
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
A new group of colored shade net has been developed that can influence the quality and quantity of the light intercepted by the plants. To explore the effect of applying colored shade nets on physiological traits of marigold and violet plants, a split-plot experiment was carried out. The trial was done with two factors including colored shade net at four levels (no net, green, yellow and red net) and two plant species (marigold and violet). The results showed that yellow shade net increased the vegetative growth of the plants. However, the highest leaf area was related to the red shade net. Chlorophyll content, Brix°, and carotenoid content were significantly higher in plants grown under the yellow net. The highest peroxidase was obtained from green and red shade nets. The plants grown under the green shade net had higher anthocyanin and antioxidant activities than other shade nets. The interaction of “shade net × plant species” showed that the highest root and flower fresh weights were related to “yellow net × marigold”, the highest plant growth rate, total chlorophyll, and carotenoid contents was related to “yellow net × violet”. The results revealed that yellow shade nets outperformed the other nets in improving plant growth.
PUBLIC INTEREST STATEMENT
Photoselective shade nets can selectively filter the incoming solar radiation and protect the plants. This technology is based on plastic shade net products during whose production the chromophores and different light scattering and reflection elements are combined. We studied in this research, the physiological effects of these nets on the pigments, antioxidant capacity, and growth of two ornamental plants.
Competing Interests
The authors declare no competing interests.
1. Introduction
Iran is located in a region that is rated high in terms of incoming solar energy in the world. Solar radiation rate in Iran is estimated at 1800–220 kW ha−1 m−1 yr−1, which is higher than the global average (Safaii et al., Citation2005). Iran has, on average, 280 sunny days during the year, which is considerable; however, it makes it unavoidable to resort a shading to control the sunlight (Forghani & Kiani Abri, Citation2005).
The application of photoselective shade nets in crop greenhouse technology dates back to the early 2000s in the Mediterranean region. Shahak, Gal et al. (Citation2008) first concluded that colored shade nets, including red, yellow, blue, gray, and pearl, were more effective on orchard yields. Photoselective shade nets for plant cover refer to nets that can selectively filter the incoming solar radiation and protect the plants. This technology is based on plastic shade net products during whose production the chromophores and different light scattering and reflection elements are combined. These nets are designed to screen different spectral bands of solar radiation and/or convert direct light into diffuse light. The spectral manipulation aims to improve the selected physiological responses, the so-called photo-regulation process, whereas, in the light scattering process, the penetration of the modified light into the plant is enhanced. Photoselective shade net was first designed in Israel and was tested on ornamental plants, vegetables, and fruit trees. Then, it was spread to whole the world and was applied to plants, climatic regions, and agronomic practices (Shahak, Ratner et al., Citation2008).
The concept of colored nets is based on the distinguished filtration of sunlight along with the physical protection of the crops using specific nets in which some chromatic elements have been incorporated during manufacture. These nets are employed to absorb ultraviolet, blue, green, yellow, red, far-red, or near-infrared wavelengths. Along with their absorption, attention is paid to converting direct light into scattered light. Radiation spectrum is mostly manipulated to strengthen and stimulate plant physiological responses through photosynthetic and photomorphological processes. It should be mentioned that colored shade nets only change the components of solar radiation that pass through their plastic threads whereas the radiation that goes through the net holes are left untouched; that is, this is the same for diffuse components and direct sunlight that is prevalent in a certain time of day or season (Shahak, Ratner et al., Citation2008).
Researchers argue that the relative amount of blue/red light is an important factor in the technology of photoselective shade nets. Red/far-red ratio did not affect the crops shaded by colored nets. Yellow nets outperformed red nets in stimulating the vegetative growth, perhaps due to the stimulating effect of supplementing artificial green light (Kim, Goins, Wheeler, & Sager, Citation2004). Yellow nets differ from red nets in transferring green-yellow spectral range. However, it should be emphasized that these two nets differ in other effects too, such as the time of anthesis in cut flowers or fruit maturation in fruit trees (Oren-Shamir et al., Citation2003). The intense stimulating impact of yellow nets in both ornamental plants and fruit trees may be at least partially associated with the increased level of active gibberellins which is already proven to increase with the irradiance of supplemental green light (Wang & Folta, Citation2013). It has been documented that yellow and red, as well as pearl, nets increased the productivity of sweet peppers (Capsicum annuum) significantly when compared to conventional black nets. This has been attributed to the increase in the number of fruits per plant under these nets (Shahak, Gal et al., Citation2008). It has been reported that yellow nets could stimulate the vegetative growth of Pittosporum more than red nets. Also, it has been established that blue shade nets can reduce the vegetative growth and increase dwarfness in ornamental foliage and cut flower plants, whereas red and yellow nets that reduce the rate of blue light strengthen and stimulate the vegetative growth. Between yellow and red nets, the yellow has a much more strengthening effect than the red net (Shahak, Kon, & Ratner, Citation2016). Since there is little information available about the effect of photoselective shade-nets on growth parameters of the plants, the present study aimed to explore the effect of light variations by colored (red, green, and yellow) shade nets on the quantity and quality of experimental marigold and violet plants.
2. Materials and methods
2.1. Experimental design and location
To explore the effect of applying colored shade nets (red, green, and yellow) on physiological and morphological traits of marigold “Indian Prince” and violet “Yellow-Black” plants, an experiment was carried out in glasshouse of Ornamental Plant and Flower Research Station of Lahijan, Iran in 2017 and 2018. The temperature and humidity of the glasshouse were at 24–32◦C and 75–85%. The plants were planted in a garden soil substrate. Fertilization and irrigation were done identically for all plants. It was a split-plot experiment with two factors including colored shade net at four levels (no shade net, and green, yellow and red shade net) and plant species at two levels (marigold and violet) in a completely randomized block design with three replications. Each experimental plot was composed of three plants. The seeds of experimental plants and color nets were obtained from Bzr-Co and Fam-Co companies respectively. The shading intensity of color nets was about 50 %. The recorded vegetative parameters included plant height measured several times, plant growth rate (height difference), leaf and flower fresh and dry weight, flower number, leaf number, leaf area, and flower longevity.
2.2. Vegetative traits
To record the fresh and dry weight of the organs, they were weighed at the end of the experiment, were oven-dried at 105°C for 24 hours, and were re-weighed. Leaf length and width were measured for each sample. To determine leaf area, the leaves of each plant were detached, their length (L) and widest width (W) were measured, and the leaf area (A) was calculated by the following equation (Moll & Kamparth, Citation1977):
2.3. Physiological traits
The physiological traits included Brix°, chlorophyll a, b and total, carotenoid, anthocyanin, antioxidant capacity, and peroxidase. The Brix° of the leaves was measured with an N-1α hand-held refractometer (ATAGO Co., Japan).To measure chlorophyll contents of the treatments, 0.5 g of sample was weighed and ground in a mortar containing 50 cc 80% acetone (80 cc acetone + 20 cc distilled water). Then, the extract was infiltrated, adjusted to 50 cc, and poured into cuvettes. To determine chlorophyll content, it was read at 643 and 660 nm with a spectrophotometer. Chlorophyll a and b and total chlorophyll contents were estimated by the following equations (Mazumdar & Majumder, Citation2003):
To measure the carotenoid level, the treatments were sampled. Then, 0.5 g was weighed from the sample and was ground in a mortar containing 50 cc 80% acetone (80 cc acetone + 20 cc distilled water). Then, it was infiltrated, adjusted to 50 cc, and poured into cuvettes. The extracts were read at 645, 663, and 660 nm and were placed in the following equation, denoted by A, to determine carotenoid levels of the treatments (Mazumdar & Majumder, Citation2003):
To measure anthocyanin content, 0.5 of the sample was taken and ground in a Chinese mortar containing 50 cc of hydrochloric-ethanol acid (85% ethanol 95% + 15% hydrochloric acid). Then, it was infiltrated, adjusted to 50 cc, and poured into cuvettes. They were placed in a refrigerator at 4°C for 24 hours followed by 2 hours in darkness. The extract was read at 535 nm with a spectrophotometer and was placed in the following equation to determine anthocyanin content (Mazumdar & Majumder, Citation2003):
in which; a = sample weight (0.5 g),
b = the volume taken for the measurement (5 cc),
c = total volume (50 cc),
d = the fraction taken for the sample 0.1, and
e = absorption read at 535 nm.
To measure the enzymatic activity of peroxidase (POD), the extract was prepared as described above. Then, the variations of OD at 430 nm were read with a spectrophotometer once thirty seconds for two minutes (Chance & Maehly, Citation1995).In order to estimate antioxidant capacity, 1 g of the plant was wrapped in foil and was placed in liquid nitrogen for 2–3 minutes. Then, it was ground with 10 ml methanol 85% and the samples were placed in room temperature for one hour. Next, their extract was infiltrated and centrifuged for five minutes. Then, 150 ml was taken from it and was added with 850 μl DPPH. The solution was stirred fast and was kept in room temperature at the dark for 20 minutes. After placing the blank and resetting the instrument, first only DPPH was poured into the cuvette and it was read. Then, the sample was read at 517 nm by a spectrophotometer. The antioxidant capacity of the extracts was calculated by the following equation in terms of %inhibition in DPPH (Ramandeep & Savage, Citation2005):
where%DPPH = percent inhibition,
Acont = absorption rate of DPPH, and
Asamp = absorption rate (sample + DPPH).
2.4. Statistical analysis
Data were statistically analyzed with MSTATC Software Package, and the means were compared with the LSD test.
3. Results and discussion
The light intensity under the shade nets was assessed by a luxmeter on sunny, rainy, and cloudy days (Table ).The results of analysis of variance (ANOVA) in Table showed that plant height was significantly influenced by shade net type (p < 0.10) and by plant species (p < 0.01). But, the interaction of “shade net type × plant species” was not significant. According to means comparison, the highest plant height was obtained under the yellow net (Table ), related to marigold (Table ), and the lowest one was obtained from no-net treatment, which was related to violet.The results of ANOVA indicated that the effect of net type and plant species was significant on flowering stalk length (at the p < 0.05 and p < 0.01 levels, respectively), but the interaction of “shade net type × plant species” was not significant for this trait (Table ). The longest flowering stalk was observed under the yellow net (Table ) related to marigold (Table ). However, this did not differ from other nets significantly.We found that the yellow net improved morphological traits and vegetative potential of the plants so that enhancements occurred in height, growth rate, flowering stalk length, leaf number, and flower number of the plants shaded by the yellow net. It has been reported that photoselective shade nets cause different responses to similar PAR radiation intensities in leafy plants already exposed to black nets with 50–80% shading (e.g. Pittosporum variegatum, Fatsia japonica, and Montera deliciosa).Whilst blue nets inhibited the vegetative growth and caused the dwarfness of the plants, red and yellow nets enhanced vegetative growth including stem length and thickness, petioles, and leaf size in F. japonica and the overall yield of commercial cut flowers (Oren-Shamir et al., Citation2001; Shahak, Citation2008). A study on Pittosporum showed that yellow nets outperformed red nets in stimulating the vegetative growth of the plants (Shahak et al., Citation2016). In a study on the effect of different colored nets on Cordyline, Kumar Gaurav et al. (Citation2016) found that the plants grown under colored nets exhibited varied growth due to the spectral effect on plant growth.They reported that plants grown under white and red nets were taller than control (un-shaded) plants. Helianthus annuus, Lupinus luteus, Ornithogalum dubium and several cultivars of Eustoma grandiflorum were planted in a greenhouse under different colored nets with 50–60% shading. The results showed that red and yellow nets significantly increased stem length, stem weight, and inflorescence size versus black net, but the yellow net was better than the red one in stimulating the vegetative growth (Ovadia, Dori, Nissim-Levi, Shahak, & Oren-Shamir, Citation2009; Shahak, Citation2008), which is supported by our findings. Oren-Shamir et al. (Citation2003) treated several seasonal cut flowers (e.g. Lisianthus, Trachelium, Lupinus, and sunflower) with 50% shading nets. Both leafy plants and cut flowers produced longer and wider stems under red net than under black net; a similar response was observed under the yellow net, but the blue net was related to shorter stems.
Table 1. Light intensity under colored shade nets
Table 2. Analysis of variance of the effect of experimental factors on the recorded traits
Table 3. Means comparison for the effect of color shade nets on the studied traits
Table 4. Means comparison for the effect of plant species on the studied traits
Plant growth rate was significantly affected by shade net type and plant species (p < 0.01) and by the interaction of “shade net type × plant species” at the p < 0.05 level (Table ). Means comparison showed that the highest plant growth rate was related to the yellow net (Table ) obtained by violets (Table ), and the lowest was related to control plants.Also, the data pertaining to the interaction of “net type × plant species” indicated that “yellow net × violet” had the highest plant growth rate and “green net × marigold” had the lowest (Table ).
Table 5. Means comparison for the interaction of “shade net type × plant species” for the studied traits
Researchers have argued that better performance of yellow shade nets than red nets in stimulating vegetative growth is likely to relate to the stimulating effect of artificial green light supplementation (Kim et al., Citation2004; Oren-Shamir et al., Citation2003). A study on peach (Prunus persica “Hermosa”) showed the increased growth under yellow and red nets (Shahak et al., Citation2004a, Citation2004b).In our study too, the highest growth rate was observed in plants treated with yellow nets.A study on the effect of colored shade nets (20–30% shading in PAR) on several cultivars of table grape in four different regions indicated that yellow nets increased plant size significantly in all cases (Shahak, Ratner et al., Citation2008).
According to the results of ANOVA, the effect of shade net was significant on leaf number (p < 0.01) and the effect of plant species was significant at the p < 0.05 level, but their interaction was not significant for this trait (Table ).The results of means comparison showed that different photoselective shade nets increased the number of leaves, but they did not differ significantly. Control plants exhibited the least leaves (Table ).The number of flowers was significantly influenced byshade nets (p < 0.01) and plant species (p < 0.05).But, the interaction of “shade net × plant species” was not significant for this trait (Table ). Means comparison showed that the highest number of flowers was produced under yellow nets and the lowest was related to control (Table ). In addition, means comparison for the effect of plant species on flower number showed that the highest number of flowers was produced by violet plants (Table ). Since the number of flowers is a critical trait in ornamental plants and we found that the flower number was increased under the yellow net, so it is of crucial importance to use this shade net in flower industry.
In a study on the effect of light modification by colored nets on lettuce, Ilić et al. (Citation2017) reported that the highest number of leaves was obtained from pearl and red nets and the lowest from control. Although both red and yellow nets improved the vegetative growth of cut flowers, they may differ in terms of the effect of the time required for flowering. Indeed, for Ornitogalum dubium, red net advanced flowering when compared to control plants that were exposed to black shade net, whereas yellow shade net postponed flowering (Ovadia et al., Citation2009; Shahak, Citation2008). It has been reported that pearl nets increased branch number in pot Myrtuscommunis whereas in “Poorindaextasy” Crowea, the number of flowers per branch increased versus the black net (Nissim-Levi et al., Citation2008). It has been shown that photoselective shade nets can change shading quality by scattering light and changing spectral composition (Shahak, Gal, et al., Citation2008).
The results of ANOVA indicated that the effect of shade net was significant at the p < 0.05 level and the effect of plant species was significant at the p < 0.01 level on leaf fresh and dry weight (Table ). Means comparison for the effect of shade net on leaf fresh weight (Table ) showed that the highest leaf fresh weight was obtained from plants treated with yellow net and so was the highest leaf dry weight, but it did not differ from red and green nets significantly. Flower fresh weight was significantly influenced by shade net and plant species at the p < 0.01 level and by their interaction at the p < 0.05 level. Also, flower dry weight was significantly affected by shade net type at the p < 0.05 level and by plant species at the p < 0.01 level (Table ). The highest flower fresh weight was related to the yellow net (Table ) obtained by marigolds (Table ), and the highest flower dry weight was also related to green net obtained by marigolds again. The lowest flower fresh and dry weights were observed in control treatment. Mean data for the interactive effect of “net type × plant species” on flower fresh weight showed that “yellow net × marigold” produced the highest flower fresh weight and “no-net × violet” produced the lowest one (Table ).
Kumar Gaurav et al. (Citation2016) reported that the highest leaf fresh weight was obtained from the red net in which it was 85.88% higher than that of control. The highest leaf dry weight was also produced under the red net. Overall, colored shade-nets had the highest impact on this trait when compared to control that was not shaded. We also found that the highest leaf and flower fresh and dry weights were obtained from the colored shade nets.Plant response to shade nets varies with light quality and quantity (Lee et al., Citation1997). Typically, growth and biomass production is determined by light intensity whereas growth, development, and morphogenesis are dictated by light composition (quality) (Stuefer & Huber, Citation1998). In our experiment, the yellow net was related to higher fresh weight than other nets and control. As well, the highest plant growth rate was obtained fromthe red net, implying the effect of this shade net on the improvement of plant quantitative and qualitative traits. Therefore, it can be said that colored shade nets, especially yellow nets, enhance the biomass of marigolds and violets.It was found that the effect of net type was significant on flower longevity (p < 0.01). But, the trait was not significantly influenced by plant species and “net type × plant species” (Table ). According to means comparison for the effect of shade net on flower longevity (Table ), the longest longevity was 43.67 days related to yellow nets and the shortest was 31.17 days related to control.
Kumar Gaurav et al. (Citation2016) reported the longest vase life in different colored nets and stated that this increase may have been related to the role of nets in protecting the leaves against intense light. Parameters like plant species and cultivar, growth stage, light intensity and quality, and other environmental conditions affect the physiological, morphological, and anatomical responses of the plants to the use of different compositions of red and blue spectra (Fan, Xu, Liu, Tang, & Wang, Citation2013; Islam et al., Citation2012; Takemiya, Inoue, Doi, & Kinoshita, Citation2005). It has been documented that colored nets affect blooming and quality of cut flowers whose response vary with species and cultivar (Oren-Shamir et al., Citation2003). Light quality in terms of color and wavelength can affect the morphological structure of the plants (Li, Tang, & Xu, Citation2013). Therefore, the change in light quality and different spectra passing through the nets in controlled environments may improve the quantity and quality of the plants.
The effect of shade net type was significant on leaf length at the p < 0.01 level, and the effect of plant species was significant on leaf size (leaf length and width) at the p < 0.01 level, but the interaction of “shade net type × plant species” was insignificant for leaf size (Table ). According to means comparison, the longest leaves were produced under the red net and the shortest ones in control (Table ). Also, means comparison for the effect of plant species on leaf size indicated that the highest leaf size was related to marigold (Table ). As the results of ANOVA indicated, leaf area was significantly influenced by shade net type at the p < 0.05 level and by plant species at the p < 0.01 level, but the interaction of these two parameters was insignificant for this trait (Table ). The highest leaf area was 20.33 cm2 observed in red-shaded plants and the lowest was 14.14 cm2 observed in control plants (Table ). The highest leaf area was related to marigold as means comparison for the effect of plant species on leaf area showed (Table ).
We figured out that leaf area in red-shaded plants was 1.5 times as great as that of control plants.The increase in leaf area can be attributed to the relationship between plants and atmosphere. The expansion of leaf area includes the generation of new leaves, the increase in the size of the existing leaves, and the senescence of the old leaves (Ranganathan et al., Citation2001). Therefore, the measurement of leaf area is important to assess plant response to environmental conditions.Cristofori, Rouphael, Mendoza-de Gyves, and Bignami (Citation2007) reported that the best model to estimate leaf area is a linear model that is resulted from multiplying leaf length and width.We, also, used leaf length and width to measure leaf area. Leaf area is a very beneficial parameter for plant growth. It is defined as plant capacity to produce dry matter as per the use of radiation and the photosynthesis rate. Kumar Gaurav et al. (Citation2016) also showed that the highest leaf area of cordyline was obtained from red shade nets, which is consistent with our findings.Colored shade nets influenced both growth period and morphological traits of lettuceso that leaf area index was enhanced and its growth period was shortened remarkably (Ilić et al., Citation2017).
According to the results of ANOVA, the effect of shade net was significant on Brix° (°Bx) at the p < 0.05 level and the effect of plant species was significant at the p < 0.01 level, but the interaction of “shade net type × plant species” was insignificant (Table ). The highest °Bx value was 7.13% obtained from the yellow net and the lowest was 4.17% obtained from control, but this did not differ from that of the green net (5.30%) significantly (Table ). Means comparison for the effect of plant species on the °Bx value showed that the highest value of 7.69% was obtained from marigold plants (Table ).
The °Bx value is a measure of sugar content in the solution, which depends on light refraction and shows the weight percentage of solids existing in a solution with respect to the total weight of the solution. Our results showed that the flowers treated with yellow nets had higher °Bx value than the other nets. This increase is likely to associate with the stimulating effect of artificial green light supplementation under yellow nets (Kim et al., Citation2004).
The effect of shade net type, plant species and “shade net type × plant species” was significant on anthocyanin content at the p < 0.01 level (Table ). The highest anthocyanin content was obtained from the green net (Table ) related to violet (Table ), and the lowest was related to control plants (Table ). With respect to the interaction of the two parameters, the highest anthocyanin content was observed in violet plants exposed to the green net (Table ). Mancinelli (Citation1990) found that three properties of light–color, intensity, and duration—influenced plant growth and that the red:far-red ratio was influenced by the duration and photon flux in light treatment, influencing growth and anthocyanin synthesis significantly. Lefsrud, Kopsell, and Sams (Citation2008) reported that anthocyanin content was increased in LED light-treated cabbages. In our study, the green shade net increased anthocyanin content. This is related to the responses of different plants under different environmental conditions to the photoselective shade nets. The results of ANOVA showed that the effect of shade nets and plant species was significant on chlorophyll a content at the p < 0.01 level, but the interaction of “shade net type × plant species” was insignificant (Table ). Means comparison for the effect of shade net type on chlorophyll a content (Figure ) showed that the highest chlorophyll a content was related to the yellow net and violet plants and the lowest was related to the green net. ANOVA revealed that the impact of shade net type and plant species was significant on chlorophyll b content at the p < 0.01 level and the interaction of “shade net type × plant species” was significant at the p < 0.05 level (Table ). Means comparison indicated that the highest chlorophyll b content was obtained from yellow shade net and among the species, violet had the highest chlorophyll b content (Figure ). Data for the interactive effect of “shade net × species” on chlorophyll b content indicated that “no net × violet” and “yellow net × violet” had the highest one, but the other treatment did not differ significantly (Table ). According to the results of ANOVA (Table ), the effect of shade net kind and plant species was significant on total chlorophyll content at the p < 0.01 level and the interaction of “shade net type × plant species” was significant for this trait at the p < 0.05 level. Means comparison for the effect of shade net type on total chlorophyll (Figure ) indicated that the highest total chlorophyll was related to control and red shade net, and the red and green shade nets had no significant difference to one another.In addition, it was found that the highest total chlorophyll content was related to violet. The highest total chlorophyll content was obtained from “no shade net × violet” and “yellow shade net × violet” (Table ).
Figure 1. Effect of color net type on plantchlorophyll.
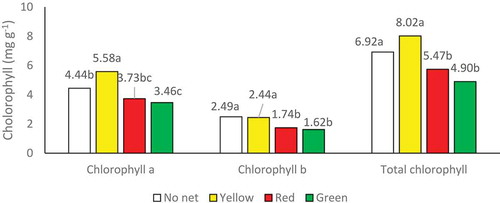
Light is not only the mere source of photosynthesis energy (impact on chlorophyll), but it is also an intermediate of photo-morphogenesis (Liu, Citation2012).When plants are exposed to very intense light, the chlorophyll degradation rate exceeds its synthesis rate, resulting in the reduction of chlorophyll content due to the inhibition of chloroplast synthesis (Fu, Li, & Wu, Citation2012; JFDC, Marenco, & Vieira, Citation2001). Although the maximum light absorption by chlorophyll happens in the red spectrum, this can be explained by the fact that the natural growth and propagation of the plants can be kept on only by the retention of certain amounts of supplemental radiation so that plants grown under red shade nets need to be supplemented with blue radiation (Brown, Schuerger, & Sager, Citation1995). Lettuce plants grown under photoselective shade nets had higher total chlorophyll, chlorophyll a, and chlorophyll b contents than control (Ilić et al., Citation2017). The variation of chlorophyll content in our study was related to the photo-conditions under the red nets at the trial time, which created a specific situation of the plants.
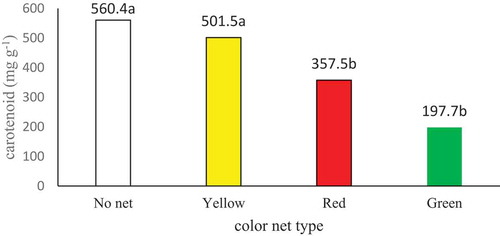
The simple effect of shade net type and plant species was significant on plant carotenoid content at the p < 0.01 level and the interaction of “shade net type × plant species” was significant for this trait at the p < 0.05 level (Table ). According to means comparison, the highest carotenoid was obtained from control and the yellow net, and the lowest from the green net (Figure ). With respect to the effect of plant species, it was found that the violets had the highest carotenoid content of 490.83 mg L−1 (Table ). Means comparison for the interactive effect of shade net type and plant species on carotenoid content indicated that the treatments “no net × violet” and “yellow net × violet” had the highest carotenoid content of 728.1 and 624 mg L−1, respectively, whereas “green net × marigold” had the lowest one of 122.2 mg L−1 (Table ). Carotenoids can absorb energy, which may lead to the formation of free oxygen out of the excited chlorophyll molecules. Carotenoids may, also, degrade free oxygen molecules synthesized during photosynthesis (Bergquist, Citation2006). It has been established that the carotenoid content of tomato was higher under red and pearl shade nets (Selahle, Sivakumar, & Soundy, Citation2014; Tinyane, Sivakumar, & Soundy, Citation2013). Kong, Avraham, Ratner, and Shahak (Citation2012) reported that red shade net induced morphological changes and increased leaf carotenoid content in peppers as compared to red shade nets,which may be attributed to the increased amount of green light under yellow nets.They related the increase in carotenoid content under yellow nets, when compared tored nets, to the adaptation with the enrichment of the green spectrum. We, also, saw the increase in carotenoid content under yellow nets. In a study on the effect of colored nets on lettuce, Ilić et al. (Citation2017) stated that carotenoid content in the leaves of lettuces grown under photoselective nets was higher than that of control plants (not treated with nets).
Figure 2. Effect of color net type on plant carotenoid.
According to the results of ANOVA, peroxidase was significantly influenced by the shade net type at the p < 0.05 level and the by the interaction of “shade net type × plant species” at the p < 0.10 level, but the effect of plant species was insignificant on this trait (Table ). The highest peroxidasewas observed in plants grown under red and green shade nets (Table ). The highest peroxidase was obtained from “green net × violet” and the lowest from “yellow net × violet” (Table ). Peroxidases are involved in most cellular processes, such as auxin metabolism, wood formation, transverse links in plant cell walls, response to environmental stresses, and so on (Yamasaki, Sakihama, & Ikehara, Citation1997).
Antioxidant capacity was significantly affected by shade net and “shade net type × plant species” at the p < 0.10 level and by plant species at the p < 0.05 level (Table ). Violet plants grown under green shade net had the highest antioxidant capacity, whereas those grown in the shade of red net exhibited the lowest antioxidant capacity (Table ). Means comparison for the interaction “net type × plant species” showed that the highest antioxidant capacity related to “green net × violet” (Table ). Antioxidants occur in plants naturally.They include such compounds as phenols and anthocyanins (Wang & Zheng, Citation2001). Antioxidant activity was more intense in plants newly grown under yellow and red shade nets. In addition, there was no significant difference in the post-harvest antioxidant content of the leaves of plants grown under different nets (Buthelezi, Soundy, Jifon, & Sivakumar, Citation2016). The control of the radiation quality by the use of red and pearl photo-selective nets contributes to maintaining antioxidant activity of the vegetables after their flowering. The increase in antioxidant levels after the manipulation of environmental light by the use of pearl nets can improve the quality of fruits in storages (Kong et al., Citation2013). Plants grown under pearl nets had significantly higher total phenols, flavonoids, and antioxidant properties than plants treated with other nets (Ilić et al., Citation2017). Wu et al. (Citation2007) showed that red LED light significantly increased antioxidants of peas by a factor of two. Kong et al. (Citation2013) found that the antioxidants of peppers were increased under yellow shade nets, implying that yellow and pearl nets may be involved in improving plant resistance to biotic stresses. These studies show that plant response to the increase in antioxidant capacity varies with the type of the photoselective net. It seems that the response of plants is different to various light spectra.
4. Conclusions
Among the studied photoselective shade nets, yellow shade net hasa much stronger effect than other shade nets and improves the vegetative growth and physiological traits of the studied plant species.
Overall, it was found that the application of photoselective shade nets provide the plants with more optimal growth conditions than when they were not used.
It seems that the passing of photosynthetically active radiation from the red shade net (i.e. red and orange spectra of visible light) enhanced important plant phenomena, such as photosynthesis and consequently, the biomass.
The results of this experiment can use by gardeners and greenhouse owners.
Additional information
Funding
Notes on contributors
Shahram Sedaghathoor
Shahram Sedaghathoor is currently Associate Professor of Horticultural Science at the Faculty of Agricultural Sciences, Rasht branch, Islamic Azad University, Rasht, Iran. This study is a part of PhD dissertation of Miss Seyedeh Khadijeh Abbasnia Zare. Shahram Sedaghathoor and Mohammad-Naghi Padasht Dahkaei are her supervisors and Davood Hashemabadi is her advisor.
References
- Bergquist, S. (2006). Bioactive compounds in baby spinach (Spinacia oleracea L.). Effects of pre- and postharvest factors ( Doctoral dissertation). Swedish University of Agricultural Sciences, Alnarp, Sweden.
- Brown, C. S., Schuerger, A. C., & Sager, J. C. (1995). Growth and photomorphogenesis of pepper plants under red lightemitting diodes with supplemental blue or far-red lighting. Journal of the American Society for Horticultural Science, 120, 808–16.
- Buthelezi, M. N. D., Soundy, P., Jifon, J., & Sivakumar, D. (2016). Spectral quality of photo-selective nets improves phytochemicals and aroma volatiles in coriander leaves (Coriandrum sativum L.) after postharvest storage. Journal of Photochemistry and Photobiology B: Biology., 161, 328–334. doi:10.1016/j.jphotobiol.2016.05.032
- Chance, B., & Maehly, A. C. (1995). Assay of catalase and peroxidase. In S. P. Colowick & N. D. Kaplan (Eds.), Methods in enzymology (Vol. 2, pp. 764–791). New York: Academic Press. doi:10.1016/s0076-6879(55)02300-8
- Cristofori, V., Rouphael, Y., Mendoza-de Gyves, E., & Bignami, C. (2007). A simple model for estimating leaf area of hazelnut from linear measurements. Scientia Horticulturae, 113, 221–225. doi:10.1016/j.scienta.2007.02.006
- Fan, X., Xu, Z., Liu, X., Tang, C., & Wang, L. (2013). Effect of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Scientia Horticulturae, 153, 50–55. doi:10.1016/j.scienta.2013.01.017
- Forghani, H., & Kiani Abri, M. (2005). A study on relative advantage of flower and ornamental plant production and exporting in selected provinces. Journal of Human and Social Science, 5, 53–64.
- Fu, W., Li, P., & Wu, Y. (2012). Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Scientia Horticulturae, 135, 45–51. doi:10.1016/j.scienta.2011.12.004
- Ilić, S. Z., Milenković, L., Dimitrijevićb, A., Stanojevićc, L., Cvetkovićc, D., Ž, K., … Mastilovićd, J. (2017). Light modification by color nets improve quality of lettuce from summer production. Scientia Horticulturae, 226, 389–397. doi:10.1016/j.scienta.2017.09.009
- Islam, M. A., Kuwar, G., Clarke, J., Blystad, D. R., Gislerod, H. R., Olsen, J. E., & Torre, S. (2012). Artificial light from light emitting diodes (LEDs) with a high portion of blue light results in shorter poinsettias compared to high pressure sodium (HPS) lamps. Scientia Horticulturae, 147, 136–143. doi:10.1016/j.scienta.2012.08.034
- JFDC, G., Marenco, R. A., & Vieira, G. (2001). Concentration of photosynthetic pigments and chlorophyll fluorescence of mahogany and tonka bean under two light environments. Revista Brasileira De Fisiologia Vegetal, 13, 149–157. doi:10.1590/s0103-31312001000200004
- Kim, H. H., Goins, G. D., Wheeler, R. M., & Sager, J. C. (2004). Green-light supplementation for enhanced lettuce growth under red- and blue-light-emitting diodes. HortScience, 39(7), 1617–1622. doi:10.21273/HORTSCI.39.7.1617
- Kong, Y., Avraham, L., Perzelan, Y., Alkalai-Tuvia, S., Ratner, K., Shahak, Y., & Fallik, E. (2013). Pearl netting affects postharvest fruit quality in ‘vergasa’ sweet pepper via light environment manipulation. Scientia Horticulturae, 150, 290–298. doi:10.1016/j.scienta.2012.11.029
- Kong, Y., Avraham, L., Ratner, K., & Shahak, Y. (2012). Response of photosynthetic parameters of sweet pepper leaves to light quality manipulation by photoselective shade nets. Acta Horticulturae, 956, 501–506. doi:10.17660/actahortic.2012.956.59
- Kumar Gaurav, A., Raju, D. V. S., Janakiram, T., Singh, B., Jain, R., & Gopala Krishnan, S. (2016). Effect of different coloured shade nets on production and quality of Cordyline. Indian Journal of Agricultural Sciences, 86(7), 865–869.
- Lee, D. W., Oberbauer, S. F., Krishnapilay, B., Mansor, M., Mohamad, H., & Yap, S. K. (1997). Effects of irradiance and spectral quality on seedling development of two Southeast Asian Hopea species. Oecologia, 110, 1–9. doi:10.1007/s004420050126
- Lefsrud, M. G., Kopsell, D. A., & Sams, C. (2008). Irradiance from distinct wave-length lightemitting diodes affect secondary metabolites in kale. HortScience, 43, 2243–2244. doi:10.21273/HORTSCI.43.7.2243
- Li, H., Tang, C., & Xu, Z. (2013). The effects of different light qualities on rapeseed (Brassica napus L.) plantlet growth and morphogenesis invitro. Scientia Horticulturae, 150, 117–124. doi:10.1016/j.scienta.2012.10.009
- Liu, W. (2012). Light environmental management for artificial protected horticulture. Agrotechnology, 1, 1–4. doi:10.4172/2168-9881.1000101
- Mancinelli, L. (1990). Interaction between light quality and light quantity in the photoregulation of antocyaninproduction. Plant Physiology, 92, 1191–1195. doi:10.1104/pp.92.4.1191
- Mazumdar, B. C., & Majumder, K. (2003). Methods on physcochemical analysis of fruits (pp. 136–150). Calcutta: University College of Agriculture.
- Moll, R. H., & Kamparth, E. J. (1977). Effect of population density upon agronomic traits associated with genetic increases in yield of (Zea mays L.). Agronomy Journal, 69, 81–84. doi:10.2134/agronj1977.00021962006900010021x
- Nissim-Levi, A., Farkash, L., Hamburger, D., Ovadia, R., Forrer, I., Kagan, S., & Oren- Shamir, M. (2008). Light-scattering shade net increases branching and flowering in ornamental pot plants. The Journal of Horticultural Science and Biotechnology, 83, 9–14. doi:10.1080/14620316.2008.11512340
- Oren-Shamir, M., Gussakovsky, E. E., Shpiegel, E., Nissim-Levi, A., Ratner, K., Ovadia, R., … Shahak, Y. (2001). Coloured shade nets can improve the yield and quality of green decorative branches of Pittosporumvariegatum. The Journal of Horticultural Science and Biotechnology, 76, 353–361. doi:10.1080/14620316.2001.11511377
- Oren-Shamir, M., Shahak, Y., Dori, I., Matan, E., Shlomo, E., Ovadia, R., … Ganelevine, R. (2003). Lisianthus: enhanced elongation of summer cut flowers grown under colored shade nets. Flortecnica, 6, 84–86.
- Ovadia, R., Dori, I., Nissim-Levi, A., Shahak, Y., & Oren-Shamir, M. (2009). Coloured shade nets influence thestem length, time to flower and flower size of ornamental crops. The Journal of Horticultural Science and Biotechnology, 84, 161–166. doi:10.1080/14620316.2009.11512498
- Ramandeep, K. T., & Savage, P. G. (2005). Antioxidant activity in different factions of tomatoes. Food Research International, 38, 487–494. doi:10.1016/j.foodres.2004.10.016
- Ranganathan, R., Chamhan, Y. S., Flower, D. J., Robertson, M. J., Anetra, C. S., & Silim, S. N. (2001). Predecting growth and development of pigeonpea: leaf area development. Field Crops Research, 69, 163–172. doi:10.1016/s0378-4290(00)00137-4
- Safaii, B., Khalaji Asadi, M., Taghzadeh, H., Jilavi, A., Taleghani, G., & Danesh, M. (2005). Estimating solar energy potential in Iran and related radiation atlas. Journal of Nuclear Science and Technology, 33, 27–34.
- Selahle, M. K., Sivakumar, D., & Soundy, P. (2014). Effect of photo-selective nettings on postharvest quality and bioactive compounds in selected tomato cultivars. Journal of the Science of Food and Agriculture, 94, 2187–2195. doi:10.1002/jsfa.6536
- Shahak, Y. (2008). Photoselective netting for improved performance of horticultural crops. A Review Ofornamental and Vegetable Studies Carried in Israel. Acta Horticulturae.770:, 161–168. doi:10.17660/actahortic.2008.770.18
- Shahak, Y., Gal, E., Offir, Y., & Ben-Yakir, D. (2008). Photoselective shade netting integrated with greenhouse technologies for improved performance of vegetable and ornamental crops. Acta Horticulturae, 797, 75–80. doi:10.17660/actahortic.2008.797.8
- Shahak, Y., Gussakovsky, E. E., Cohen, Y., Lurie, S., Stern, R., Kfir, S., … Greenblat-Avron, Y. (2004a). Color Nets: a new approach for light manipulation in fruit trees. Acta Horticulturae, 636, 609–616. doi:10.17660/actahortic.2004.636.76
- Shahak, Y., Gussakovsky, E. E., Gal, E., & Ganelevin, R. (2004b). ColorNets: crop protectionand light-qualitymanipulation in one technology. Acta Horticulturae, 659, 143–151. doi:10.17660/actahortic.2004.659.17
- Shahak, Y., Kon, Y., & Ratner, K. (2016). The wonders of yellow netting. Acta Horticulturae, 1134, 327–334. doi:10.17660/actahortic.2016.1134.43
- Shahak, Y., Ratner, K., Giller, Y. E., Zur, N., Or, E., Gussakovsky, E., … Greenblat-Avron, Y. (2008). Improving solar energy utilization, productivity and fruit quality in orchards and vineyardsbyphotoselective netting. Acta Horticulturae, 772, 65–72. doi:10.17660/actahortic.2008.772.7
- Stuefer, J. F., & Huber, H. (1998). Differential effects of light quantity and spectral light quality on growth, morphology and development of two stoloniferous Potentilla species. Oecologia, 117, 1–8. doi:10.1007/s004420050624
- Takemiya, A., Inoue, S., Doi, M., & Kinoshita, T. (2005). Phototropins promote plant growth in response to blue light in low light environments. The Plant Cell, 17, 1120–1127. doi:10.1105/tpc.104.030049
- Tinyane, P. P., Sivakumar, D., & Soundy, P. (2013). Influence of photo-selective netting on fruit quality parameters andbioactive compounds in selected tomato cultivars. Scientia Horticulturae, 161, 340–349. doi:10.1016/j.scienta.2013.06.024
- Wang, S. Y., & Zheng, W. (2001). Effect of plant growth temperature on antioxidant capacity in strawberry. Journal of Agricultural and Food Chemistry, 49, 4977–4982. doi:10.1016/j.scienta.2013.06.024
- Wang, Y., & Folta, K. M. (2013). Contributions of green light to plant growth and development. American Journal of Botany, 100, 70–78. doi:10.3732/ajb.1200354
- Wu, M. C., Hou, C. Y., Jiang, C. M., Wang, Y. T., Wang, C. Y., Chen, H. H., & Chang, H. M. (2007). A novel approach of LED light radiation improves the antioxidant activity of peaseedlings. Food Chemistry, 101, 1753–1758. doi:10.1016/j.foodchem.2006.02.010
- Yamasaki, H., Sakihama, Y., & Ikehara, N. (1997). Flavonoid-peroxidase reaction as a detoxification mechanism of plant cells against H2O2. Plant Physiology115, 1405–1412. doi:10.1104/pp.115.4.1405