
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
A three dimensional (3D) stage-structured predator–prey model is proposed and analyzed to study the effect of intraguild predation with harvesting of the adult species. Time lags in reproduction and maturation of the organism are introduced in the system and conditions for local asymptotic stability of steady states of delay differential forms of the ODE model are derived. The length of the delay preserving the stability is also estimated. Moreover, it is shown that the system undergoes a Hopf bifurcation when the time lags cross certain critical values. The stability and direction of the Hopf bifurcations are determined by applying the normal form method and the center manifold theory. Computer simulations have been carried out to illustrate various analytical results.
Public Interest Statement
Organisms go through multiple life stages as they proceed from birth to death. Cannibalism has become a prevalent feature of stage-structured populations in both aquatic and terrestrial food webs. Two fishes,Parrotfish and Pterois Volitans in coral reef ecosystem, are considered for the study of interactions within and among the stage-structured populations. With the proliferation of Pterois Volitans, there is a reduction in the density of herbivorous reef-fish. The reduction in herbivory in coral reefs can lead to a change of community structure of coral reefs for which corals decline with the proliferation of seaweeds. In this paper it is observed that cannibalism of Pterois Volitans population ensures the release of predation pressure on Parrotfish population. With the proliferation of Pterois Volitans, the system becomes oscillatory. Harvesting of adult Pterois Volitans helps in stabilizing the system.
1. Introduction
Many organisms go through multiple life stages as they proceed from birth to death. A stage-structured model of population growth consists of juvenile and adult organisms where the juveniles have no reproductive ability (Wang & Chen, Citation1997). Field observations by Rudolf (Citation2007) suggest that cannibalism is a prevalent feature of stage-structured populations in both aquatic and terrestrial food webs. In this type of ecological interaction, two species of the same trophic level interact as predator and prey, known as intraguild predation (IGP). As stated by Polis, Myers, and Holt (Citation1989), IGP can be classified as symmetrical or asymmetrical. Symmetric IGP occurs when there is mutual predation between two species, whereas in asymmetrical interaction, one species consistently preys upon the other (Zhang, Chen & Neumann, Citation2000). Stage structure with cannibalism is an asymmetric IGP (Rudolf, Citation2008).
Two fishes, Parrotfish and Pterois Volitans are considered for the study of interactions within and among the stage-structured populations. As observed by Morris et al. (Citation2009), Pterois Volitans are one of the top levels of the food web in many coral reef environments. They are known to feed mostly on small fishes, which include juveniles of their own species. Apart from preying on Parrotfish, adult Pterois Volitans exhibits a distinct cannibalistic attitude towards its juvenile species. As observed by Goreau and Hayes (Citation1994), Hare and Whitfield (Citation2003), and Albins and Hixon (Citation2008), the proliferation of predatory Pterois Volitans reduces the population density of herbivorous Parrotfish by changing the community structure of coral reefs for which corals decline with an increase in abundance of seaweeds. According to NOAA (National Oceanic and Atmospheric Administration), commercial harvesting of adult Pterois Volitans is required to reduce the numbers of Pterois Volitans to mitigate their impact on coral reef ecosystems (Morris & Whitfield, Citation2009).
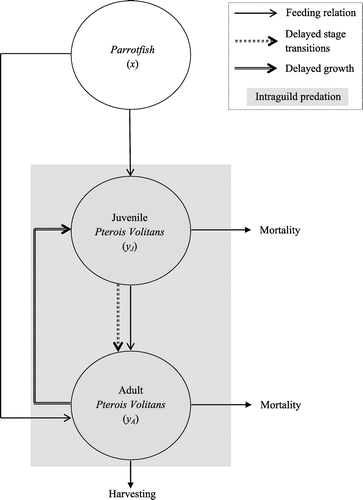
The role of time delay on ecosystem models has been investigated by Jiao, Yang, Cai and Chen (Citation2009), and Dhar and Jatav (Citation2013). Motivated by their works, we have studied a stage-structured system (Bhattacharyya & Pal, Citation2013) with Parrotfish at the first trophic level, and juvenile and adult Pterois volitans at the second and third trophic level, respectively. The model is based on the assumption that juvenile Pterois volitans can neither predate nor reproduce and the reproduction of adult Pterois volitans is mediated by some discrete time lag. The model used in this paper is a direct extension of the stage-structured model studied in (Bhattacharyya & Pal, Citation2013) with adult Pterois Volitans as IG-predator, juvenile Pterois Volitans as IG-prey, and Parrotfish as their common resource under the assumption that juveniles can predate, but do not reproduce. In the model, the IG-predator and IG-prey compete for Parrotfish, while the IG-predator exhibits asymmetric IGP. We assume that the reproduction of IG-predator is not instantaneous, but is be mediated by some discrete time delay required for egg deposition, embryo development, and hatching (Martin & Ruan, Citation2001). Further, we incorporate a discrete time lag, known as stage delay required by the IG-prey to become adult (Gourley & Kuang, Citation2004). The IG-predator is harvested to control its growth. We examine the effects of discrete time lags in reproduction and maturation of Pterois volitans on the dynamics of the IG-system. A schematic diagram of the system is given in Figure .
Figure 1. Schematic diagram of the intraguild system with delays.
Stability analysis of the systems are performed. We have estimated the delays for which the system preserves its stability around the positive steady state. We show that when the parameters representing reproduction and maturation time delay pass through some critical value, the interior equilibrium loses its stability and a Hopf bifurcation occurs. Numerical simulations under a default set of parameter values have been performed to support our analytical findings.
2. The basic model
We take a stage-structured model where Parrotfish is growing on the system with concentration at time
. The organisms, juvenile and adult Pterois Volitans, are introduced in the system with concentrations
and
, respectively, at time
. It is assumed that adult Pterois Volitans is cannibalistic and the juveniles have no reproductive ability—an exhibition of asymmetric IGP. The parameter
represents the delay in reproduction and
represents the delay in maturation of Pterois Volitans. Adult Pterois Volitans is harvested to control its growth.
We make the following assumptions in formulating the mathematical model:
| (H1) | In absence of Pterois Volitans, the growth equation of Parrotfish follows logistic growth with intrinsic growth rate | ||||
| (H2) | The per capita feeding rate of juvenile Pterois Volitans is assumed to follow Holling II functional response. Adult Pterois Volitans feed on Parrotfish and juvenile Pterois Volitans; accordingly the per capita feeding rate of adult Pterois Volitans follows a generalized form of Holling II functional response as a function of the concentrations of Parrotfish and juvenile Pterois Volitans. | ||||
| (H3) | The mortality rates of juvenile and adult Pterois Volitans are | ||||
| (H4) | Adult Pterois Volitans ( | ||||
| (H5) | Also, adult Pterois Volitans ( | ||||
with the initial conditions , where
the Banach space of continuous functions, mapping the interval
into
, where we define
.
Here represents the total time spent by Pterois Volitans in its juvenile stage and
is the harvesting rate of adult Pterois Volitans. Also
are the maximum growth rates,
are the half saturation constants, which are the concentrations of resource at which the growth rate of the organism is half maximal,
are the interference of Parrotfish and juvenile Pterois Volitans on the per capita growth rate of adult Pterois Volitans, and
are growth efficiencies (
); all of these are positive quantities.
For , we have
Therefore, for ,
implies
and
implies
, where
.
Thus, the system is well posed in if
,
, and
.
3. Invariance and boundedness of the System
Obviously, the right-hand sides of the equations in system are continuous nonnegative smooth functions on
. Indeed they are Lipschitzian on
and so the solution of the system
exists and is unique. Therefore, it is possible of Theorem
to prove that the interior of the positive octant of
is an invariant region.
Theorem 3.1
For large values of , all the solutions of the system
enter into the set
, where
, and
.
4. Qualitative analysis of the system without delay
In this section, we restrict ourselves to analyze the model in the absence of delay. Thus, only the interaction parts of the model system are taken into account. In the absence of delay, the system reduces to
(2)
(2)
where , and
The system will be permanent if there exist such that
, for each organism
in the system (Hofbauer & Sigmund, Citation1989). Permanence represents convergence on an interior attractor from any positive initial conditions and so it can be regarded as a strong form of coexistence (Ruan, Citation1993). From a biological point of view, permanence of a system ensures the survival of all the organisms in the long run.
Since as
it follows that there exist positive numbers
with
such that
and
for large values of
.
The following lemma rules out the possibility of extinction of any organism in the system under suitable conditions.
Lemma 4.1
If there exist finite positive real numbers with
,
, and
, then for large values of
each solution of
enters in the compact set
and remains in it finally.
The system possesses the following equilibria:
| (i) | organism-free equilibrium | ||||
| (ii) | Pterois Volitans-free equilibrium | ||||
| (iii) | the interior equilibrium | ||||
Lemma 4.2
The critical point of the system
is always unstable.
Thus, under no circumstances the system collapses.
The characteristic equation of the system at
is
, with one eigenvalue
and the remaining two eigenvalues will have negative real parts if
.
Lemma 4.3
The critical point of the system
is locally asymptotically stable if
.
Thus, with high harvesting rate of adult Pterois Volitans, the system stabilizes at .
The characteristic equation of the system at
is
where
,
and
.
Lemma 4.4
The critical point of the system
is locally asymptotically stable if
and
, where
and
.
Thus, harvesting of adult Pterois Volitans plays a critical role on the stability of the system at the interior equilibrium.
Lemma 4.5
The system undergoes a Hopf bifurcation at
iff
| (i) |
| ||||
| (ii) | |||||
| (iii) | |||||
The condition is equivalent to
where
Thus, using numerical methods, condition can be verified by showing that the curves
and
intersect at
, whereas the condition
can be verified by showing that the tangent to the curve
at
is not parallel to the
axis (Siekmann, Malchow & Venturino, Citation2008).
5. Qualitative analysis of the system with delays
In this section, we analyze the stability of the delayed systems at different equilibria and existence of local Hopf bifurcations.
The characteristic equation of the system at
is
.
Since , it follows that the critical point
of the systen
is always unstable.
The characteristic equation of the system at
is
, where
, and
.
The Jacobian of the system at
is
where so that
, and
.
The characteristic equation of the system at
is
, where
,
,
,
,
,
,
,
,
,
, and
.
In order to investigate the distribution of roots of the characteristic equations, we use the following Lemma by Ruan and Wei (Citation2003).
Lemma 5.1
For the transcendental equation , as
vary, the sum of orders of the zeros of
in the open right half-plane can change, and only a zero appears on or crosses the imaginary axis.
5.1. System with reproduction time delay only 
We now consider the case in which reproduction of the adult species is not instantaneous, but mediated by some discrete time lag .
In the absence of maturation time delay, the system reduces to
(3)
(3)
The characteristic equation of the system at
is
where
, and
.
Lemma 5.1.1
If is small, stability of the system
at
implies the stability of system
at
.
Proof
Let the system is locally asymptotically stable at
. Then
holds. If
is small, we can write
.
In this case, the characteristic equation of the system at
becomes
.
Since , all the roots of this equation have negative real parts, and so the stability of the system without delay at
implies the stability of system
at
.
Lemma 5.1.2
A sufficient condition for the system to be locally asymptotically stable at
is
and
, where
, and
Proof
The system is locally asymptotically stable at
for all
if the following conditions given by Gopalsamy (Citation1992) and Beretta and Kuang (Citation2002) hold:
| (a) | the real parts of all the roots of | ||||
| (b) | for all real | ||||
Sufficient conditions for the nonexistence of satisfying
can be written as:
.
Thus, for all real and for any
if
and
Also, the real parts of all the roots of are negative if
.
Therefore, the statement of lemma holds.
The characteristic equation of the system at
is
, where
, and
Lemma 5.1.3
If is small, stability of the system
at
implies the stability of system
at
.
Proof
Let the system is locally asymptotically stable at
. Then we have
and
.
For small , we can write
and so the characteristic equation of
at
becomes
, where
and .
Now, and so
.
Also, for , we have
and so for small
, we have
.
Therefore, for small , the stability of the system without delay at
implies the stability of system
at
.
Lemma 5.1.4
If the system is stable at
and if there exists a
in
such that
holds, then
is the maximum value
length of delay
for which the system
is locally asymptotically stable at
, where
, and
.
Proof
Let be the respective linearized variables of the model.
Then system can be expressed as
(4)
(4)
where .
Taking the Laplace transformation of system , we have
(5)
(5)
where and
The inverse Laplace transform of , and
will have terms which exponentially increase with time if
, and
have a pole with positive real parts. Since
needs to be locally asymptotically stable, it is necessary and sufficient that all poles of
, and
have negative real parts. We shall employ the Nyquist criterion, which states that if
is the arc length of a curve encircling the right half-plane, the curve
, and
will encircle the origin a number of times equal to the difference between the number of poles and the number of zeroes of
, and
in the right half-plane.
Let . Also, let
be the smallest positive root of
Then is locally asymptotically stable if
and .
Also,
Therefore, the positive solution of
is always greater than or equal to
.
We obtain where
is independent of
.
Also, where
and
Therefore, is an estimate for the length of delay for which the stability of the system at
is preserved.
We know that is a root of
if and only if
and
.
Eliminating , we obtain
, where
, and
Let and
. Then
takes the form
.
Lemma 5.1.5
Suppose that the conditions of Lemma are satisfied. Then the following results hold:
| (i) | If | ||||
| (ii) | If | ||||
| (iii) | If conditions in | ||||
Proof
If the conditions of Lemma hold, then the roots of,
have negative real parts.
Let be a root of
. For ease of notation, we denote
and
. Now,
and
gives
, where
.
| (i) | If | ||||
| (ii) | If | ||||
| (iii) | Let either | ||||
Thus, .
Therefore, if holds, then
and so the system
undergoes a Hopf bifurcation at
.
5.2. System with only maturation time delay
Now we consider the case in which the maturity time from juvenile to adult is mediated by a discrete time lag , while the reproduction process is instantaneous.
In the absence of reproduction time delay, the system reduces to
(6)
(6)
The characteristic equation of the system at
is
.
Lemma 5.2.1
If and
is small,
is always a saddle point of the system
.
Proof
For small values of we have
.
In this case, the characteristic equation of the system at
becomes
Since and
, it follows that at least one eigenvalue of the characteristic equation will always have positive real part and consequently,
is always a saddle point of the system
.
The characteristic equation of the system at
is
where
, and
.
Lemma 5.2.2
If is small and
, the stability of the system
at
implies the stability of system
at
.
Proof
Let the system be locally asymptotically stable at
. Then we have
.
If is small, we can write
. Then, the characteristic equation of the system
at
is
.
Since is small,
and so, the stability of the system
at
implies the stability of system
at
.
Lemma 5.2.3
A sufficient condition for the system to be locally asymptotically stable at
is
, and
, where
, and
Proof
implies
and
. This gives
, where
, and
Sufficient conditions for the nonexistence of satisfying
can be written as:
.
Thus, for all real and for any
if
, and
Also, the real parts of all the roots of are negative if
.
Therefore, the statement of lemma holds.
The characteristic equation of the system at
is
, where
, and
Lemma 5.2.4
If is small, stability of the system
at
implies the stability of system
at
.
Proof
Let the system be locally asymptotically stable at
. Then we have
, and
.
For small , we can write
and so the characteristic equation of
at
becomes
, where
,
, and
Now, and so
(since
is small);
.
Also, if , then for small
, we have
.
Thus, for small , the stability of the system
at
implies the stability of system
at
.
Lemma 5.2.5
If the system is stable at
and if there exists a
in
such that
holds, then
is the maximum value (length of delay) for which the system
is locally asymptotically stable at
, where
, and
.
Proof
Then system can be expressed as
(7)
(7)
where , and
.
Taking the Laplace transformation of system , we have
where
We shall employ the Nyquist criterion, which states that if is the arc length of a curve encircling the right half-plane, the curves
, and
will encircle the origin a number of times equal to the difference between the number of poles and the number of zeroes of
, and
in the right half-plane.
Let . Also, let
be the smallest positive root of
Then is locally asymptotically stable if
and
.
Now,
Therefore, the positive solution of
is always greater than or equal to
. We obtain
where
is independent of
.
Also,
where
, and
Therefore, is an estimate for the length of delay for which the stability of the system at
is preserved.
We know that is a root of
if and only if
and
.
Eliminating , we obtain
, where
, and
Let and
. Then
takes the form
. Then similar to Lemma
we get the following result
Lemma 5.2.6
Suppose that the conditions of Lemma are satisfied. Then the following results hold:
| (i) | If | ||||
| (ii) | If | ||||
| (iii) | If conditions in | ||||
5.3. System with both delays
The characteristic equation of the system at
is independent of
and is identical to the characteristic equation of the system
at
. Therefore, for all
and for small
satisfying
, the critical point
is always a saddle point of the system
.
Lemma 5.3.1
For small satisfying
, the stability of the system
at
implies the stability of system
at
.
Proof
Let the system be locally asymptotically stable at
.
Then we have and
.
If are small, we can write
and
.
The characteristic equation of the system at
becomes
.
Since ,
and so, the stability of the system without delay at
implies the stability of system
at
.
Lemma 5.3.2
For small satisfying
, stability of the system
at
implies the stability of system
at
.
Proof
Let the system be locally asymptotically stable at
. Then we have
, and
.
For small , we can write
and
and so the characteristic equation of
at
becomes
, where
,
and
Now, and so
(since
and
are small).
Also, if , then for small
, we have
.
Thus, for small , the stability of the system
at
implies the stability of system
at
.
Lemma 5.3.3
If the system is stable at
and if there exists
in
such that
holds, then
is the maximum value (length of delay) for which the system
is locally asymptotically stable at
, where
,
, and
.
Proof
Let be the respective linearized variables of the model. Then system
can be expressed as
(8)
(8)
where .
Taking the Laplace transformation of system , we have
We shall employ the Nyquist criterion, which states that if is the arc length of a curve encircling the right half-plane, the curves
, and
will encircle the origin a number of times equal to the difference between the number of poles and the number of zeroes of
, and
in the right half-plane.
Let . Also, let
be the smallest positive root of
Then is locally asymptotically stable if
and
.
Now,
Therefore, the positive solution of
is always greater than or equal to
, where
, and
.
We obtain where
is independent of
.
Also, where
.
This implies where
, and
Therefore, are estimates for the length of delays for which the stability of the system at
is preserved
.
Now, we consider in its stable interval and regard
as a parameter.
Without any loss of generality, we assume that Lemma and
hold.
Then the system is stable for all
.
Let be a root of
. Then,
and
.
Squaring and adding these equations we get , where
,
,
,
,
and
.
Let .
If , then
and as
it follows that
has finite positive roots, say,
.
For every fixed , there exists a sequence
such that
holds.
Let
When , the equation
has a pair of purely imaginary roots
for
.
We assume that .
By the general Hopf bifurcation theorem for functional differential equations, we get the following result:
Lemma 5.3.4
For the system , assume that Lemma
and
are satisfied for all
. Then, if
and
hold,
is locally asymptotically stable when
and the system
undergoes a Hopf bifurcation at
as
crosses
.
5.4. Direction and stability of the Hopf bifurcation
In this section, we shall study the bifurcation properties using techniques from normal form and center manifold theory by Hassard, Kazarinoff, and Wan (Citation1981). Throughout this section, we assume that system undergoes Hopf bifurcation at the positive equilibrium
for
, and
are the corresponding roots of
at
.
Without any loss of generality, we assume that , where
.
Also, let , and
, where
.
Then the system is transformed into a functional differential equation in
as
, where
,
, for
, and
are given by
and
(9)
(9)
where(10)
(10)
and
.
By Riesz representation theorem, there exists a matrix whose components are bounded variation functions ,
such that
for
.
In fact, we can choose
For , define
and
Then system is equivalent to
For , define
and a bilinear inner product , where
. Then
and
are adjoint operators. Now,
are eigenvalues of
and therefore they are also eigenvalues of
.
Let be an eigenvector of
corresponding to the eigenvalue
and
be an eigenvector of
corresponding to the eigenvalue
.
Then, gives
and
Also, gives
and
.
We have .
This gives,
Furthermore, .
Next, we study the stability of bifurcating periodic solutions. The bifurcating periodic solutions has amplitude
and nonzero Floquet exponent
with
. Under our hypothesis,
are given by
, and
.
The sign of indicates the direction of bifurcation, while that
determines the stability of
.
is stable if
and unstable if
. To derive the coefficients in these expansions, we first construct the coordinates to describe a center manifold
near
, which is a local invariant, attracting a two dimensional (2D) manifold.
Let and
.
On the manifold we have
, where
In fact, and
are local coordinates of center manifold
in the direction of
and
, respectively. The existence of center manifold
enables us to reduce
in a single complex variable on
. For the solutions
, since
, we have
, where
.
Therefore, , where
Now, gives
. This gives,
,
,
,
Since there are and
in
we need to compute them.
We have and so
Let , where
Differentiating with respect to
we get
.
Also, we have .
Therefore, .
Also,
Comparing the coefficients we get and
.
Now,
gives
and
where ,
,
, and
.
Now, implies
.
This gives , where
is a constant vector. Also,
implies
.
This gives , where
is a constant vector.
and
can be determined by setting
in
.
Since and
are continuous in
, we have
and
.
Next we focus on the computation of .
gives
(11)
(11) where
,
,
and .
Again, gives
(12)
(12) where
,
and
.
Thus, we can determine and
. Furthermore,
can be expressed by the parameters and delays. Then, we can obtain the expression of
, and
which determine the nature of bifurcating periodic solutions in the center manifold at the critical value.
6. Numerical simulations
In this section, we investigate numerically the effect of the various parameters on the qualitative behavior of the system using parameter values given in Table throughout, unless otherwise stated. Under the set of parameter values as given in Table , the systems ,
,
, and
are locally asymptotically stable at
.
Table 1. Set of parameter values used in numerical analysis.
Figure 2. Phase-plane diagrams of the systems for other parameter values in Table . All the systems are LAS at
.
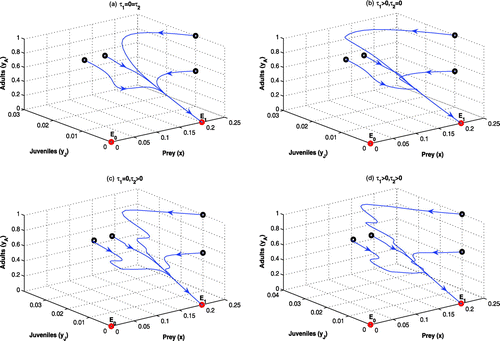
Figure 3. Time evolution of the populations for other parameter values in Table . The systems
and
are LAS at
, whereas, the systems
and
are oscillatory around
(solid). For
and
, other parameter values in Table , all the systems are LAS at
(dotted).
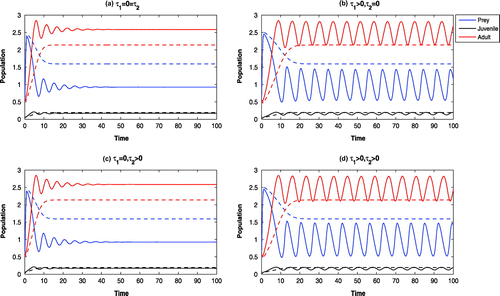
Figure 4. Time evolution of the populations for and other parameter values in Table . All the systems are oscillatory around
(solid). For
and
, other parameter values in Table , all the systems are LAS at
(dotted).
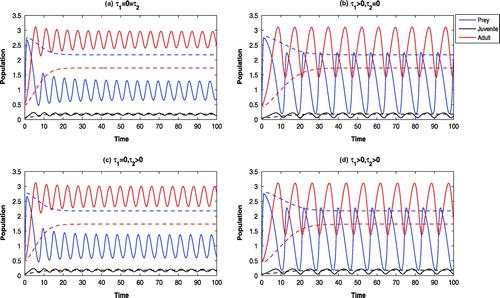
Figure 5. Time evolution of the populations for and other parameter values in Table . All the systems are oscillatory around
(solid). For
and
, other parameter values in Table , all the systems are LAS at
(dotted).
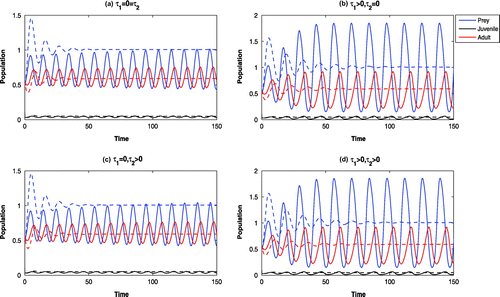
Figure 6. Time evolution of the populations for and parameter values in Table . The systems
and
are LAS at
, whereas, the systems
and
are oscillatory around
(solid). For
and
, other parameter values in Table , all the systems are LAS at
(dotted).
We carried out numerical simulations and interpret the resulting figures by varying the parameter(s) under investigation, keeping all other parameters fixed.
6.1. The effect of varying the carrying capacity ()
With low carrying capacity of Parrotfish (viz., ), the juvenile and adult Pterois Volitans cannot survive and all the systems become stable at
(cf. Figure ). By increasing the value of
(viz.,
), the system without delay and the system with maturation time delay only are stable at
, whereas, the systems with reproduction delay only and with both delays are oscillatory around
(cf. Figure (solid)).
With high carrying capacity of Parrotfish (viz., ), all the systems are oscillatory around
(cf. Figure (solid)).
6.2. The effect of varying the intrinsic growth rate ()
With low intrinsic growth rate of Parrotfish (viz., ), all the systems are oscillatory around
(cf. Figure (solid)). By increasing the value of
(viz.,
), the system without delay and the system with maturation time delay only are stable at
, whereas, the systems with reproduction delay only and with both delays are oscillatory around
(cf. Figure (solid)).
6.3. The effects of invasion
With high invasiveness of adult Pterois Volitans, the maximal growth rate of adult Pterois Volitans on Parrotfish becomes high. It is observed that, for the system without delay and the system with maturation time delay only are stable at
, whereas, the systems with reproduction delay only and with both delays are oscillatory around
(cf. Figure (solid)).
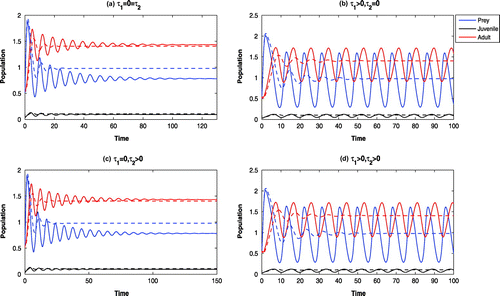
Figure 7. Time evolution of the populations for and other parameter values in Table . The systems
and
are LAS at
, whereas, the systems
and
are oscillatory around
(solid). For
and
, other parameter values in Table , all the systems are LAS at
(dotted).
With high growth rate of the adult Pterois Volitans (viz., ), all the systems become oscillatory around
(cf. Figure (solid)).
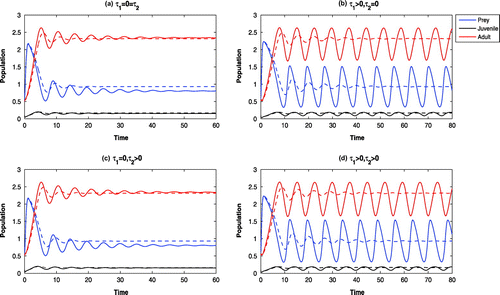
Figure 8. Time evolution of the populations for and other parameter values in Table . All the systems are oscillatory around
(solid). For
and
, other parameter values in Table , all the systems are LAS at
(dotted).
6.4. The effects of cannibalism
The rate of cannibalism is dependent on the maximal growth rate of adult maximal growth rate of on its juveniles. In absence of cannibalism (viz., ), all the systems become oscillatory around
(cf. Figure (solid)). Increasing the value of
(viz.
), the systems
and
are LAS at
, whereas, the systems
and
are oscillatory around
(cf. Figure (solid)). Further increase of
stabilizes the systems at
.
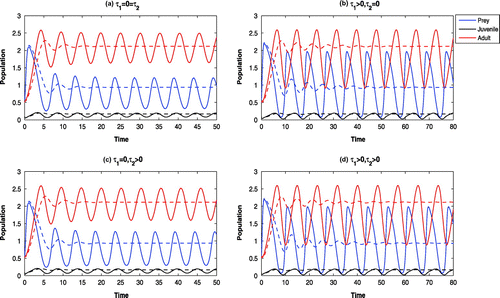
Figure 9. Time evolution of the populations for and other parameter values in Table . All the systems are oscillatory around
(solid). For
and
, other parameter values in Table , all the systems are LAS at
(dotted).
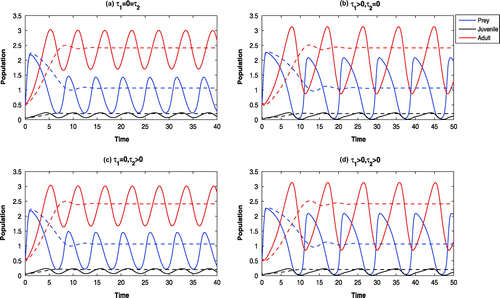
Figure 10. Time evolution of the populations for and other parameter values in Table . The systems
and
are LAS at
, whereas, the systems
and
are oscillatory around
(solid). For
and
, other parameter values in Table , all the systems are LAS at
(dotted).
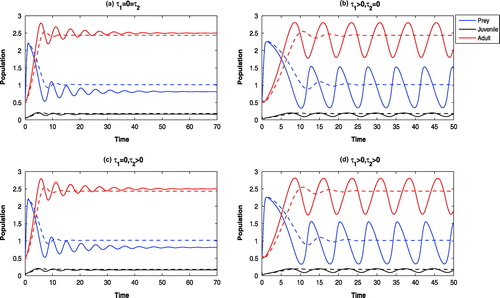
6.5. The effects of harvesting:
In absence of harvesting of adult Pterois Volitans the systems are oscillatory around (cf. Figure (solid)). By increasing the harvesting rate (viz.,
), the system without delay becomes stable at
and the systems with delay are oscillatory around
(cf. Figure (dotted)). Further increase of harvesting (viz.
) stabilizes all the systems at
. With high rate of harvesting, juvenile as well as adult Pterois Volitans are eliminated from the system. Further, we observe the following effects:
| (a) | The systems are oscillatory around | ||||
| (b) | With low intrinsic growth rate of Parrotfish (viz., | ||||
| (c) | With high invasiveness of the adult Pterois Volitans (viz., | ||||
| (d) | In absence of cannibalism of Pterois Volitans (viz., | ||||
Figure 11. Time evolution of the populations for and other parameter values in Table . The systems are oscillatory around
(solid). For
and other parameter values in Table , the systems
and
are LAS at
, whereas, the systems
and
are oscillatory around
(dotted).
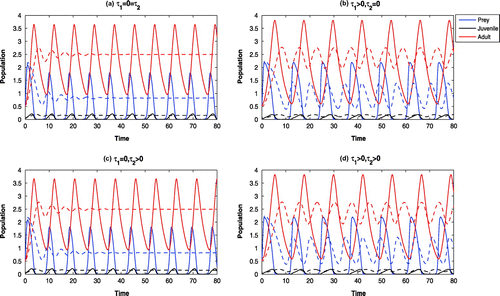
6.6. Hopf bifurcation
We observe that in absence of harvesting of adult Pterois Volitans, other parameter values as in Table , the system (2) is oscillatory around (Figure (solid)). Increasing the maximal rate of harvesting, the system
becomes locally asymptotically stable at
(Figure (dotted)). We therefore consider
as a bifurcation parameter. From Figure (a) it is observed that
and
intersect at
, indicating that the system
changes its stability when the parameter
crosses the threshold
. More specifically, for
we see that
, satisfying the condition of stability at
as as given in Lemma
. Also, for
we see that
and so the system
is unstable at
. Moreover, we observe that the tangent to
at
is not parallel to the
axis (Figure (b)), satisfying the condition
(Lemma
). Thus the system
undergoes a subcritical Hopf bifurcation when the parameter
is increased through
.
The bifurcation thresholds of the systems , and
, for different parameter values, are given in Table .
Table 2. Hopf bifurcation thresholds.
Figure 12. The relative position of
showing that the system
satisfies Lemma
when the two curves intersect at the
.
The tangent to the curve
at
is not parallel to the
axis, satisfying Lemma
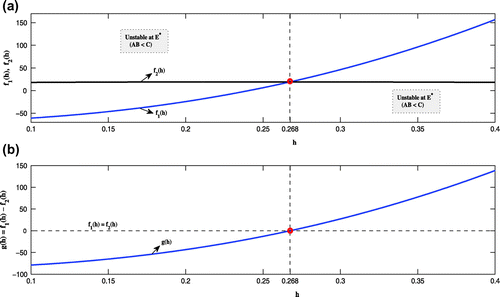
Figure 13. Bifurcation diagrams with as bifurcation parameter.
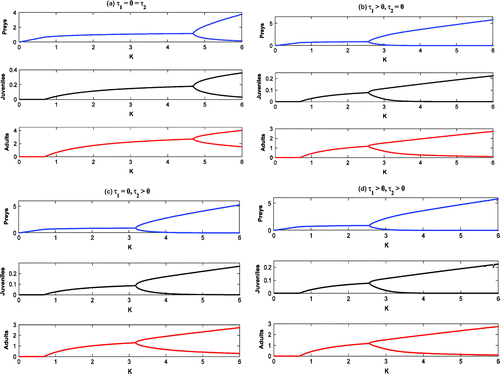
Figure 14. Bifurcation diagrams with as bifurcation parameter.
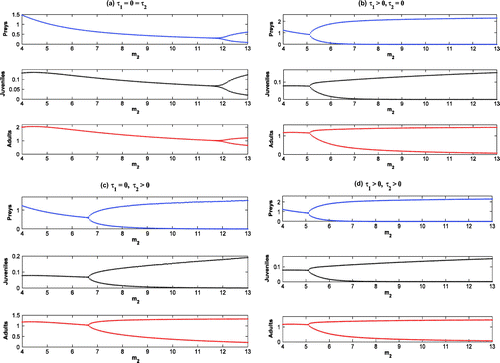
Figure 15. Bifurcation diagrams with as bifurcation parameter.
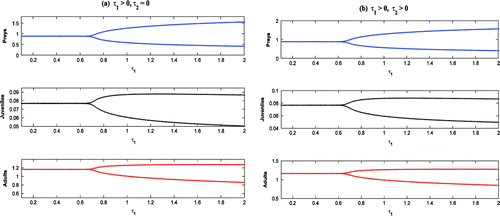
Figure 16. Bifurcation diagrams with as bifurcation parameter.
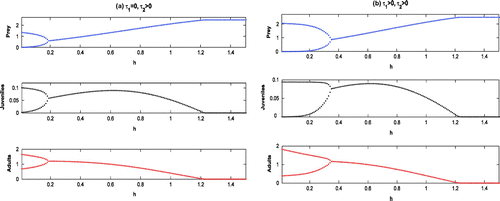
7. Discussion
We have considered an intraguild system in which adult Pterois Volitans exhibit cannibalism toward juveniles of its species and are subjected to harvesting. The main objective of this paper is to study the dynamic behavior of the system in the presence of discrete time lags in reproduction and maturation of Pterois Volitans. Experimental observations by Murray et al. (Citation2013) reveal that Holling II functional response is quite accurate in predicting the observed functional response of fishes. This prompts us to use Holling II response function in our model. Based upon the observations of Castillo-Chavez et al. (Citation2002) it is reasonable to assume that the mortality and maturity rates of fishes are proportional to the number of fishes present in our system. We have shown that solutions of the system are bounded in the long run. We have obtained conditions for permanence and the stability of the system at the coexistence equilibrium. It is observed that if the maximal harvesting rate of adult Pterois Volitans exceeds a certain threshold, the system stabilizes at the coexistence equilibrium through a subcritical Hopf bifurcation. Also, we have established that the systems remain locally asymptotically stable and all solutions approach whenever the magnitude of the delays lies below some threshold values. In order to find the expression for these threshold values, we have obtained the estimated length of stability preserving delays. The stability as well as the direction of bifurcation is obtained by applying the algorithm due to Hassard, Kazarinoff, and Wan (Citation1981) that depends on the centre manifold theorem. From numerical simulation it is observed that in the absence of cannibalism and with low harvesting rate of adult Pterois Volitans, the IG-systems become oscillatory around the interior equilibrium. In this case, increase of cannibalism up to a certain threshold stabilizes the system at the interior equilibrium, justifying the observations of Bosch, Roos, and Gabriel (Citation1988) that decrease of predator density due to cannibalism releases prey from predation pressure by inducing stability. Also, with high rate of cannibalism of adult Pterois Volitans, the coexisting populations show oscillatory dynamics. This supports the observations from previous modeling analyses by Diekmann et al. (Citation1986), Hastings (Citation1987), and Magnússon (Citation1999) that high cannibalism level can have destabilizing effects leading to oscillations. The IG-systems become oscillatory around the interior equilibrium with low intrinsic growth rate of Parrotfish in absence of harvesting of adult Pterois volitans, a phenomenon which has not been observed in the previous studies by Dhar and Jatav (Citation2013), Bhattacharyya and Pal (Citation2013), and deserve further investigation. If the reproduction time delay is high, the system with both time lags becomes oscillatory around the positive equilibrium. Also, high rate of invasion of adult Pterois Volitans on Parrotfish induces oscillations around the positive equilibrium, leading to dynamic instability. This represents the phenomenon of ecological imbalance due to the presence of the invasive Pterois Volitans in a coral reef ecosystem, justifying the observations of Albins and Hixon (Citation2008) that high predation rates of adult Pterois Volitans are detrimental to coral reef ecosystems. The dynamic instability can be controlled by increasing the maximal harvesting rate of adult Pterois Volitans. This too justifies that harvesting of these species is beneficial for coral reef ecosystem as observed by Morris, Shertzer and Rice (Citation2011).
Throughout the article an attempt is made to search for a suitable way to control the growth of Parrotfish and Pterois Volitans and maintain stable coexistence of all the species. From analytical and numerical observations, it is seen that increase of the harvesting of adult Pterois Volitans induces stability of the system. Moreover, we observe that harvesting at higher rates are necessary to obtain stability of the systems with delay than that of the system without delay.
Acknowledgements
The authors would like to thank the referees very much for their valuable comments and suggestions.
Additional information
Funding
Notes on contributors
Joydeb Bhattacharyya
Our research group is headed by Dr Samares Pal and includes Dr Joydeb Bhattacharyya as a post-doctoral researcher. Research in the group addresses a wide range of questions broadly concerning the dynamics of coral-reef ecosystems under natural and anthropogenic stress. The main emphasis of our work includes mathematical modeling of coral reef ecosystems affected by seaweed allelopathy-induced coral diseases, overfishing of herbivores and invasion of predatory Pterois Volitans. The research reported in this paper will help in studying the catastrophic regime shifts in coral-reefs under the proliferation of predatory Pterois Volitans and the subsequent loss in herbivory.
References
- Albins, M. A., & Hixon, M. A. (2008). Invasive Indo-Pacific lionfish ( Pterois Volitans) reduce recruitment of Atlantic coral-reef fishes. Marine Ecology Progress Series, 367, 233–238. doi:10.3354/meps07620.
- Beretta, E., & Kuang, Y. (2002). Geometric stability switch criteria in delay differential systems with delay dependent parameters. SIAM J. Math. Anal., 33, 1144–1165. doi:10.1137/s0036141000376086.
- Bhattacharyya, J., & Pal, S. (2013). Stage-structured cannibalism with delay in maturation and harvesting of an adult predator. Journal of Biological Physics, 39, 37–65. doi:10.1007/s10867-012-9284-6.
- Castillo-Chavez, C., Blower, S., Driessche, P., Kirschner, van d.D., & Yakubu, A. A. (2002). Mathematical approaches for emerging and reemerging infectious diseases: An introduction. Vol. 126 of the IMA volumes in mathematics and its applications. New York, NY: Springer.
- Dhar, J., & Jatav, K. S. (2013). Mathematical analysis of a delayed stage-structured predator-prey model with impulsive diffusion between two predators territories. Ecological Complexity, 16, 59–67. doi:10.1016/j.ecocom.2012.08.001.
- Diekmann, O., Nisbet, R. M., Gurney, W. S. C., , & van d Bosch, F., (1986). Simple mathematical models for cannibalism: A critique and a new approach. Mathematical Biosciences, 78, 21–46. doi:10.1016/0025-5564(86)90029-5.
- Gopalsamy, K. (1992). Stability and oscillations in delay differential equations of population dynamics. MA: Kluwer Academic Publishers.
- Goreau, T. J., & Hayes, R. (1994). Coral bleaching and ocean "hot spots". Ambio, 23, 176–180.
- Gourley, S. A., & Kuang, Y. (2004). A stage structured predator-prey model and its dependence on maturation delay and death rate. Journal of Mathematical Biology, 49, 188–200. doi:10.1007/s00285-004-0278-2.
- Hassard, B., Kazarinoff, N., & Wan, Y. (1981). Theory and applications of Hopf bifurcation. Cambridge (MA): Cambridge University Press.
- Hastings, A. (1987). Cycles in cannibalistic egglarval interactions. Journal of Mathematical Biology, 24, 651–666. doi:10.1007/BF00275508.
- Hofbauer, J., & Sigmund, K. (1989). On the stabilizing effect of predators and competitors on ecological communities. Journal of Mathematical Biology, 27, 537–548. doi:10.1007/bf00288433.
- Hare, J. A., & Whitfield, P. E. (2003). An integrated assessment of the introduction of lionfish ( Pterois volitans/miles complex) to the western Atlantic Ocean. NOAA Technical Memorandum, NOS NCCOS, 2, 1–21.
- Jiao, J., Yang, X., Cai, S., & Chen, L. (2009). Dynamical analysis of a delayed predator-prey model with impulsive diffusion between two patches. Mathematics and Computers in Simulation, 80, 522–532. doi:10.1016/j.matcom.2009.07.008.
- Magnusson, K. G. (1999). Destabilizing effect of cannibalism on a structured predator-prey system. Mathematical Biosciences, 155, 61–75. doi:10.1016/S0025-5564(98)10051-2.
- Martin, A., & Ruan, S. (2001). Predator-prey models with delay and prey harvesting. Journal of Mathematical Biology, 43, 247–267. doi:10.1007/s002850100095.
- Morris, J. A., Akins, J. L., Barse, A., Cerino, D., Freshwater, D. W., Green, S. J., Munoz, R. C., Paris, C., & Whitefield, P. E. (2009). Biology and ecology of invasive lionfishes, pterois miles and Pterois Volitans. Gulf and Caribbean Fisheries Institute, 61, 1–6.
- Morris, J. A., Jr, Shertzer, K. W., & Rice, J. A. (2011). A stage-based matrix population model of invasive lionfish with implications for control. Biological Invasions, 13, 7–12. doi:10.1007/s10530-010-9786-8.
- Morris, J.A. Jr, & Whitfield, P.E., (2009). Biology, ecology, control and management of the invasive Indo-Pacific lionfish: An updated integrated assessment. NOAA Technical Memorandum NOS-NCCOS, 99, 1–65.
- Murray, G. P. D., Stillman, R. A., Gozlan, R. E., & Britton, J. R. (2013). Experimental predictions of the functional response of a freshwater fish. Ethology, 119, 751-761. doi:10.1111/eth.12117.
- Polis, G. A., Myers, C. A., & Holt, R. D. (1989). The ecology and evolution of intraguild predation: Potential competitors that eat each other. Annual Review of Ecology and Systematics, 20, 297–330. doi:10.1146/annurev.ecolsys.20.1.297.
- Ruan, S. (1993). Persistence and coexistence in zooplankton-phytoplankton-nutrient models with instantaneous nutrient recycling. Journal of Mathematical Biology, 31, 633–654. doi:10.1007/bf00161202.
- Ruan, S., & Wei, J. (2003). On the Zeros of Transdental functions with applications to stability of delay differential equations with two delays. Dynamics of Continuous, Discrete and Impulsive Systems Series A: Mathematical Analysis, 10, 863–874.
- Rudolf, V. H. W. (2007). The interaction of cannibalism and omnivory: Consequences for communitty dynamics. Ecology, 88, 2697–2705. doi:10.1890/06-1266.1.
- Rudolf, V. H. W. (2008). The impact of cannibalism in the prey on predator-prey dynamics. Ecology, 89, 3116–3127. doi:10.1890/07-0709.1.
- Siekmann, I., Malchow, H., & Venturino, E. (2008). An extension of the Beretta-Kuang model of viral diseases. Mathematical Biosciences and Engineering, 5, 549–565. doi:10.3934/mbe.2008.5.549.
- van den Bosch, F., Roos, A. M., & Gabriel, W. (1988). Cannibalism as a life boat mechanism. Journal of Mathematical Biology, 26, 619–633. doi:10.1007/BF00276144.
- Wang, W., & Chen, L. (1997). A predator-prey system with stage structure for predator. Computers & Mathematics with Applications, 33, 83–91. doi:10.1016/S0898-1221(97)00056-4.
- Zhang, X., Chen, L., & Neumann, A. U. (2000). The stage-structured predator-prey model and optimal harvesting policy. Mathematical Biosciences, 168, 201–210. doi:10.1016/s0025-5564(00)00033-x.
Appendix 1
Proof of boundedness of the system (Theorem 3.1)
Let be a solution of the system
satisfying the initial conditions.
Then, corresponding to , there exists
such that
, for all
.
Let
Then for all we get,
, where
.
Let , where
.
Let be the solution of
satisfying
.
Then as
By comparison, it follows that proving the theorem.
Proof of Theorem 4.1
Since it follows that
.
Therefore, there exists such that
and
, for all
where
are finite positive constants with
.
For all , we have
This implies, for all
, where
and
.
Also, as
. Therefore, there exists
such that
for all
.
Therefore, for all .
Again, for all ,
if
.
Let there exists such that
.
Then for
and
and so in this case there exists
and
such that
for all
.
Therefore, for all , if
, then
.
Let . Then for all
, there exists finite positive real numbers
with
and
such that
.
Proof of Lemma (4.4)
The characteristic equation of the system at
is
, where
,
and
.
Now, if
, (say)
and .
Therefore, by the Routh–Hurwitz criterion of stability, the system is stable at
if
and
.
Proof of Lemma (4.5)
The characteristic equation of the variational matrix at is
, where
The necessary and sufficient conditions for Hopf bifurcation to occur at are that
and
,
where .
For , the characteristic equation becomes
.
For the roots are in general of the form:
, and
Therefore, gives
, where
Therefore,
If , then
.
Therefore, if
and
,
all hold, then a Hopf bifurcation occurs at and also it is nondegenerate.