
Abstract
Recent electroencephalography (EEG) studies accumulated evidence that support a dynamic view of the self (multiple aspect theory). In this study, we test the specific hypothesis of the multiple aspect theory of the self by using EEG data from the HisMine paradigm. In the HisMine paradigm German pronouns are visually presented in a stream of additional stimuli, while changes in brain activity are determined by means of EEG. At an early cortical processing stage, the presentation of the possessive pronouns "mein" ("my"), "sein" ("his") and "dein" ("your") elicited significantly different brain activities when compared to the non-personal pronoun "ein" ("a"). At a later processing stage, cortical processing of the self-related possessive pronoun ("mein") differed from the processing of all other possessive and the non-personal pronoun. In line with previous observations (and the multiple aspect theory of the self), the early effect (here referred to as Me1) was found at specifically left occipito-parietal electrode locations (e.g. PO9), whereas the later effect (here referred to as Me2) occurred specifically at the left fronto-temporal electrode site, F7. This finding supports the idea that the human self is consisting of multiple aspects. Potential implications are discussed.
Public Interest Statement
The present study confirms previous findings supporting the idea that the human self is not a singular construct, but consists of at least two aspects that are processed by separate brain structures as part of a hierarchy. A Me1 represents any personal engagement (more like a We), while a Me2 then forms the proper self that is distinct and different from other. Those two aspects could represent developmental stages during early childhood. Further, respective knowledge might help to better diagnose and treat clinical conditions that are associated with disordered self-concepts.
Competing Interests
The authors declare no competing interest.
1. Introduction
Since William James, the self has been a central topic in science and philosophy. With the advent of neuroimaging techniques, researchers started to investigate those brain structures involved in self-referential processing to better understand how brain activity contributes to the phenomenon of the self. Northoff and colleagues (Citation2006) conducted a meta-analysis including nearly almost all studies on self-referential processing that were published between 2000 and 2004. The studies included in the meta-analysis used functional magnetic resonance imaging or positron emission tomography in combination with a broad range of tasks. The meta-analysis revealed the so-called cortical midline structures (CMS) as a network commonly activated in association with self-referential processing independently from task and stimulus modality. A more recent meta-analysis of 107 neuroimaging studies confirmed that especially the ventral medial prefrontal cortex as part of the CMS as well as the left ventrolateral prefrontal cortex and the left anterior insula are frequently activated during self-processing tasks that require participants to judge whether a stimulus is related to the self or somebody else (Denny, Kober, Wager, & Ochsner, Citation2012).
While those imaging studies answered the question which brain structures are associated with self-referential processing, a number of recent studies have explored the time course of self-referential processing by means of electroencephalography (EEG) or magnetencephalography. The results of these studies support the idea that self-referential processing is a dynamic process and related to different aspects of the self.
Walla et al., for instance, conducted a series of EEG and MEG studies whose results confirmed a multiple aspect theory of the self. The studies used language stimuli such as possessive pronouns combined with nouns (e.g. “my garden”, “his garden” and “a garden”) to elicit self-referential versus other-referential processing (e.g. Walla, Duregger, Greiner, Thurner, & Ehrenberger, Citation2008; Walla, Greiner, Duregger, Deecke, & Thurner, Citation2007). By doing so, it could be shown that at around 200 ms post-stimulus onset, self-related stimuli such as “my garden” are processed similarly to other-related stimuli (“his garden”) but that self- and other-related stimuli are processed differently from stimuli having no personal reference (“a garden”). Later though brain activity patterns elicited by self-related stimuli (“my garden”) differed significantly from other-related stimuli (“his garden”) as well as from personally unrelated stimuli (“a garden”) at around 400 ms post-stimulus onset especially over left fronto-temporal areas. Taken together the results are in line with the ideas of the multiple aspect theory of the self (Walla et al., Citation2007). This theory assumes that the human brain first processes the self just as somebody else (other), but different from an un-personal condition (non- or un-personal self, Me1; more like a “We”), whereas only later then it considers information related to the self as distinct from information related to the other. This latter step might indicate the theoretical concept of an elaborate or proper self (personal self, Me2). The idea of two different aspects of the self is consistent with very early views about separate self-aspects such as the “Me” and the “I” that were suggested by James (Citation1890) and that distinguish between at least two aspects of the self.
So far, a number of further EEG studies that all used pronoun–noun pairs (e.g. Herbert, Herbert, Ethofer, & Pauli, Citation2011; Herbert, Pauli, & Herbert, Citation2010) or pronouns as stimuli (Blume & Herbert, Citation2014; Shi, Zhou, Liu, Zhang, & Han, Citation2010; Zhou et al., Citation2010) have accumulated evidence in support of a dynamic and multiple aspect view of the self. Nevertheless, the results of these studies have been mixed with regard to the processing stages at which discrimination between self from non-self and self from other occurs: Herbert, Herbert, Ethofer, and Pauli (Citation2011) used pronoun–noun pairs as stimuli. In contrast to Walla et al., pronoun–noun pairs could be of emotional or neutral content and self-referential or not (e.g. my fear, his fear, the fear, my garden, his garden and the garden). Participants’ task was to read the words silently while EEG was recorded. Modulation of early brain potentials differentiated self- and other-related pronoun–noun pairs from those without personal reference, but showed—as predicted from the multiple aspect theory—no differentiation between self- and other-related pronoun–noun pairs. Moreover and again in line with the multiple aspect theory, modulation of late event-related brain potentials showed a clear processing advantage for self-related emotional pronoun–noun pairs as compared to other-related or unreferenced emotional word pairs at about 500 ms after stimulus onset. However, notably all other stimuli including self-related neutral pronoun–noun pairs did not elicit any significant event-related potential (ERP) amplitude changes in later time windows.
Shi et al. (Citation2010) and Zhou et al. (Citation2010) as well as Blume and Herbert (Citation2014) investigated whether pronouns related to the self would be processed differently from pronouns unrelated to the self when these are presented visually either as rare stimuli in an oddball task or with the same presentation frequency amongst other stimuli in an RSVP design. While in the oddball tasks, Shi et al. and Zhou et al. found no generally significant differentiation between self- and other-related pronouns at early processing stages in line with the idea of a non-personal self, a near-significant trend towards larger P2 amplitudes for self- vs. other-related pronouns was observed at specifically the left frontal electrode site F7. Blume and Herbert (Citation2014) could demonstrate that when stimuli are presented in rapid streams at a rate of for instance 2.5 Hz, self-related pronouns are spontaneously processed preferentially compared to other-related pronouns already during initial processing stages starting as early as in the P1 time window. The latter findings challenge the assumption that processing of simple self-related stimuli such as possessive pronouns would always reliably elicit the two prior described self- aspects (including the Me1 as a non-personal self, unable to discriminate self from other and an elaborate self, the Me2 able to discriminate self from other and self from non-self). However, in all three pronoun studies mentioned before, no comparisons between self-related with unrelated pronouns were included for why it is unclear whether and if so at which processing stage differentiation between personal and non-personal information might have taken place. Similarly, all three studies focused on the analyses of robust ERP effects, whereas Walla et al. analysed the whole time frame of cortical activity without looking at discrete ERP modulation patterns. Perhaps then, locally more refined temporal analyses of ERP effects as reported by Walla et al. (Citation2007) could help reveal subtle temporal differences in stimulus processing that might have gone unnoticed in previous analyses focusing on modulation of ERP amplitudes.
The aim of the present paper therefore was to investigate the boundary conditions of the multiple aspect theory by determining whether or not simple visual presentations of pronouns would elicit the two prior described self-aspects (Me1 and Me2) when compared to un-personal pronouns. In particular, by using EEG data from the HisMine paradigm (Blume & Herbert, Citation2014), which comprises all personal and possessive as well as non-personal pronouns and articles of the German language will allow us to test the specific hypothesis of the multiple aspect theory of the self (Walla et al., Citation2007): Firstly, whether visual presentations of the German possessive pronouns “mein” (my) and “sein” (his) would elicit similar brain activity at early stages around 200 ms post-stimulus that differs from the non-personal pronoun (“ein”) (a). Secondly, whether processing of the possessive pronoun “mein” would enhance brain activity at around 400 ms post-stimulus separating self- from other-referential processing at left frontal areas.
2. Methods
2.1. Participants
Data from 28 native speakers of German (23 female, 5 male) with a mean age of 22.9 years (SD = 3.8) were analysed. Participants had normal or corrected-to-normal vision, and none of them had a history of prior somatic, neurological or psychiatric disorders. The study was conducted in accordance with the Declaration of Helsinki. All participants provided written informed consent. They were reimbursed financially or received course credit for their participation.
2.2. Stimuli
Stimuli consisted of the German pronouns “ich” (“I”), “du (“you”)”, “er” (“he”), (personal pronouns) and “mein” (“my”), “dein” (“your”), “sein” (“his”) (possessive pronouns) which were presented together with the pronoun “ein” (“a”) and the articles “es” (“it”) and “das” (“the”) containing no self- or other-reference. For the purpose of this study, we only analysed the “ein”, “mein”, “sein” and “dein” conditions. Previous analyses showed that visual presentation of the possessive pronoun “dein” might contain both, self- and other-reference, depending on the specificities of the task and the reader’s perspective (Blume & Herbert, Citation2014). In the present analysis, the “dein” condition was therefore also included as a further control and compared against non-personal pronouns. In line with the multiple aspect theory, it could be expected that the second-person possessive pronoun “dein” elicits brain activity similar to the first- and third-person possessive pronouns “mein” and “sein” at early stages, while also eliciting brain activity that differs from the “mein” condition at later stages.
2.3. Procedure
All words were presented visually in black font (Times font, 40 points) at the centre of a white computer screen. Stimuli were presented for 1,000 ms (1 Hz) in a serial visual stream. Each word was presented 60 times in a random sequence which controlled for frequency of occurrence such that each stimulus had the same transition probability. Participants were instructed to silently read the words. The experiment was programmed and controlled by Presentation® software (Neurobehavioral Systems, Inc.).
The processing condition under examination here was part of a larger experimental set-up consisting of different processing conditions that comprised passive viewing conditions that were followed by active counting conditions (for an overview see Blume & Herbert, Citation2014), which are not part of the present analysis. Here, data from only the 1 Hz passive viewing condition were reanalysed with regard to the hypothesis of the multiple aspect theory (Walla et al., Citation2007; Citation2008) to validate, if the different processing stages of the two aspects of the Me proposed by the multiple aspect theory of the self occur for the self-referential and other-referential possessive pronouns “mein” (“my”), “dein” (“your”) and “sein” (“his”) in relation to the non-personal pronoun “ein” (“a”). To this end, effects of interest were analysed at exactly the same electrode positions as suggested and previously reported by Walla and colleagues (see Walla et al., Citation2007).
2.4. Electroencephalography
Participants were seated on a comfortable chair at a distance of about 70 cm from the computer screen. Thirty-two active Ag/AgCl electrodes were placed on the scalp according to the international 10–20 system using an actiCAP® system (Brain Products GmbH). The electro-oculogram (EOG) was recorded from two Ag/AgCl electrodes placed above and below the participant’s right eye (VEOG) and from two electrodes that were fixed laterally to the outer canthus of each eye (HEOG). Impedance was kept below 10 kΩ and raw EEG signals were continuously recorded at a sampling rate of 500 Hz using a BrainVision BrainAmp® DC amplifier. Electrodes were connected to ground and referenced to the vertex (Cz).
2.5. Analysis
All EEG data were processed with EEGDISPLAY, a custom-made software package to process EEG data of various different formats (Ross Fulham). Raw EEG data files were downsampled from 500 to 256 samples per second. Then, data were visually inspected and a band pass filter from .1 to 30 Hz was applied. An EEGDISPLAY inbuilt algorithm was used for eye blink correction and epochs were generated by using a 100 ms baseline (for baseline correction) and a 1,000 ms post-stimulus period. Finally, averages were calculated across single trials for each of the four conditions of interest (“mein”, “sein”, “dein” and “ein”) corresponding to “my”, “his” “your” and “a”.
ERPs were generated to display brain potential changes over time for all four conditions of interest at two selected electrode sites (F7 and PO9) (see Figure ). For the purpose of statistical analysis, the post-stimulus epoch was split into 39 × 26 ms time windows and all data points within these time windows were averaged to result in single amplitude values. Short intervals were chosen to keep statistical temporal resolution at a high level. The mean amplitudes were then taken as dependent variables and subjected to a 4 (factor condition: “ein”, “mein”, “sein”, “dein”) × 2 (factor electrode: PO9, F7) repeated measures (RM) analysis of variance (ANOVA). One RM ANOVA was calculated for each time interval separately. Also, topographical maps were created for all four conditions and t-maps were calculated to show significant differences between the conditions. As already mentioned, the selection of electrodes was based on previous EEG and MEG studies (see for an overview Walla et al., Citation2007, Citation2008) which revealed that the Me1 and Me2 effects are particularly pronounced at these two electrodes.
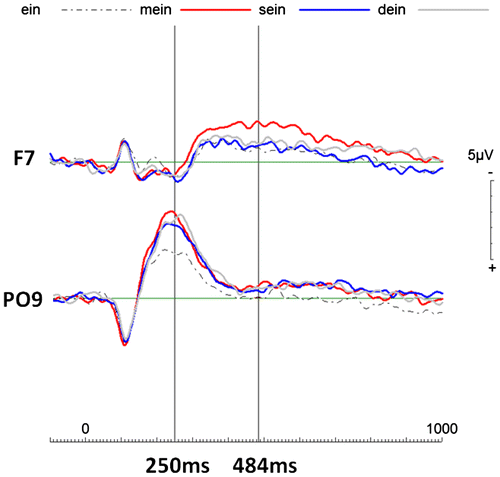
Figure 1. Event-related potentials (ERPs) of all four pronouns of interest (possessive pronouns: “mein” (my), “sein” (his), “dein” (your) and non-personal pronoun “ein” (a)).
3. Results
3.1. EEG
The ERPs shown in Figure clearly demonstrate that at electrode location PO9 (left occipito-parietal), the possessive pronouns “mein” (“my”), “dein” (“your”) and “sein” (“his”) elicited similar brain activity that differs from the non-personal pronoun “ein” (“a”). This difference between any person and no person starts before 200 ms post-stimulus onset (see Figure ). This early neural phenomenon is here referred to as the Me1 (non-personal self). At the left frontal electrode (F7), brain activity elicited by the self-related possessive pronoun “mein” (“my”) differed from brain activity elicited by all other pronouns (“ein” (“a”), “dein” (“your”) and “sein” (“his”)) from about 250 ms until about 700 ms post-stimulus onset (maxim effect at around 484 ms post-stimulus onset). This later phenomenon is here referred to as the Me2 (proper and actual self).
The repeated measure ANOVAs revealed condition main effects starting from 224 ms after stimulus onset until the end of the entire epoch (1,000 ms). This result confirms that the conditions of interest indeed elicited significantly different brain activities. Further, condition × electrode interactions occurred between 146 ms and 328 ms post-stimulus (see Table ). The early condition × electrode interactions are resulting from PO9-specific Me1 effects, whereas the later condition main effects without electrode interactions result from differences in electrophysiological activity at both electrode locations. However, the expected pattern of distinct brain activity differences was only found at F7 (see Figures and ).
Table 1. Consecutive repeated measures ANOVA results for every single post-stimulus time interval showing p-values (including F value and observed power) related to main effects (cond) and cond × electrode interactions (c × ele)
Figure 2. Sample t-maps demonstrating that at early time intervals all possessive pronouns elicited significantly different electrophysiological activity compared to the no-personal pronoun “ein” (a). Later, here exemplified at the maximum effect (484 ms), only processing of the self possessive pronoun “mein” (my) differs from the processing of the non-personal pronoun “ein” (a).
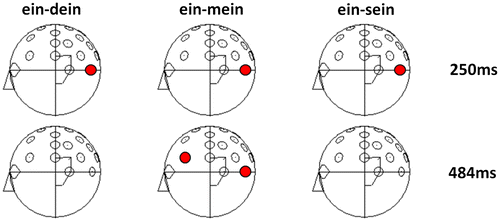
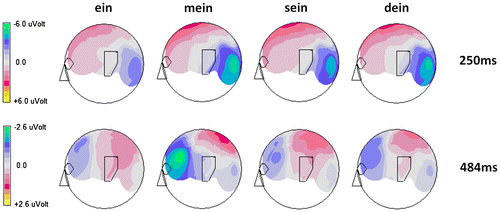
T-tests comparing the no-person condition (“ein” (a)) with all person conditions separately (“ein-dein” (a-your), “ein-mein” (a-my) and “ein-sein” (a-his)) for every single time interval revealed that indeed all personal possessive pronouns elicited different electrophysiological activity compared to the no-personal pronoun at the left occipito-parietal electrode (PO9) at an early stage (exemplified for maximum effects at 250 ms post-stimulus in Figure ), whereas the exclusive difference between the no-personal pronoun “ein” (a) and the self-possessive pronoun “mein” (my) clearly occurred at a later stage of processing at the left frontal area (see Figure ). Finally, separate topographies for every single pronoun condition of interest show that around the early time period (250 ms post-stimulus) all three possessive pronouns are associated with increased left occipito-parietal negative going ERPs. Later, at around 484 ms the self-possessive pronoun “mein” (my) elicits stronger negativity as compared to all other conditions at the left frontal electrode site F7, replicating Walla et al. (Citation2007) (Figure ).
Figure 3. Brain topographic maps revealed that at early times (250 ms) the pronouns “mein” (my), “sein” (his) and “dein” (your) all elicited similar brain activities that differed from “ein” (a). On the other hand, later at 484 ms “mein” (my) elicited different brain activity than all other pronouns (“ein” (a), “sein” (his) and “dein” (your), respectively.
4. Discussion
4.1. General
The present paper analysed data from the HisMine Paradigm (Blume & Herbert, Citation2014) in order to test the boundary conditions of the multiple aspect theory of the self proposed by Walla et al. (Citation2007, Citation2008). As predicted from this theory, the current analysis revealed two distinct spatiotemporally separate effects distinguishing self- and other-related pronouns from non-personal pronouns in early time windows and self from other-related pronouns in later time windows. Moreover, in line with previous observations (e.g. Walla et al., Citation2007), the early effect occurred at specifically left occipito-parietal electrode locations (e.g. PO9), whereas the later effect was found specifically at the left fronto-temporal electrode site, F7. Although the present analysis was restricted to a few sensors, it exactly replicates findings of previous studies of Walla and colleagues who, using similar analyses strategies, observed identical cortical activity patterns during processing of pronoun–noun pairs. Thus, the present observations might provide support for the previously stated multiple aspect theory (Walla et al., Citation2007, Citation2008). This dynamic self-model assumes two different aspects of the self, the Me1 and the Me2. The Me1 is like a first low-level processing stage of the self that exists in the brain and understands the self as simply being personal or non-personal, but not different to other. Perhaps, this aspect of the self could also be seen as a “We” reflecting any personal versus no personal engagement. In contrast, the Me2 is defined as the personal self that clearly distinguishes between self and other.
In the literature, many distinctions between two and even more kinds (e.g. Neisser, Citation1988) of self have been made previously (e.g. minimal versus narrative self; pre-reflective versus reflective self; see Dor-Ziderman, Berkovich-Ohana, Glicksohn, & Goldstein, Citation2013 for definitions of other multiple aspects of the self). One such distinction was put forward by Esslen, Metzler, Pascual-Marqui, and Jancke (Citation2008) who additionally provided neurophysiological evidence to support a dynamic and multiple aspect view of the self by what the authors call a pre-reflective self and a reflective self (for a detailed distinction see Legrand, Citation2007). What they report does widely confirm to the notion of distinct aspects of self-processing as suggested by the multiple aspect theory, although their idea of pre-reflective and reflective self is conceptually different from the idea of a Me1 and Me2. Esslen et al. (see also Legrand, Citation2007) aimed to draw the distinction between a non-conscious, but distinctively self-related (pre-reflective) and a conscious (reflective) aspect of the self, which they investigated by means of sentences starting with the personal pronouns “I” or “He/She” to examine the pre-reflective self on one hand, and the reflective self on the other hand by asking subjects to judge trait adjectives as self- or other-related. Shi et al. (Citation2010) and Blume and Herbert (Citation2014) investigated the extent to which pre-reflective and reflective self is reflected in event-related brain potential (ERP) modulation patterns elicited by personal and possessive pronouns, but unexpectedly no significant difference in pre-reflective and reflective temporal processing of personal and possessive pronouns was found. Shi et al. reported only a marginally significant trend while in the study by Blume and Herbert self-related pronouns were processed preferentially regardless of pronoun type. Moreover, during early processing stages, ERP modulation also indicated that processing of possessive pronouns was more pronounced than processing of personal pronouns. Thus, whereas the extent to which the pre-reflective self and the reflective self is reflected in the processing of pronouns might be up to future research, more evidence from especially pronoun studies seems to exist for the distinction between a Me1 and Me2, although the observation of preferential processing of self-related possessive pronouns compared to other-related pronouns during initial processing stages are a challenge for both theories and should be addressed in future studies.
What could be the implications of a multiple and hierarchical aspect theory of self that distinguishes between a Me1 and a Me2?
4.2. Impact of multiple and hierarchical self-aspect theory and future research
The concept of a Me1 means only being a person (no difference between self and other). It forms an important basis for a more elaborate form of self, the Me2 and its underlying spatiotemporal dynamics. Decety and Sommerville (Citation2003) put together pieces from developmental and social psychology as well as neuroscience leading to the idea of a shared network between self and other whereby among others the inferior parietal cortex seems to play a crucial role. This goes well in line with previous findings (e.g. Herbert, Herbert, Ethofer, & Pauli, Citation2011; Walla et al., Citation2007), and the present observation showing similar brain activity at the occipito-parietal electrode PO9 for self- and other-related pronouns as compared to the non-personal pronoun “a” supporting the suggestion of a more primitive, non-personal Me1.
Theoretically, these two self-aspects interact, but still are distinct. The non-personal self (Me1) might exist without the personal self (Me2). The personal self however might not exist without the non-personal self, which can be interpreted as representing a hierarchical structure. Thinking in terms of evolutionary lineage, the Me1 might have evolved earlier than the Me2. This could mean that the Me1 might also exist in non-human mammals and also that a human being during early childhood starts off with only the Me1, while later on the Me2 develops. Future developmental studies using the HisMine paradigm (or similar approaches) will test this idea. In addition, besides further strengthening the multiple aspect theory of the human self, investigating the self from a dynamic point of view may also provide useful implications for clinical investigations.
Various psychological and neurological diseases are associated with disordered self-concepts or malfunctioning theory of mind capacities (e.g. Korkmaz, Citation2011). Disordered self in the schizophrenia spectrum (Parnas & Henriksen, Citation2014), self-defeating personality disorder (Reich, Citation1987) and autism (Uddin, Citation2011) are just a few of many more such diseases. Patients with disordered self-concepts may demonstrate selectively disordered Me1 or Me2 functions. Early lesion studies have shown deficient self-awareness and self-control in lesions of the right inferior parietal lobe (Posner & Rothbart, Citation1998; Stuss & Levine, Citation2002), which could be seen as an index for selective Me1 impairments. In such cases, it would be predicted that Me2 functions are affected as well. On the other hand, it is predicted that Me2 impairments can occur while leaving Me1 functions intact. Future EEG or MEG studies could test this prediction. The HisMine paradigm (Blume & Herbert, Citation2014) comprising all pronouns of the first, second and third person as well as non-personal pronouns would be perfectly suited to this end. Due to its simplicity, it could be even administered to severely impaired patients groups.
Previous imaging research showed that the CMS play an important role in self-referential processing. However, the precise role of this network is still unclear, especially with regard to the concepts of consciousness and awareness. Activation of CMS including the anterior cingulate cortex (ACC) has been found in a number of tasks including pronoun processing (e.g. Herbert, Herbert & Pauli, Citation2011; Shi et al., Citation2010). It might be related to attention (Carter et al., Citation1998; Elliott & Dolan, Citation1998; Devinsky, Morrell, & Vogt, Citation1995; Paus, Koski, Caramanos, & Westbury, Citation1998; Peterson et al., Citation1999; Posner & Rothbart, Citation1998; Weissman et al., Citation2004; Whalen et al., Citation1998). Given that attention is crucial for conscious processing, it might be that activations of CMS such as the ACC are mediating consciousness in concert with other networks or brain structures. One such other structure could be the insular cortex as suggested by Walla et al. (Citation2007). When both structures are simultaneously active self-awareness might be generated.
Although the present results are based on very locally restricted single electrode analyses and therefore cannot tell us anything about the localisation of the observed effects in the brain, they do support the idea that changes in brain activity and thus in self-referential processing do occur within the millisecond range. Importantly, these changes might have a crucial influence on whether midline structures are active simultaneously to self-referential processing (see Herbert, Herbert, Ethofer, Pauli, Citation2011; Herbert, Herbert, & Pauli, Citation2011; Northoff, Citation2013; Qin & Northoff, Citation2011) and whether or not self-referential processing is associated with self-awareness.
While this is only speculation, future research using the present design and following a dynamic approach of the self could reveal insight into various clinical conditions that are associated with disordered self-concepts including research on self-awareness and consciousness.
Author contribution
This manuscript is a shared first authorship and correspondence.
Acknowledgements
Many thanks to Ross Fulham for his highly appreciated data analysis contribution.
Additional information
Funding
Notes on contributors
Cornelia Herbert
Cornelia Herbert and Peter Walla form a cooperative partnership. Both authors contributed equally to this manuscript (shared first authorship and correspondence). Cornelia’s own individual neurophysiological focus is on the self, language and emotion, and the development of novel paradigms. Peter’s long-term interest focuses on an evolutionary perspective related to the development of the self. Their common applied interest is to unravel the boundary conditions of self-referential processing to contribute to a better understanding of clinical conditions with disordered self-aspects. In the present paper, EEG data from the HisMine paradigm was analysed with regard to the hypothesis of the multiple aspect theory of the self to test the validity and boundary conditions of this theoretical approach. The HisMine paradigm was developed by Cornelia Herbert within the framework of her grant HE5880/3-1.
References
- Blume, C., & Herbert, C. (2014). The HisMine-paradigm: A new paradigm to investigate self-awareness employing pronouns. Social Neuroscience, 9, 289–299.10.1080/17470919.2014.886616
- Carter, C. S., Braver, T. S., Barch, D. M., Botvinick, M. M., Noll, D., & Cohen, J. D. (1998). Anterior cingulate cortex, error detection, and the online monitoring of performance. Science, 280, 747–749.10.1126/science.280.5364.747
- Decety, J., & Sommerville, J. A. (2003). Shared representations between self and other: A social cognitive neuroscience view. Trends in Cognitive Sciences, 7, 527–533.10.1016/j.tics.2003.10.004
- Denny, B. T., Kober, H., Wager, T. D., & Ochsner, K. N. (2012). A meta-analysis of functional neuroimaging studies of self- and other judgments reveals a spatial gradient for mentalizing in medial prefrontal cortex. Journal of Cognitive Neuroscience, 24, 1742–1752.10.1162/jocn_a_00233
- Devinsky, O., Morrell, M. J., & Vogt, B. A. (1995). Review article. Brain, 118, 279–306.10.1093/brain/118.1.279
- Dor-Ziderman, Y., Berkovich-Ohana, A., Glicksohn, J., & Goldstein, A. (2013). Mindfulness-induced selflessness: A MEG neurophenomenological study. Frontiers in Human Neuroscience, 7, 582. doi:10.3389/fnhum.2013.00582
- Elliott, R., & Dolan, R. J. (1998). Activation of different anterior cingulate foci in association with hypothesis testing and response selection. NeuroImage, 8, 17–29.10.1006/nimg.1998.0344
- Esslen, M., Metzler, S., Pascual-Marqui, R., & Jancke, L. (2008). Pre-reflective and reflective self-reference: A spatiotemporal EEG analysis. NeuroImage, 42, 437–449.10.1016/j.neuroimage.2008.01.060
- Herbert, C., Herbert, B. M., Ethofer, T., & Pauli, P. (2011). His or mine? The time course of self-other discrimination in emotion processing. Social Neuroscience, 6, 277–288.10.1080/17470919.2010.523543
- Herbert, C., Herbert, B. M., & Pauli, P. (2011). Emotional self-reference: Brain structures involved in the processing of words describing one’s own emotions. Neuropsychologia, 49, 2947–2956.10.1016/j.neuropsychologia.2011.06.026
- Herbert, C., Pauli, P., & Herbert, B. M. (2010). Self-reference modulates the processing of emotional stimuli in the absence of explicit appraisal instructions. Social Cognitive and Affective Neuroscience, 6, 653–661.
- James, W. (1890). The principles of psychology. New York, NY: Holt.10.1037/11059-000
- Korkmaz, B. (2011). Theory of mind and neurodevelopmental disorders of childhood. Pediatric Research, 69, 101R–108R.10.1203/PDR.0b013e318212c177
- Legrand, D. (2007). Pre-reflective self-as-subject from experiential and empirical perspectives. Consciousness and Cognition, 16, 583–599.10.1016/j.concog.2007.04.002
- Neisser, U. (1988). Five kinds of self-knowledge. Philosophical Psychology, 1, 35–59.10.1080/09515088808572924
- Northoff, G. (2013). Unlocking the brain: Consciousness (Vol. 2). Oxford: Oxford University Press.
- Northoff, G., Heinzel, A., de Greck, M., Bermpohl, F., Dobrowolny, H., & Panksepp, J. (2006). Self-referential processing in our brain—A meta-analysis of imaging studies on the self. NeuroImage, 31, 440–457.10.1016/j.neuroimage.2005.12.002
- Parnas, J., & Henriksen, M. G. (2014). Disordered self in the schizophrenia spectrum. Harvard Review of Psychiatry, 22, 251–265.10.1097/HRP.0000000000000040
- Paus, T., Koski, L., Caramanos, Z., & Westbury, C. (1998). Regional differences in the effects of task difficulty and motor output on blood flow response in the human anterior cingulate cortex. NeuroReport, 9, R37–R47.10.1097/00001756-199806220-00001
- Peterson, B. S., Skudlarski, P., Gatenby, J. C., Zhang, H. P., Anderson, A. W., & Gore, J. C. (1999). An fMRI study of stroop word-color interference: Evidence for cingulate subregions subserving multiple distributed attentional systems. Biological Psychiatry, 45, 1237–1258.10.1016/S0006-3223(99)00056-6
- Posner, M. I. & Rothbart, M. K. (1998). Attention, self-regulation and consciousness. Philosophical Transactions of the Royal Society B: Biological Sciences, 353, 1915–1927.10.1098/rstb.1998.0344
- Qin, P., & Northoff, G. (2011). How is our self related to midline regions and the default-mode network? Neuroimage, 57, 1221–1233.
- Reich, J. (1987). Prevalence of DSM-III-R self-defeating (masochistic) personality disorder in normal and outpatient populations. The Journal of Nervous and Mental Disease, 175, 52–54.10.1097/00005053-198701000-00009
- Shi, Z., Zhou, A., Liu, P., Zhang, P., & Han, W. (2010). An EEG study on the effect of self-relevant possessive pronoun: Self-referential content and first-person perspective. Neuroscience Letters, 494, 174–179.
- Stuss, D. T., & Levine, B. (2002). Adult clinical neuropsychology: Lessons from studies of the frontal lobes. Annual Review of Psychology, 53, 401–433.10.1146/annurev.psych.53.100901.135220
- Uddin, L. Q. (2011). The self in autism: An emerging view from neuroimaging. Neurocase, 17, 201–208.
- Walla, P., Duregger, C., Greiner, K., Thurner, S., & Ehrenberger, K. (2008). Multiple aspects related to self-awareness and the awareness of others: An electroencephalography study. Journal of Neural Transmission, 115, 983–992.10.1007/s00702-008-0035-6
- Walla, P., Greiner, K., Duregger, C., Deecke, L., & Thurner, S. (2007). Self-awareness and the subconscious effect of personal pronouns on word encoding: A magnetoencephalography (MEG) study. Neuropsychologia, 45, 796–809.10.1016/j.neuropsychologia.2006.08.017
- Weissman, D. H., Gopalakrishnan, A., Hazlett, C. J., & Woldorff, M. G. (2004). Dorsal anterior cingulate cortex resolves conflict from distracting stimuli by boosting attention toward relevant events. Cerebral Cortex, 15, 229–237.10.1093/cercor/bhh125
- Whalen, P. J., Bush, G., McNally, R. J., Wilhelm, S., McInerney, S. C., Jenike, M. A., & Rauch, S. L. (1998). The emotional counting Stroop paradigm: A functional magnetic resonance imaging probe of the anterior cingulate affective division. Biological Psychiatry, 44, 1219–1228.
- Zhou, A. B., Shi, Z., Zhang, P. Y., Liu, P. R., Han, W., Wu, H. F., ... Xia, R. X. (2010). An ERP study on the effect of self-relevant possessive pronoun. Neuroscience Letters, 480, 162–166.10.1016/j.neulet.2010.06.033