
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Nitrogen is the major plant nutrient that limits common bean production throughout sub-Saharan Africa. Four experimental trials were conducted at four areas namely Babillae, Fedis, Haramaya, and Hirna experimental sites to determine if various inherent soil fertility status and soil total N might affect the N use efficiency of common bean var. Dursitu. Six levels of N application and two inoculation treatments were factorially combined and laid out in randomized complete block design with three replications. The agronomic efficiency of N (AE-N) by common bean obtained from different locations displayed significant difference at p ≤ 0.05. The highest AE-N was obtained from Babillae site, while the lowest from Hirna site. In general, overall AE-N declined with increase in rates of N application. Slight increase of AE-N was observed in Babillae and Hirna sites when 20 kg N ha−1 was applied. Haramaya and Hirna sites had the highest of all investigated growth parameters, except 100 seeds weight and harvest index. The regression analysis indicated strong and negative association (R 2 = 0.498 and R 2 = 0.390 at p ≤ 0.05) between AE-N and N rates of application in Babillae site followed by Fedis site, respectively. A stronger and significant association (R 2 = 0.276, p ≤ 0.05) of AE-N and grain yield was observed only in Hirna site. In general, this study indicated the remarkable effect of soil inherent fertility and soil total N content on N use efficiency of common bean in the study sites.
Public Interest Statement
The agronomic efficiency of N of common bean for different localities with various soil fertility is important to avoid unnecessary excess and low amounts of N application. This article described how the inherent soil fertility including the soil total N affects the use efficiency of N by common bean in major common bean growing areas of Eastern Ethiopia. This research work will help in further investigation on how soil with low inherent soil fertility influences the use efficiency of N by common bean.
Competing interests
The authors declare no competing interests.
1. Introduction
Common bean (Phaseolus vulgaris L.) is the chief source of dietary protein for most of Latin American and sub-Saharan African people, supplying about 20% of the protein intake per person (Centro Internacional de Agricultura Tropical, Citation2001; Van Rheenen, Citation1982). In Ethiopia, common bean is one of the major grain legumes cultivated, with its production concentrated in small farmers’ fields where the use of N fertilizer is limited and average yields are low, usually less than 1.3 ton ha–1 (Central Statistical Authority, Citation2013). Nitrogen is the most important element limiting crop production in the tropics (Sanchez, Citation1976). Previous surveys estimated that over 60% of the bean production areas in Central, Southern, and Eastern Africa was affected by N deficiency (Thung & Rao, Citation1999; Wortmann, Kirkby, Eledu, & Allen, Citation1998). This caused yield losses of up to 40% compared to the N-fertilized areas (Singh, Citation1999; Thung & Rao, Citation1999). Beside this, common bean is considered to be a poor fixer of atmospheric N when compared with other crop legumes (Graham, Citation1981; Piha & Munns, Citation1987) and generally responds poorly to inoculation of rhizobia in the field conditions (Buttery, Park, & Findlay, Citation1987; Graham, Viteri, Mackie, Vargas, & Palacios, Citation1982; Huntington, Smith, Thomas, & Blevins, Citation1986; Moxley, Hume, & Smith, Citation1986) and rarely derives more than 50% of their N from the atmosphere (Bliss, Citation1993; Wortmann, Citation2001). Moreover, Giller (Citation2001) indicated the nodule senescence of P. vulgaris often coincides with seed development, thus reducing the yield of common bean. However, Redden and Herridge (Citation1999) found that common bean responds to N fertilizer even under conditions where it grows and fixes N2 well. As a result, common bean is being generally considered as more responsive than other legumes to N fertilization (Graham, Citation1981). Silveira, Braz, Kliemann, and Zimmermann (Citation2005) indicated that large N inputs (more than 100 kg N ha−1 year−1) would be required to improve bean seed yields above 2,000 kg ha−1 year−1. Beans usually respond well to N fertilization up to 200 kg ha−1 (Anderson, Citation1974; Daba & Haile, Citation2002; Edje, Mughogho, & Ayonoadu, Citation1975; Mayona & Kamasho, Citation1988; Silveira et al., Citation2005). However, blanket fertilizer recommendations may be of limited relevance in smallholder farms, which experience a great heterogeneity in soil types and past managements (Zingore, Murwira, Delve, & Giller, Citation2007).
Inherent soil fertility which is highly diverse and heterogeneous in Africa affects the nutrient use efficiency, and consequently the yield of the crops (Giller, Rowe, de Ridder, & van Keulen, Citation2006). Wide variation in the response to fertilizers within farmers’ fields due to the existence of soil fertility gradients was recently observed in different areas of sub-Saharan Africa (Vanlauwe, Tittonell, & Mukalama, Citation2006; Wopereis et al., Citation2006; Zingore, Citation2011). Field experiments show that when soil fertility gradients are steep, nutrient resources, especially mineral fertilizers, are used most efficiently when applied on more fertile than on depleted soils (Wopereis et al., Citation2006; Zingore et al., Citation2007). This is because the depleted soils suffer multiple nutrient deficiencies and have poor physical structure. While it is known that common bean requires high levels of P, K, S, Ca, Mg, Mo, Co, and B (Franco & Day, Citation1980; Saito & Ruschel, Citation1978; Tsai, Bonetti, Agbala, & Rossetto, Citation1993), production is restricted mostly to marginal soils where sufficient nutrients are not available. Gates and Muller (Citation1979) observed that nodulation in soybean is affected by unbalanced nutritional conditions of N, P, and S, and that N2 fixation was not affected by mineral N only in presence of high levels of P and S. Tittonell, Zingore, van Wijk, Corbeels, and Giller (Citation2007) investigated that supply of balanced nutrients through manure application for maize improved the N and P use efficiency in Zimbabwe soils.
Lack of nutrients may restrict the development of a population of free-living rhizobia in the rhizosphere, limit the growth of the host plant, restrict nodulation itself, and cause impaired nodule function (Giller & Wilson, Citation1991). A sufficient number of appropriate rhizobia in the rhizosphere of legumes and adequate soil nutrients are a prerequisite for good nodulation and nitrogen fixation. Therefore, we hypothesized that fertile soil results in high agronomic efficiency of N (AE-N) by common bean than those on low fertile soil. And, the objective of this study was to determine the effect of inherent soil fertility on the agronomic efficiency of N, nodulation, and yield of common bean.
2. Materials and methods
2.1. Description of experimental sites
The experiment was established in the 2012/2013 cropping season under rain-fed condition at the eastern part of Ethiopia. Field experiments were conducted on four locations of Eastern Ethiopia, located in the Hirna, Fedis, Babillae, and Haramaya agricultural research centers. The soils had not been inoculated before with rhizobia strain nodulating common bean. Before sowing, soil samples were taken from a depth of 0 to 20 cm to determine baseline soil properties. Soil samples were air dried, crushed, and passed through a 2-mm sieve prior to physical and chemical analyses. Details of physical and chemical characteristics of the soil of experimental sites and geographical locations are presented in Table . Detailed description of all the soil analysis methods is given in the Soil Analysis Manual of Sahlemedhin and Taye (Citation2000).
Table 1. Soil analysis of experimental sites before sowing
2.2. Sources of seeds and Rhizobium strain
Common bean var. Dursitu were supplied by Lowland pulses research project, Haramaya University, Ethiopia. The variety was selected based on their yield, their maturity time, and recently improved released varieties of the experimental region. Strain of Rhizobium spp. (HUPvR-16) was obtained from Biofertilizer research and production project, Haramaya University (Haramaya, Ethiopia).
2.3. Inoculums preparation
Agar slope of strain of Rhizobium was supplied by Soil microbiology research laboratory, Haramaya Univeristy, Ethiopia. For purification of the isolate, the strain was preliminarily cultured in YEMA medium (10-g mannitol, 1-g yeast extract, 1-g KH2PO4, 0.1-g NaCl, and 0.2-g MgSO4.7H2O per liter, pH 6.8) and incubated at 28°C for five days. The pure colony of the isolate was later transferred to YEM broth medium with gentle shaking at 120 rpm for five days. By this procedure, the Rhizobium culture reached the middle or late logarithmic phase, and cell density in the culture was estimated by measuring optical density (540 nm). Rhizobium inoculant was prepared by mixing 30-g sterilized decomposed filter mud with 15-ml broth cultures of the appropriate Rhizobium strain in polyethylene bags. After incubating the inoculated filter mud for 2 weeks at 28°C, the count of the viable number of isolate was 1 × 109 g−1 inoculum. A population of Rhizobium in the inoculum was determined by duplicate plate counts (Vincent, Citation1970).
2.4. Experimental design and treatments
The initial indigenous rhizobia population was determined by the plant infection technique, using dilution of soil for nodulation tests according to the method of Brockwell (Citation1963). The MPN was calculated from the most likely number, using the MPN tables of Vincent (Citation1970). Field trials were conducted in order to investigate the effects of inherent soil fertility on the agronomic efficiency of N, nodulation, and yield of common bean. The experimental design was a spilt plot in randomized complete block design. Main plot treatments consisted of six N fertilizer rates: 0, 20, 40, 60, 80, and 100 kg N ha−1. Subplot treatments were two levels of inoculation with Rhizobium (inoculated and uninoculated). Treatments were randomized within blocks with three replicates per treatment. Nitrogen fertilizer in each level was divided into two equal parts; the first part of the N (20 kg N ha−1) was applied along the furrow by hand and incorporated before the planting time, and the remaining part was used in the flowering stages. In addition, 100-kg triple superphosphate ha−1 was applied manually as basal fertilizers in furrow at sowing.
The size of each experimental plot was 3 × 2 m2. There were five rows per plot and the spacing was 40 cm between rows, 10 cm between plants, 1 m between sub plots, and 1.5 between main plots. Seeds of common bean var. Dursitu were surface sterilized with calcium hypochlorite (6.7%) and then washed thoroughly in 10 changes of sterile distilled water. Disinfected seeds were inoculated (7-g inoculant per kg seed) containing HUPvR16 isolate after they were moistened with a 20% solution of sucrose and then inoculated. The area was moldboard-plowed and disked before planting. Inoculated seeds were hand planted on 7 July. Two seeds were sown per hill. After germination, the plants were thinned to one seedling per hill to obtain about 30 plants per row. Weeds were controlled over the growth period with hand hoeing.
2.5. Plant sampling and harvesting
At late flowering and early pod-setting stages, five plants from each plot were randomly chosen and harvested from central rows to record the number of nodule plant−1, nodule dry weight plant−1, and shoot dry weight plant−1. Shoots of the plants were dried at 70°C for 48hrs and later ground to pass a 0.5-cm sieve. Total N was determined using the Kjeldahl method of Bremner (Citation1965). At physiological maturity stage, on 30 October, yield and yield attributed of common bean were measured. Number of pod plant−1, number of seed plant−1, and plant height at harvest, dry biomass yield and grain yield (GY) (at 13% moisture content), and 100 seeds weight were properly determined. AE-N, a parameter representing the ability of the plant to increase yield in response to N applied, was calculated using the following formula (Fageria, Carvalho, Knupp, & Moraes, Citation2013).
where Gf is the GY of the N-fertilized plot, Gu is the GY of the unfertilized plot, and Na is the quantity of N applied.
2.6. Data analysis
The experiments were performed in a complete randomized block design with three replicates. Data were submitted to analysis of variance (SAS Institute, Citation1999). Statistically significant differences between means were also determined by the LSD test (SAS Institute, Citation1999).
3. Result and discussion
As indicated in Table , the soil of four experimental locations had similar amounts of Zn and B with higher SOM in Hirna soil followed by Fedis soil. The highest total and mineral N (
4
+ and
) and available P were measured at Hirna soil, while the lowest was obtained from Babillae soil. The present study also indicated a significant difference in AE-N (kg grain yield increase per kg N applied) in different experimental sites located in eastern part of Ethiopia with and without inoculation of Rhizobium leguminosarum bv. phaseoli (Table ). Along inoculated treatments, significantly higher AE-N (12.55 kg kg−1 N) was observed at Fedis site and the lowest AE-N of 7.36 kg kg−1 N was obtained from Hirna site. These AE-N are less than the average AE-N (23.17 kg kg−1 N) which was reported by Fageria, Melo, Ferreira, Oliveira, and Knupp (Citation2014). These various values of AE-N obtained from different locations are probably related with the difference in inherent soil fertility that prevailed in different study sites. Similarly, Minotta and Pinzauti (Citation1996) reported the highest nitrogen use efficiency of peach at low fertile soil when compared with those obtained from higher fertile soil. In contrast to the present study, the highest nutrient use efficiency was observed in fertile soil when compared to the less fertile soil (Wopereis et al., Citation2006; Zingore et al., Citation2007). The presence of higher total N and inorganic N (NH4 and NO3) in Hirna soil could be the cause of lower AE-N of common bean (Vanlauwe et al., Citation2011) beside the other soil-related limiting factors.
Table 2. Nodulation status, SDW, and agronomic efficiency of N of common bean var. Dursitu obtained from selected areas of Eastern Ethiopia
In uninoculated treatment, Babillae site had significantly higher AE-N (8.46 kg kg−1 N) than those measured from other experimental sites. The lowest AE-N was 5.47 kg kg−1 N at Haramaya site. Relatively lower total and mineral nitrogen at Babillae soil could help the plant to utilize efficiently the applied N fertilizer (Vanlauwe et al., Citation2011). Similarly, Jarrell and Beverly (Citation1981) indicated that plants grown at the lowest nutrient concentrations will inevitably have the highest utilization quotient because of dilution effects. Regardless of experimental sites, the highest AE-N was observed in Rhizobium inoculated than the uninoculated treatment. Production of plant growth hormones and solubilizing unavailable P by N-fixing bacteria (Cattelan, Hartel, & Fuhrmann, Citation1999; Chabot, Beauchamp, Kloepper, & Antoun, Citation1998) can increase root branching and root hair production (Vessey, Citation2003). This consequently enhances the uptake of plant nutrients including nitrate, potassium, and phosphorus (Lin, Okon, & Hardy, Citation1983).
Figure (a) shows a significant and polynomial association between the AE-N and nodule number per plant (NN) in all study locations, excluding Hirna site. A strong and concave downward association with highest coefficient of determination (R 2 = 0.442 at p ≤ 0.05) was determined at Haramaya site followed by Fedis site. This figure also indicates increase in AE-N with increase in NN up to 150 per plant, but the decrease in AE-N with increase in number of nodules beyond 150 was observed at Haramaya site. Abbasi, Manzoor, and Tahir (Citation2010) reported the direct correlation between the number of nodules production with that of grain and biomass yield of soybean though improving N derived from symbiotic N2 fixation. High fixer legume, for instance, soybean and lupin, needs less inorganic N than the less fixer legume (Common bean) (Giller, Citation2001).
Figure 1. Regression of agronomic efficiency of N of common bean var. Dursitu with (a) nodule number per plant, (b) nodule dry weight (NDW), (c) plant total tissue N (PTTN), and (d) grain yield (GY) over four representative locations of Eastern Ethiopia
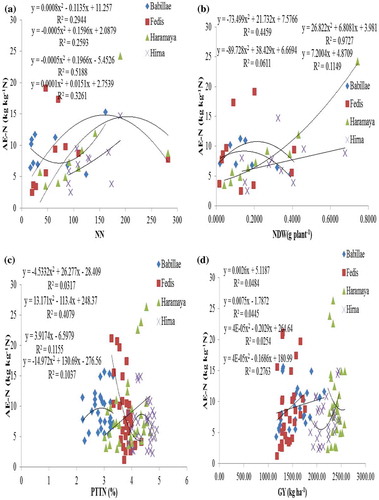
Significant and concave downward association was obtained from Babillae site with R 2 value of 0.445. A significant and concave upward association was observed at Babillae site with R 2 = 0.294 at p ≤ 0.05, indicating decrease in AE-N with increase in NN up to 150 number of nodules; above this value, the graph showed positive relationship between the NN and AE-N. This indicates the importance of good nodulation to improve the efficient utilization by common bean in Babillae site. Similarly, significant and polynomial association between AE-N and nodule dry weight (NDW) was also observed at Babillae and Haramaya sites but not at Fedis and Hirna sites (Figure (b)). At Haramaya site, strong and concave upward association was observed with highest R 2 = 0.854 at p ≤ 0.05. Beside this, the nodule produced at Haramaya and Fedis sites might not be effective in N fixation and thereby the crop becomes more responsive for applied N fertilizer application.
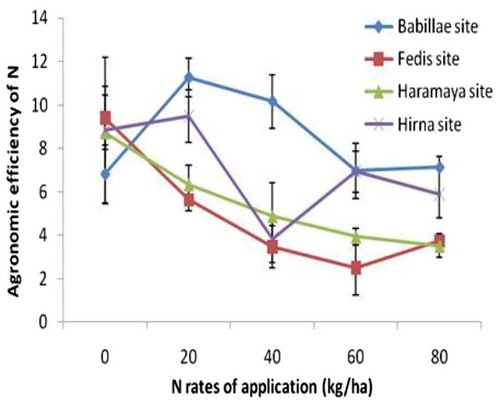
It is also noted that regardless the status of soil inherent fertility, AE-N decreased with increase in rates of N fertilizer application (Figure ). Similar trends for AE-N by common bean were observed (Fageria et al., Citation2014). Significant and negative association between AE-N and N rates of application was observed at all experimental locations, excluding Babillae site (Figure ). The highest coefficient of determination (R 2 = 0.498 at p < 0.05) was obtained from Haramaya site followed by Fedis and Hirna sites. A decrease in N use efficiency with increase in rates of N application has been previously reported by Salvagiotti et al. (Citation2008). Slight increase in AE-N at 20-kg N ha−1 was observed at Babillae and Hirna sites (Figure ). Lower N at Babillae site and higher available P at Hirna site could increases the yield of common bean at 20-kg N ha−1 application (Leidi & Rodríguez-Navarro, Citation2000), thereby improving AE-N.
Figure 2. Regression of agronomic efficiency of N of common bean var. Dursitu on N rates of application over different representative locations of Eastern Ethiopia
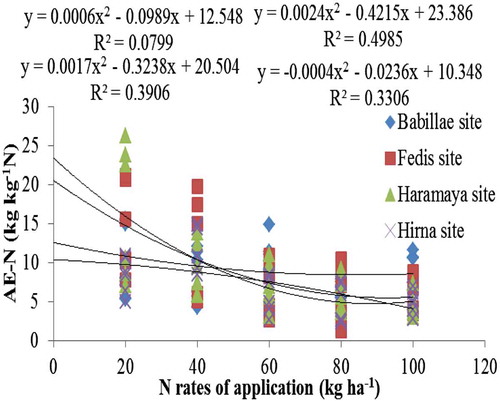
Figure 3. Line graph of agronomic efficiency of N of common bean var. Dursitu along different rates of N application
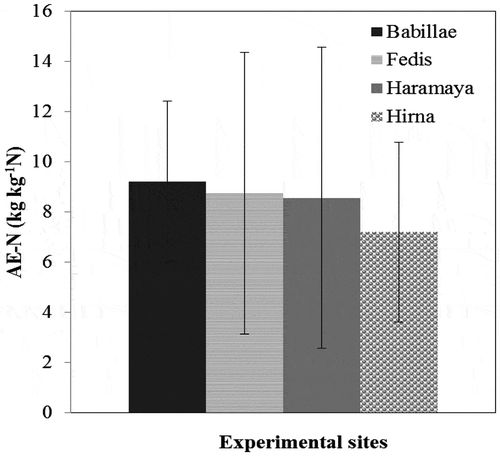
The highest overall mean of AE-N was calculated in Babillae site followed by Fedis site, while Hirna site scored the lowest AE-N (Figure ). The highest AE-N of common bean at Fedis and Haramaya was calculated at the control treatment (i.e. without inoculation and N application). Higher soil N with low P in these soils is probably the reason for non-responsiveness of common bean for inorganic N application (Vanlauwe et al., Citation2011; Zingore, Murwira, Delve, & Giller, Citation2008). Nodulation process and N2 fixation of common bean are highly sensitive and suppressed by external supply of N (Abaidoo & van Kessel, Citation1989; George, Singleton, & Bohlool, Citation1988), which may consequently reduce the yield of common bean.
Figure 4. Bar graph of agronomic efficiency of N of common bean var. Dursitu in different experimental locations
Common bean var. Dursitu cultivated in the eastern part of Ethiopia revealed significant difference in NN obtained from four experimental sites (Table ). The highest NN (131.67) and NDW (0.4959 g) was measured from Hirna site from Rhizobium inoculated and uninoculated treatments, respectively. This indicates the presence of high and competitive native rhizobia nodulating common bean in this soil. The result of the present experiment also indicated conducive soil fertility conditions (higher available P, CEC and SOM, and total Nitrogen) at Hirna site, which are probably suited for higher nodule formation of common bean. Similarly, Tsai et al. (Citation1993) reported that average nodule number and weight increased under high fertility levels. Olivera, Tejera, Iribarne, Ocana, and Lluch (Citation2004) indicated that P supply increased the nodule number and NDW. In contrast, Leidi and Rodríguez-Navarro (Citation2000) found that at high nitrate concentration, nodulation and N2 fixation did not improve with increase in P supply. We also found that significantly lower NN was measured from Fedis site as compared to those obtained from Haramaya and Hirna sites. This could be due to the fact that this soil had higher soil N which might inhibit the nodulation development (Abaidoo & van Kessel, Citation1989; George et al., Citation1988).
Regardless of inoculation treatment, the lowest NN was obtained from Babillae site. Low soil fertility, especially low available P and soil organic matter in Babillae soil, could reduce the survival of native rhizobia in the soil, thereby inhibiting the nodule formation (Giller & Wilson, Citation1991). Israel (Citation1987) determined that common bean nodule formation is more sensitive than plant growth for P deficiency. Sulieman, Abdalla, Omer, and Hago (Citation2009) found that the highest nodule formation was observed in higher P application with organic matter incorporation compared to those obtained from low P level without organic matter amendment. However, Musiyiwa, Mpepereki, and Giller (Citation2005) found the poor correlation between rhizobial population in soil with soil pH, CEC, P, N, and organic carbon.
A significant variation of shoot dry weight (SDW) measured at late flowering stage was obtained from four experimental locations, regardless of inoculation treatments (Table ). In both inoculation treatments, the highest mean SDW (80.58 g) was observed at Haramaya site followed by Hirna site. Better soil fertility status and good nodulation development may favor for higher biomass accumulation of common bean at late flowering stage (Pacheco, Brito, Straliotto, Pérez, & Araújo, Citation2012). Chemining’wa, Theuri, and Muthomi (Citation2011) found the positive and significant correlation between the nodule number per plant and plant biomass production. However, Hungria et al. (Citation2000) found that increases in nodulation did not result in significant yield increases. The lowest SDW was measured at Babillae site which was 44.46 and 32.54% lower when compared to those of the highest SDW obtained from Haramaya site at inoculated and uninoculated treatments, respectively. The lower SDW at Babillae site could be related with lower N derived from symbiotic N2 fixation due to low nodulation besides low inherent soil fertility that prevailed in the experimental site (Tsai et al., Citation1993).
Number of pods per plant is one of the important yield component in determining GY of common been. A significant difference in NPP obtained from different experiments sites at p ≤ 0.05 was observed (Table ). The highest NPP was measured at Hirna and Haramaya sites with inoculated treatment. In uninoculated treatment, significantly higher NPP was observed at Haramaya site. The native soil rhizobia nodulating common bean could be higher and effective in N2 fixation at Haramaya site and thus improved NPP. The highest mean NPP was observed at inoculated treatment in comparison to those NPP obtained from uninoculated treatment, regardless of experimental locations. Similar result was reported by Salinas-Ramírez, Escalante-Estrada, Rodríguez-González, and Sosa-Montes (Citation2011) who found that inorganic N achieved significant improvement of pod numbers of Snap bean in Mexico when it was applied with Rhizobium sp. inoculation. They also hypothesized that the application of inoculation could stimulate root growth, thus favoring the absorption of more nutrients and water, and thereby increasing the photosynthesis and improving the productivity of common bean. Regardless of inoculation treatments, the lowest mean NPP was observed at Babillae site. This is probably related with lower native soil fertility that prevailed at the study site (Rys & Bonish, Citation1981). Low soil fertility might have been the cause of insufficient and non-effective rhizobia of common bean harboring in the soil (Chemining’wa & Vessey, Citation2006).
Table 3. Number of pods per plant, NSP, and 100 seeds weight of common bean var. Dursitu obtained from selected areas of Eastern Ethiopia
The ANOVA showed the significant variation of number of seeds per pod (NSP) obtained from different experimental locations at p ≤ 0.05 (Table ). In Rhizobium inoculated treatment, significantly lower NSP was observed at Fedis site. The early termination of rainfall at this experimental site could affect the nodule formation by the host plant (Javaheri, Citation1996), consequently reducing the yield and yield traits of common bean. In uninoculated treatment, the data indicated non-significant difference of NSP obtained from four experimental locations. Similar finding was also reported by Karasu, Oz, and Dogan (Citation2011) on the same host, regardless of inoculation and nitrogen application. This could be due to the fact that NSP is controlled by genetic makeup of the plant rather than the external environmental factors (Fageria & Santos, Citation2008).
Table indicates a significant difference in 100 seeds weight measured from four experimental sites at p ≤ 0.05. In both inoculation treatments, the highest 100 seeds weight was produced at Fedis site, while the lowest 100 seeds weight was observed at Hirna site. Similarly, there was also a significant difference in total biomass yield (TBY) which was produced at four different experimental sites (Table ). Regardless of Rhizobium inoculated treatment, the highest TBY was obtained from Hirna site followed by Haramaya site. The maximum TBY production (7,211.4 kg ha−1) obtained from Hirna site was 61% higher than that measured at Babillae site. Similarly, Pacheco et al. (Citation2012) found that higher biomass production of common bean was observed in soils having better soil fertility than those obtained from soils having less fertility. This could also be related with an improvement in N derived from symbiotic N2 fixation at higher fertile soil (Carpena et al., Citation2000; Giller, Citation2001).
Table 4. Total biomass, GY, HI, and plant total tissue N of common bean var. Dursitu obtained from selected areas of Eastern Ethiopia
It is also noted that a significant difference of GY was obtained from four experimental locations at p ≤ 0.05 (Table ). Similarly, the effect of inherent soil fertility on the productivity of common bean has been previously reported by Pacheco et al. (Citation2012). In Rhizobium inoculated treatment, significantly higher GY was observed at Haramaya followed by Hirna site. This indicates that the rhizobia nodulating common bean at Haramaya site may have been effective in N2 fixation than those that harbored other experimental sites. Beside this, Fageria and Baligar (Citation1997) found that phosphorus followed by calcium, magnesium, boron, and zinc is the most limited nutrient for common bean. The highest available P and higher CEC at Haramaya and Hirna sites could promote the uptake of N (Zhou, Liang, Chen, Shen, & Jing, Citation2006), thus stimulating photosynthesis (Seemann, Sharkey, Wang, & Osmond, Citation1987) and leading to higher GYs. They also found higher photosynthetic performance of soybean in rhizobia inoculation compared to N-limited plants, as we observed in the present study. Similarly, improvement of cation nutrients including CEC resulted in increase in the yield of common bean (Fageria, dos Santos, & Moreira, Citation2010). Rondon, Lehmann, Ramírez, and Hurtado (Citation2007) found that biochar application improved P, K, Mg, Ca, Mo, and B availability, and thus improved common bean production. Moreover, owing to the fact that for N2 fixation, considerably higher levels of P are required (Olivera et al., Citation2004). Increased mineral nutrient levels in the plants would not only benefit Rhizobium directly, but will also lead to increase in photosynthesis, making a greater proportion of photosynthates available to the Rhizobium nodules (Harris, Pacovsky, & Paul, Citation1985; Mortimer, Pérez-Fernández, & Valentine, Citation2008). The highest mean GY (2,355.51 kg ha−1) was produced at Haramaya site with inoculation of Rhizobium sp. The lowest was 1,513.42 kg ha−1 obtained from Fedis without Rhizobium sp inoculation. The lowest GY produced at this study is higher than that previously reported by Singh et al. (Citation2003), who found that the highest GY obtained from higher fertile soil was <1,313 kg ha−1. This may be probably related with low P uptake and utilization due to the semi-arid weather in the study site (Al-Karaki, Clark, & Sullivan, Citation1995). Lynch, Läuchli, and Epstein (Citation1991) found that P deficiency primarily reduced leaf area by diminishing the number of leaves through effects on the number of nodes, branching, and relative leaf appearance rate, and secondarily by reducing leaf expansion.
Harvest index (HI) is an important trait associated with the dramatic increases in crop yields (Sinclair, Citation1998). The result of the experiment revealed a significant variation of HI of common bean var. Dursitu at p ≤ 0.05 (Table ). The highest mean value of HI (0.63) was calculated from Babillae site, while the lowest mean HI (0.29) was observed at Hirna site. Similar finding was observed on common bean by Araújo, Teixeira, and Almeida (Citation2000). They reported that vegetative growth at higher P supply was greater than the GY improvement, thus reducing the harvesting index of common bean.
It is also observed that there was a significant variation of plant total tissue N (PTTN) obtained from four experimental locations (Table ). Regardless of inoculation treatments, the highest PTTN was determined from Hirna site followed by Haramaya site. 4.2672 and 4.5050% were the highest mean PTTNs analyzed from Rhizobium inoculated and uninoculated treatments, respectively, from Hirna site. This indicates the presence of relatively balanced plant nutrients which enhance N uptake from the soil and improve the symbiotic N2 fixation at Hirna site (Wang, Pan, Chen, Yan, & Liao, Citation2011; Yanni, Citation1992). The lowest mean PTTN was 2.8183 and 2.7844% obtained from inoculated and uninoculated treatments, respectively, from Babillae site. Lower native soil N and the presence of low soil fertility probably reduced the N derived from symbiotic N2 fixation, thereby reducing plant accumulated N (Leidi & Rodríguez-Navarro, Citation2000).
4. Conclusions
The result of this study, therefore, indicated the significant effect of inherent soil fertility on the use efficiency of N by common bean. This study indicated the remarkable effect of soil total N and inherent soil fertility on the use efficiency of applied inorganic N by common bean. The highest AE-N of common bean was observed in soil having fertile soil with moderate N content, while the lowest AE-N was observed in soil having high soil fertility with high N content. Moreover, the effectiveness of the native rhizobia harboring different study sites might have been varied due to different soil fertility status which may also affect the AE-N. This study also indicated the significant effect of the inherent soil fertility on the common bean production in the study sites. Therefore, we conclude that the effectiveness of native rhizobia nodulating common bean in addition to the inherent soil fertility should be considered when we recommend inorganic N for common bean.
Additional information
Funding
Notes on contributors
Anteneh Argaw
Anteneh Argaw is a MSc research and teaching scholar at Haramaya University, Ethiopia, employed as an assistant professor since 2009. He completed his MSc (Applied Microbiology) in 2007 from Addis Ababa University and BSc (Plant Sciences) in 2001 from Alemaya University, Ethiopia. His research interests lay in the characterization, selection, and use of elite isolates of Rhizobium sp. nodulating different food legumes and development of best integrated soil fertility management for different crops grown in different farming systems. Daniel Muleta is a PhD student in Adeliade University, Australia, since 2012. He holds a MSc (Applied Microbiology) degree from Addis Ababa University and BSc (Biology) degree from Alemaya University. He has been working in Ethiopia Institute of Agricultural Research as an associate researcher in soil microbiology and microbial biotechnology research project. His research interest lay in the isolation, characterization, and selection of acid-tolerant rhizobia nodulating legume crop and development of plant growth promoting rhizosphereic bacteria tolerant to acidic soils.
References
- Abaidoo, R. C. , & van Kessel, C. (1989). 15N-uptake, N2-fixation and rhizobial interstrain competition in soybean and bean, intercropped with maize. Soil Biology and Biochemistry , 21 , 155–159.10.1016/0038-0717(89)90025-4
- Abbasi, M. K. , Manzoor, M. , & Tahir, M. M. (2010). Efficiency of rhizobium inoculation and P fertilization in enhancing nodulation, seed yield, and phosphorus use efficiency by field grown soybean under hilly region of Rawalakot Azad Jammu and Kashmir, Pakistan. Journal of Plant Nutrition , 33 , 1080–1102.10.1080/01904161003729782
- Al-Karaki, G. N. , Clark, R. B. , & Sullivan, C. Y. (1995). Effects of phosphorus and water stress levels on growth and phosphorus uptake of bean and sorghum cultivars. Journal of Plant Nutrition , 18 , 563–578.10.1080/01904169509364923
- Anderson, G. D. (1974). Bean responses to fertilizers on Mt Kilimanjaro in relation to soil and climatic conditions. East African Agricultural and Forestry Journal , 39 , 272–288.
- Araújo, A. P. , Teixeira, M. G. , & Almeida, D. L. D. (2000). Growth and yield of common bean cultivars at two soil phosphorus levels under biological nitrogen fixation. Pesquisa Agropecuária Brasileira , 35 , 809–817.10.1590/S0100-204X2000000400019
- Bliss, F. A. (1993). Utilizing the potential for increased nitrogen fixation in common bean. Plant and Soil , 152 , 157–160.10.1007/BF00016346
- Bremner, J. M. (1965). Inorganic forms of nitrogen. In C. A. Black , D. D. Evans , L. E. Ensminger , J. L. White , & F. E. Clark (Eds.), Methods of soil analysis (Part 2, 2nd ed., Agronomy Monograph, No. 9). Madison, WI: ASA and SSSA.
- Brockwell, J. (1963). Accuracy of a plant-infection technique for counting populations of Rhizobium trifolii . Applied Microbiology , 2 , 377–383.
- Buttery, B. R. , Park, S. J. , & Findlay, W. I. (1987). Growth and yield of white bean (Phaseolus vulgaris L.) in response to nitrogen, phosphorus and potassium fertilizer and to inoculation with rhizobium. Canadian Journal of Plant Science , 67 , 425–432.10.4141/cjps87-061
- Carpena, R. , Esteban, E. , Sarro, M. , Peñalosa, J. , Gárate, A. , Lucena, J. , & Zornoza, P. (2000). Boron and calcium distribution in nitrogen fixing pea plants. Plant Science , 151 , 163–170.10.1016/S0168-9452(99)00210-1
- Cattelan, A. J. , Hartel, P. G. , & Fuhrmann, J. J. (1999). Screening for plant growth-promoting rhizobacteria to promote early soybean growth. Soil Science Society of America Journal , 63 , 1670–1680.10.2136/sssaj1999.6361670x
- Central Statistical Authority . (2013). Report on area and production for major crops in 2012/2013 . Addis Ababa: Author.
- Centro Internacional de Agricultura Tropical . (2001). Plant genetic resources: Beans . Retrieved from http://www.ciat.cgiar.org/pgr/beans.htm
- Chabot, R. , Beauchamp, C. J. , Kloepper, J. W. , & Antoun, H. (1998). Effect of phosphorus on root colonization and growth promotion of maize by bioluminescent mutants of phosphate-solubilizing Rhizobium leguminosarum biovar phaseoli . Soil Biology and Biochemistry , 30 , 1615–1618.10.1016/S0038-0717(98)00054-6
- Chemining’wa, G. N. , Theuri, S. W. M. , & Muthomi, J. W. (2011). Abundance of indigenous rhizobia nodulating cowpea and common bean in Central Kenyan soils. African Journal of Horticultural Science , 5 , 92–97.
- Chemining’wa, G. N. , & Vessey, J. K. , (2006). The abundance and efficacy of Rhizobium leguminosarum bv. viciae in cultivated soils of the eastern Canadian prairie. Soil Biology and Biochemistry , 38 , 294–302.10.1016/j.soilbio.2005.05.007
- Daba, S. , & Haile, M. (2002). Effects of rhizobial inoculant and nitrogen fertilizer on yield and nodulation of common bean under intercropped conditions. Journal of Plant Nutrition , 25 , 1443–1455.10.1081/PLN-120005401
- Edje, O. T. , Mughogho, L. K. , & Ayonoadu, U. W. U. (1975). Responses of dry beans to varying nitrogen levels. Agronomy Journal , 67 , 251–255.10.2134/agronj1975.00021962006700020020x
- Fageria, N. K. , & Baligar, V. C. (1997). Response of common bean, upland rice, corn, wheat, and soybean to soil fertility of an oxisol. Journal of Plant Nutrition , 20 , 1279–1289.10.1080/01904169709365335
- Fageria, N. K. , Carvalho, M. C. S. , Knupp, A. M. , & Moraes, M. F. (2013). Nutrient uptake and use efficiency of dry bean in tropical lowland soil. Communications in Soil Science and Plant Analysis , 44 , 2852–2859.10.1080/00103624.2013.823982
- Fageria, N. K. , dos Santos, A. B. , & Moreira, A. (2010). Yield, nutrient uptake, and changes in soil chemical properties as influenced by liming and iron application in common bean in a no-tillage system. Communications in Soil Science and Plant Analysis , 41 , 1740–1749.10.1080/00103624.2010.489137
- Fageria, N. K. , Melo, L. C. , Ferreira, E. P. B. , Oliveira, J. P. , & Knupp, A. M. (2014). Dry matter, grain yield, and yield components of dry bean as influenced by nitrogen fertilization and rhizobia. Communications in Soil Science and Plant Analysis , 45 , 111–125.10.1080/00103624.2013.848877
- Fageria, N. K. , & Santos, A. B. (2008). Yield physiology of dry bean. Journal of Plant Nutrition , 31 , 983–1004.10.1080/01904160802096815
- Franco, A. A. , & Day, J. M. (1980). Effects of lime and molybdenum on nodulation and nitrogen fixation of Phaseolus vulgaris in acid soils of Brazil. Turrialba , 30 , 99–105.
- Gates, C. T. , & Muller, W. J. (1979). Nodule and plant development in the soyabean, Glycine max (L.) Merr.: Growth response to nitrogen, phosphorus and sulfur. Australian Journal of Botany , 27 , 203–215.10.1071/BT9790203
- George, T. , Singleton, P. W. , & Bohlool, B. B. (1988). Yield, soil nitrogen uptake, and nitrogen fixation by soybean from four maturity groups grown at three elevations. Agronomy Journal , 80 , 563–567.10.2134/agronj1988.00021962008000040004x
- Giller, K. E. (2001). Nitrogen fixation in tropical cropping systems (2nd ed.). New York, NY: CABI Publishing.10.1079/9780851994178.0000
- Giller, K. E. , Rowe, E. , de Ridder, N. , & van Keulen, H. (2006). Resource use dynamics and interactions in the tropics: Scaling up in space and time. Agricultural Systems , 88 , 8–27.10.1016/j.agsy.2005.06.016
- Giller, K. E. , & Wilson, K. J. (1991). Nitrogen fixation in tropical cropping systems . Wallingford: CAB International.
- Graham, P. H. (1981). Some problems of nodulation and symbiotic nitrogen fixation in Phaseolus vulgaris L.: A review. Field Crops Research , 4 , 93–112.10.1016/0378-4290(81)90060-5
- Graham, P. H. , Viteri, S. E. , Mackie, F. , Vargas, A. T. , & Palacios, A. (1982). Variation in acid soil tolerance among strains of Rhizobium phaseoli . Field Crops Research , 5 , 121–128.10.1016/0378-4290(82)90012-0
- Harris, D. , Pacovsky, R. S. , & Paul, E. A. (1985). Carbon economy of soybean–rhizobium–glomus associations. New Phytologist , 101 , 427–440.10.1111/nph.1985.101.issue-3
- Hungria, M. , Andrade, D. S. , Chueire, L. M. D. O. , Probanza, A. , Guttierrez-Mañero, F. J. , & Megı́as, M. (2000). Isolation and characterization of new efficient and competitive bean (Phaseolus vulgaris L.) rhizobia from Brazil. Soil Biology and Biochemistry , 32 , 1515–1528.10.1016/S0038-0717(00)00063-8
- Huntington, T. G. , Smith, M. S. , Thomas, G. W. , & Blevins, R. L. (1986). Response of Phaseolus vulgaris to inoculation with Rhizobium phaseoli under two tillage systems in the Dominican Republic. Plant and Soil , 95 , 77–85.10.1007/BF02378854
- Israel, D. W. (1987). Investigation of the role of phosphorus in symbiotic dinitrogen fixation. Plant Physiology , 84 , 835–840.10.1104/pp.84.3.835
- Jarrell, W. M. , & Beverly, R. B. (1981). The dilution effect in plant nutrition studies. Advanced Agronomy , 34 , 197–224.10.1016/S0065-2113(08)60887-1
- Javaheri, F. (1996). Naturally nodulating (promiscuous) soybean varieties for smallholder farmers: The Zambian experience. In S. Mpepereki , F. Makonese , & K. E. Giller (Eds.), Soybeans in smalleholder cropping systems of Zimbabwe (pp. 63–73). Harare: SoilFerNet/CIMMYT.
- Karasu, A. , Oz, M. , & Dogan, R. (2011). The effect of bacterial inoculation and different nitrogen doses on yield and yield components of some dwarf dry bean cultivars (Phaseolus vulgaris L.). Bulgarian Journal of Agricultural Science , 17 , 296–305.
- Leidi, E. , & Rodríguez-Navarro, D. N. (2000). Nitrogen and phosphorus availability limit N2 fixation in bean. New Phytologist , 147 , 337–346.10.1046/j.1469-8137.2000.00703.x
- Lin, W. , Okon, Y. , & Hardy, R. W. F. (1983). Enhanced mineral uptake by Zea mays and Sorghum bicolor roots inoculated with Azospirillum brasilense . Applied and Environmental Microbiology , 45 , 1775–1779.
- Lynch, J. , Läuchli, A. , & Epstein, E. (1991). Vegetative growth of the common bean in response to phosphorus nutrition. Crop Science , 31 , 380–387.10.2135/cropsci1991.0011183X003100020031x
- Mayona, C. M. , & Kamasho, J. (1988). Research experiences with inorganic and organic fertilizers in the southern highlands of Tanzania. In C. S. Wortmann (Ed.), Proceedings of a workshop on soil fertility research for bean cropping systems in Africa ( CIAT African Workshop Series No. 3). Addis Ababa: CIAT.
- Minotta, G. , & Pinzauti, S. (1996). Effects of light and soil fertility on growth, leaf chlorophyll content and nutrient use efficiency of beech (Fagus sylvatica L.) seedlings. Forest Ecology and Management , 86 , 61–71.10.1016/S0378-1127(96)03796-6
- Mortimer, P. E. , Pérez-Fernández, M. A. , & Valentine, A. J. (2008). The role of arbuscular mycorrhizal colonization in the carbon and nutrient economy of the tripartite symbiosis with nodulated Phaseolus vulgaris . Soil Biology and Biochemistry , 40 , 1019–1027.10.1016/j.soilbio.2007.11.014
- Moxley, J. C. , Hume, D. J. , & Smith, D. L. (1986). N2 fixation and competitiveness of Rhizobium phaseoli strains isolated from Ontario soils. Canadian Journal of Plant Science , 66 , 825–836.10.4141/cjps86-104
- Musiyiwa, K. , Mpepereki, S. , & Giller, K. E. (2005). Symbiotic effectiveness and host ranges of indigenous rhizobia nodulating promiscuous soyabean varieties in Zimbabwean soils. Soil Biology and Biochemistry , 37 , 1169–1176.10.1016/j.soilbio.2004.12.004
- Olivera, M. , Tejera, N. , Iribarne, C. , Ocana, A. , & Lluch, C. (2004). Growth, nitrogen fixation and ammonium assimilation in common bean (Phaseolus vulgaris): Effect of phosphorus. Physiologia Plantarum , 121 , 498–505.10.1111/ppl.2004.121.issue-3
- Pacheco, R. S. , Brito, L. F. , Straliotto, R. , Pérez, D. , & Araújo, A. P. (2012). Seeds enriched with phosphorus and molybdenum as a strategy for improving grain yield of common bean crop. Field Crops Research , 136 , 97–106.10.1016/j.fcr.2012.07.017
- Piha, M. I. , & Munns, D. N. (1987). Nitrogen fixation capacity of field-grown bean compared to other grain legumes. Agronomy Journal , 79 , 690–696.10.2134/agronj1987.00021962007900040022x
- Redden, R. J. , & Herridge, D. F. (1999). Evaluation of genotypes of navy and culinary bean (Phaseolus vulgaris L.) selected for superior growth and nitrogen fixation. Australian Journal of Experimental Agriculture , 39 , 975–980.10.1071/EA99108
- Rondon, M. A. , Lehmann, J. , Ramírez, J. , & Hurtado, M. (2007). Biological nitrogen fixation by common beans (Phaseolus vulgaris L.) increases with bio-char additions. Biology and Fertility of Soils , 43 , 699–708.10.1007/s00374-006-0152-z
- Rys, G. J. , & Bonish, P. M. (1981). Effectiveness of Rhizobium trifolii populations associated with Trifolium species in Taranaki, New Zealand. New Zealand Journal of Experimental Agriculture , 9 , 329–335.10.1080/03015521.1981.10425430
- Sahlemedhin, S. , & Taye, B. (2000). Procedures for soil and plant analysis ( EARO, Technical Paper No. 74). Addis Ababa: National Soil Research Center.
- Saito, S. M. T. , & Ruschel, A. P. (1978). Influence of lime, phosphate and micronutrients on the natural nodulation of beans (Phaseolus vulgaris L.). Anais de ESALQ , 35 , 545–556.
- Salinas-Ramírez, N. , Escalante-Estrada, J. A. , Rodríguez-González, M. T. , & Sosa-Montes, E. (2011). Yield and nutritional quality of snap bean in terms of biofertilization. Tropical and Subtropical Agroecosystem , 13 , 347–355.
- Salvagiotti, F. , Cassman, K. G. , Specht, J. E. , Walters, D. T. , Weiss, A. , & Dobermann, A. (2008). Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Research , 108 , 1–13.10.1016/j.fcr.2008.03.001
- Sanchez, P. A. (1976). Properties and management of soil in the tropics . New York, NY: Wiley.
- SAS Institute . (1999). SAS/GRAPH Software (Version 8). Cary, NC: Author.
- Seemann, J. S. , Sharkey, T. D. , Wang, J. L. , & Osmond, C. B. (1987). Environmental effects on photosynthesis, nitrogen-use efficiency, and metabolite pools in leaves of sun and shade plants. Plant Physiology , 84 , 796–802.10.1104/pp.84.3.796
- Silveira, P. M. , Braz, A. J. B. P. , Kliemann, H. J. , & Zimmermann, F. J. P. (2005). Nitrogen fertilization of common bean grown under no-tillage system after several cover crops. Pesquisa Agropecuária Brasileira , 40 , 377–381.10.1590/S0100-204X2005000400009
- Sinclair, T. R. (1998). Historical changes in harvest index and crop nitrogen accumulation. Crop Science , 38 , 638–643.
- Singh, S. P. (1999). Production and utilization. In S. P. Singh (Ed.), Common bean improvement in the twenty-first century (pp. 1–24). Dordrecht: Kluwer.10.1007/978-94-015-9211-6
- Singh, S. P. , Terán, H. , Muñoz, C. G. , Osorno, J. M. , Takegami, J. C. , & Thung, M. D. T. (2003). Low soil fertility tolerance in landraces and improved common bean genotypes. Crop Science , 43 , 110–119.10.2135/cropsci2003.0110
- Sulieman, S. A. , Abdalla, M. A. , Omer, E. T. A. , & Hago, T. E. M. (2009). Phosphorus supply and Phaseolus vulgaris performance grown in Shambat clay alkaline soil and influenced by farmyard manure. Australian Journal of Basic Applied Science , 3 , 2598–2606.
- Thung, M. , & Rao, I. M. (1999). Integrated management of abiotic stresses. In S. P. Singh (Ed.), Common bean improvement in the twenty-first century (pp. 331–370). Dordrecht: Kluwer.10.1007/978-94-015-9211-6
- Tittonell, P. , Zingore, S. , van Wijk, M. T. , Corbeels, M. , & Giller, K. E. (2007). Nutrient use efficiencies and crop responses to N, P and manure applications in Zimbabwean soils: Exploring management strategies across soil fertility gradients. Field Crops Research , 100 , 348–368.10.1016/j.fcr.2006.09.003
- Tsai, S. M. , Bonetti, R. , Agbala, S. M. , & Rossetto, R. (1993). Minimizing the effect of mineral nitrogen on biological nitrogen fixation in common bean by increasing nutrient levels. Plant and Soil , 152 , 131–138.10.1007/BF00016342
- Van Rheenen, H. A. (1982). Report from the National Dry Bean Council Research Conference . Malawi: Gainstville.
- Vanlauwe, B. , Kihara, J. , Chivenge, P. , Pypers, P. , Coe, R. , & Six, J. (2011). Agronomic use efficiency of N fertilizer in maize-based systems in sub-Saharan Africa within the context of integrated soil fertility management. Plant and Soil , 339 , 35–50.10.1007/s11104-010-0462-7
- Vanlauwe, B. , Tittonell, P. , & Mukalama, J. (2006). Within-farm soil fertility gradients affect response of maize to fertilizer application in western Kenya. Nutrient Cycling in Agroecosystems , 76 , 171–182.
- Vessey, J. K. (2003). Plant growth promoting rhizobacteria as biofertilizers. Plant and Soil , 255 , 571–586.10.1023/A:1026037216893
- Vincent, J. M. (1970). A manual for the practical study of root-nodule bacteria ( IBP Handbook No. 15). Oxford: Blackwell.
- Wang, X. , Pan, Q. , Chen, F. , Yan, X. , & Liao, H. (2011). Effects of co-inoculation with arbuscular mycorrhizal fungi and rhizobia on soybean growth as related to root architecture and availability of N and P. Mycorrhiza , 21 , 173–181.10.1007/s00572-010-0319-1
- Wopereis, M. C. S. , Tamélokpo, A. , Ezui, K. , Gnakpénou, D. , Fofana, B. , & Breman, H. (2006). Mineral fertilizer management of maize on farmer fields differing in organic inputs in the West African savanna. Field Crops Research , 96 , 355–362.10.1016/j.fcr.2005.08.004
- Wortmann, C. S. (2001). Nutrient dynamics in a climbing bean and sorghum crop rotation in the Central Africa highlands. Nutrient Cycling in Agroecosystems , 61 , 267–272.10.1023/A:1013776514526
- Wortmann, C. S. , Kirkby, R. A. , Eledu, C. A. , & Allen, D. J. (1998). Atlas of common bean (Phaseolus vulgaris L.) production in Africa ( CIAT Publication No. 293, 133 pp.). Cali: CIAT Pan-African Bean Research Alliance.
- Yanni, Y. G. (1992). The effect of cyanobacteria and azolla on the performance of rice under different levels of fertilizer nitrogen. World Journal of Microbiology & Biotechnology , 8 , 132–136.10.1007/BF01195832
- Zhou, X. J. , Liang, Y. , Chen, H. , Shen, S. H. , & Jing, Y. X. (2006). Effects of rhizobia inoculation and nitrogen fertilization on photosynthetic physiology of soybean. Photosynthetica , 44 , 530–535.10.1007/s11099-006-0066-x
- Zingore, S. (2011). Maize productivity and response to fertilizer use as affected by soil fertility variability, manure application, and cropping system. Better Crops , 95 , 4–6.
- Zingore, S. , Murwira, H. K. , Delve, R. J. , & Giller, K. E. (2007). Influence of nutrient management strategies on variability of soil fertility, crop yields and nutrient balances on smallholder farms in Zimbabwe. Agriculture, Ecosystems & Environment , 119 , 112–126.
- Zingore, S. , Murwira, H. K. , Delve, R. J. , & Giller, K. E. (2008). Variable grain legume yields, responses to phosphorus and rotational effects on maize across soil fertility gradients on African smallholder farms. Nutrient Cycling in Agroecosystems , 80 , 1–18.10.1007/s10705-007-9117-3