforth, R. Nagaya, E. Van Loo, and W. Verbeke. 2015. “Revisiting GMOs: Are There Differences in European Consumers’ Acceptance and Valuation for Cisgenically vs Transgenically Bred Rice?” PLoS One 10: 1–16. doi:10.1371journal.pone.0126060 doi: 10.1371/journal.pone.0126060
- Deredec, A., A. Burt, and H. Godfray. 2008. “The Population Genetics of Using Homing Endonuclease Genes in Vector and Pest Management.” Genetics 179: 2013–2026. doi:10.1534/genetics.108.089037.
- Didion, J., D. Threadgill, and F. Pardo-Manuel de Villena. 2012. “Origins of Invasive House Mouse Population on Southeast Farallon Island.” University of North Carolina, Chapel Hill and North Carolina State University. Presentation to the EBM meeting Marseille, France.
- Doherty, T., A. Glen, D. Nimmo, E. Ritchie, and C. Dickman. 2016. “Invasive Predators and Global Biodiversity Loss.” Proceedings of the National Academy of Sciences 113: 11261–11265. doi:10.1073/pnas.1602480113.
- Esvelt, K., A. Smidler, F. Catteruccia, and G. Church. 2014. “Concerning RNA-guided Drives for the Alteration of Wild Populations.” eLIFE. doi: 10.7554/eLife.03401
- Fonio, E., Y. Benjamini, and I. Golani. 2012. “Short and Long Term Measures of Anxiety Exhibit Opposite Results.” PLoS One 7: e48414. doi:10.1371/journal.pone.0048414.
- Harper, G., and N. Bunbury. 2015. “Invasive Rats on Tropical Islands: Their Population Biology and Impacts on Native Species.” Global Ecology and Conservation 3: 607–627. doi:10.1016/j.gecco.2015.02.010.
- Harper, G., M. van Dinther, J. Russell, and N. Bunbury. 2015. “The Response of Black Rats (Rattus rattus) to Evergreen and Seasonally Arid Habitats: Informing Eradication Planning on a Tropical Island.” Biological Conservation 185: 66–74. doi:10.1016/j.biocon.2014.11.044.
- Harrison, R., and E. Larson. 2014. “Hybridization, Introgression, and the Nature of Species Boundaries.” Journal of Heredity (Special Issue), 105: 795–809. doi:10.1093/jhered/esu033.
- Hill, R., and C. Sendashonga. 2003. “General Principles for Risk Assessment of Living Modified Organisms: Lessons from Chemical Risk Assessment.” Environmental Biosafety Research 2: 81–88. doi:10.1051/ebr:2003004.
- Holmes, N., K. Campbell, B. Keitt, R. Griffiths, J. Beek, C. Donlan, and K. Broome. 2015. “Reporting Costs for Invasive Vertebrate Eradications.” Biological Invasions 17: 2913–2925. doi:10.1007/s10530-015-0920-5.
- Howald, G., C. Donlan, J. Galván, J. Russell, J. Parkes, A. Samaniego, Y. Wang, et al. 2007. “Invasive Rodent Eradication on Islands.” Conservation Biology 21: 1258–1268. doi:10.1111/j.1523-1739.2007.00755.x.
- Ishizuka, M., T. Tanikawa, K. Tanaka, M. Heewon, F. Okajima, K. Sakamoto, and S. Fujita. 2008. “Pesticide Resistance in Wild Mammals – Mechanisms of Anticoagulant Resistance in Wild Rodents.” The Journal of Toxicological Sciences 33: 283–291. doi:10.2131/jts.33.283.
- Koolhaas, J., S. de Boer, C. Coppens, and B. Buwalda. 2010. “Neuroendocrinology of Coping Styles: Towards Understanding the Biology of Individual Variation.” Neuroendocrinology 31: 307–321. doi:10.1016/j.yfrne.2010.04.001.
- Kuzma, J. 2016. “Policy: Reboot the Debate on Genetic Engineering.” Nature 531: 165–167. doi:10.1038/531165a.
- Lavery, J., P. Tinadana, T. Scott, L. Harrington, J. Ramsel, C. Ytuarte-Nuñez, and A. James. 2010. “Towards a Framework for Community Engagement in Global Health Research.” Trends in Parasitology 26 (6): 279–283. doi:10.1016/j.pt.2010.02.009.
- Le Roux, V., J-L. Chapuis, Y. Frenot, and P. Vernon. 2002. “Diet of the House Mouse (Mus musculus) on Guillou Island, Kerguelen Archipelago, Subantarctic.” Polar Biology 25: 49–57. doi: 10.1007/s003000100310
- Levidow, L., S. Carr, and D. Wield. 2005. “European Union Regulation of Agri-Biotechnology: Precautionary Links between Science, Expertise and Policy.” Science & Public Policy 32 (4): 261–276. doi: 10.3152/147154305781779452
- Lindholm, A., K. Dyer, R. Firman, L. Fishman, W. Forstmeier, L. Holman, H. Johannesson, et al. 2016. “The Ecology and Evolutionary Dynamics of Meiotic Drive.” Trends in Ecology & Evolution 31: 315–326. doi:10.1016/j.tree.2016.02.001.
- Lindholm, A., K. Musolf, A. Weidt, and B. König. 2013. “Mate Choice for Genetic Compatibility in the House Mouse.” Ecology and Evolution 3: 1231–1247. doi:10.1002/ece.3.534 doi: 10.1002/ece3.534
- Lyon, M. 2003. “Transmission Ratio Distortion in Mice.” Annual Review of Genetics 37: 393–408. doi:10.1146/annurev.genet.37.110801.143030.
- Lyttle, T. 1991. “Segregation Distorters.” Annual Review of Genetics 25: 511–581. doi:10.1146/annurev.ge.25.120191.002455.
- Mackay, J., J. Russell, and E. Murphy. 2007. “Eradicating House Mice from Islands: Successes, Failures, and the Way Forward.” In Managing Vertebrate Invasive Species: Proceedings of an International Symposium, edited by G. Witmer, W. Pitt, and K. Fagerstone, 294–304. Fort Collins, CO: USDA/APHIS/WS, National Wildlife Research Center.
- Manser, A., B. König, and A. Lindholm. 2015. “Female House Mice Avoid Fertilization by t Haplotype Incompatible Males in a Mate Choice Experiment.” Journal of Evolutionary Biology 28: 54–64. doi:10.1111/jeb.12525.
- Min, J., A. Smidler, and K. Esvelt. Forthcoming. “Harnessing Gene Drive.” Journal of Responsible Innovation.
- National Academies of Sciences, Engineering, and Medicine. 2016. Gene Drives on the Horizon: Advancing Science, Navigating Uncertainty, and Aligning Research with Public Values. Washington, DC: The National Academies Press. doi:10.17226/23405.
- Oakeshott, J. 1974. “Social Dominance, Aggressiveness and Mating Success among Male House Mice (Mus musculus).” Oecologia 15: 143–158. doi:10.1007/BF00345742.
- Parkes, J. 2008. A Feasibility Study for the Eradication of House Mice from Gough Island. RSPB Research Report No.34. United Kingdom: The Royal Society for the Protection of Birds.
- Pergams, O., D. Byrn, K. Lee, and R. Jackson. 2015. “Rapid Morphological Change in Black Rats (Rattus rattus) After an Island Introduction.” PeerJ: e812. doi:10.7717/peerj.812.
- Piaggio, A., G. Segelbacher, P. Seddon, L. Alphey, E. Bennett, R. Carlson, R. Friedman, et al. 2017. “Is It Time for Synthetic Biodiversity Conservation?” Trends in Ecology & Evolution 32: 97–107. doi:10.1016/j.tree.2016.10.016.
- Russell, J., D. Towns, and M. Clout. 2008. “Review of Rat Invasion Biology: Implications for Island Biosecurity.” Science for Conservation, 1–54. Wellington: Department of Conservation.
- Sax, D., S. Gaines, and J. Brown. 2002. “Species Invasions Exceed Extinctions on Islands Worldwide: A Comparative Study of Plants and Birds.” The American Naturalist 160: 766–783. doi:10.1086/43877 doi: 10.1086/343877
- Sharpe, L. 2014. “Public Perspectives on Genetic Biocontrol Technologies for Controlling Invasive Fish.” Biological Invasions 16: 1241. doi:10.1007/s10530-013-0545-5.
- Singleton, G. 2003. “Impacts of Rodents on Rice Production in Asia.” IRRI Discussion Paper. Phillippines: International Rice Research Institute.
- Singleton, G., and C. Krebs. 2007. “The Secret World of Wild Mice.” In Mouse Biomed. Res. Hist. Wild Mice, Genet., edited by J. Fox, S. Barthold, C. Davisson, F. Quimby, and A. Smith, 25–51. Amsterdam: Elsevier Inc.
- Singleton, G., C. Krebs, S. Davis, L. Chambers, and P. Brown. 2001. “Reproductive Changes in Fluctuating House Mouse Populations in Southeastern Australia.” Proceedings of the Royal Society of London B 268: 1741–1748. doi:10.1098/rspb.2001.1638.
- Song, Y., S. Endepols, N. Klemann, D. Richter, F.-R. Matuschka, C.-H. Shih, M. Nachman, and M. Khon. 2011. “Adaptive Introgression of Anticoagulant Rodent Poison Resistance by Hybridization between Old World Mice.” Current Biology 21: 1296–1301. doi:10.1016/j.cub.2011.06.043.
- South East Farallon Islands Invasive House Mouse Eradication Project: EIS. 2013. http://www.regulations.gov docket #: FWS-R8-NWRS-2013-0036.
- Stirling, A. 2008. “‘Opening Up’ and ‘Closing Down’: Power, Participation, and Pluralism in the Social Appraisal of Technology.” Science Technology Human Values 33: 262–294. doi:10.1177/0162243907311265.
- Towns, D., I. Atkinson, and C. Daugherty. 2006. “Have the Harmful Effects of Introduced Rats on Islands Been Exaggerated?” Biological Invasions 8: 863–891. doi:10.1007/s10530-005-0421-z.
- WHO. 2014. “The Guidance Framework for Testing Genetically Modified Mosquitoes.” World Health Organization, Programme for Research and Training in Tropical Diseases.
- Wilkinson, I., and D. Priddel. 2011. “Rodent Eradication on Lord Howe Island: Challenges Posed by People, Livestock, and Threatened Endemics.” In Island Invasives: Eradication and Management, 508–514. Gland: IUCN.
- Willison, K., and M. Lyon. 2000. “A UK-centric History of Studies on the Mouse t-Complex.” International Journal of Developmental Biology 44: 57–63.
- Zavaleta, E., R. Hobbs, and H. Mooney. 2001. “Viewing Invasive Species Removal in a Whole-ecosystem Context.” TRENDS in Ecology & Evolution 16: 454–459. doi:10.1016/50169-5347(01)02194-2 doi: 10.1016/S0169-5347(01)02194-2