
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
In nature, allelopathic substances are likely to influence the germination of seeds. Callistemon viminalis, Tephrosia vogelii, Senna spectabilis, Cupressus lusitanica, and Polyscias fulva are found around some tomato culture fields in Cameroon. These plants may produce allelochemicals that can influence tomato seed germination and seedling characteristics. Methanol extracts and their fractions were prepared from leaves of these plants and tested on seed germination rate, shoot diameter, root and stem lengths, and vigor index of seedlings. The tested substances did not significantly (p > 0.05) affect the seed germination rate. Aqueous extracts of T. vogelii, C. lusitanica, and C. viminalis exerted the highest inhibition on seed germination while methanol extract, hexane, and ethyl acetate fractions of all the plants induced significant increases in diameters of seedlings. Aqueous extracts were effective on the stem length. All treatments generally reduced the root length. Methanol extracts induced significantly higher vigor indices (p < 0.05).
Public Interest Statement
With increasing global population and decreasing arable soil superficies, achieving food security has become a challenge for most of governments. The major threats to crop production are weeds, insect pests, diseases, and drought. Unfavorable but unavoidable climatic changes are other important threats to food security. In order to face some of these challenges, there is use of great quantities of chemical inputs whose excessive use has negative impacts of the quality of agricultural commodities, environment, and human health. That is why aware and well-adviced consumers increasingly prefer produces which are not susceptible to negatively affect their health. Allelopathy has emerged as one of the sustainable and integrated approaches (e.g. crop rotations, cover crops, intercropping, use of powder, or extracts of biological material) to tackle these problems. The present findings, that some plant extracts led to the improvement in tomato seedling growth parameters, are in straight line with this approach.
Competing Interest
The authors declare no competing interest.
1. Introduction
Tomato (Solanum lycopersicum L.) is one of the most produced and consumed fruits in world, and Cameroon in particular. It is a rich source of vitamins A and C, folic acid, α-lipoic acid, lycopene, choline, β-carotene, and lutein (Afzal et al., Citation2009). Its production in Cameroon has significantly increased, passing from 25,000 t in 1960 to 954,000 t in 2013 (FAOSTAT, Citation2013). However, this production is unequally distributed in the country, with the western highlands among the high production areas. According to Tonfack et al. (Citation2009), tomato cultures in the western highlands of Cameroon produce very weak outputs despite the use of large quantities of fertilizers. It is known that the production yield of tomato and other plants depends on many factors including soil nature, pest management, the variety used, seed germination, and seedling problems. It can also be affected by neighboring plants through allelopathic effects.
Indeed, plants interfere with each other in their environments in different ways, through secondary metabolites released into the soil as exudates or resulting from the decomposition of plant residues (Kuzyakov, Hill, & Jones, Citation2007). Besides the classic competition for water, nutrients and light, it has been demonstrated that many plant species synthesize allelopathic molecules capable of inhibiting or stimulating the germination and growth of neighboring plants (Dayan, Romagni, & Duke, Citation2000). Allelopathy is then considered as an ecological and chemical interaction characterized by stimulatory or inhibitory effects among different plant families (Badmus & Afolayan, Citation2012). This is particularly evident in the context of integrated and organic agricultural production, as an alternative to chemical treatments against weeds that are currently scarce and very expensive, with the allelopathy phenomenon being highly selective.
In the west Region of Cameroon, tomato is very often grown together with other plant species such as Senna spectabilis, Tephrosia vogelii, Polyscias fulva, Cupressus lusitanica and Callistemon viminalis, some of which are ornamental and others used as fertilizers. Sometimes, the leaves of S. spectabilis or T. vogelii are wrinkled and spread on the soil before the sowing. The potential allelopathic effect of C. lusitanica (used to materialize the limits of a field) on seed germination, radicle, and seedling growth was investigated on chickpea, maize, pea, and teff (Lisanework & Michelsen, Citation1993). Cupressus arizonica extracts also showed an inhibitory effect on the seed germination and seedling growth of Lolium perenne and Poa pratensis. The author found that the inhibition rates of the two studied grasses increased with extract concentration (Aliloo, Citation2012). Bioassay showed that the aqueous extract from fresh leaves of T. vogelii significantly inhibited seed germination and seedling growth of Festuca arundinacea, Cynodon dactylon, and Digitaria sanguinalis (Ruilong et al., Citation2011).
Based on the above facts, we hypothesized that C. lusitanica, S. spectabilis, T. vogelii, C. viminalis, and P. fulva may contain substances that could affect the germination of tomato and subsequently plant vigor index in the field. This can be verified in vitro in a preliminary study before field investigation. Thus, this work was designed to investigate the in vitro allelopathic effects of extracts and their fractions from these five plant species on tomato seed germination and vigor.
2. Material and methods
2.1. Plant material
Plant material consisted of leaves from the following species: C. viminalis, T. vogelii, S. spectabilis, C. lusitanica, collected in January 2014 at the campus of the University of Dschang, as well as P. fulva collected in May 2014 in Bafou, all situated in the western administrative Region of Cameroon. The plant materials were identified at the Cameroon National Herbarium in Yaoundé where a voucher specimen was kept, respectively, under the reference numbers 49872/HNC, 43546/HNC, 45740/HNC, 35436/HNC, and 47801/HNC. The leaves of each plant species were dried in the shade for three weeks at 22 ± 2°C indoor. They were finely crushed in a mechanical mill and the resulting powders were used to prepare plant extracts.
Tomato seeds (Rio Grande, lot N° 58480, packaging date September 2013. Vikima Seed A/S from Denmark) respecting the EC Standard Norms, were purchased from Holland Farming Cmr Sarl, Cameroon. All experiments were conducted in a randomized complete block design. Treatments of each experiment were replicated three times and all experiments were repeated thrice. The initial seed germination count was carried out after two days of incubation. The final germination, percentage of inhibition, seedling diameter, and lengths were evaluated after 10 days.
2.2. Preparation of plant extracts and fractions
Aqueous extracts were prepared by macerating, respectively, 2.5, 1.25, and 0.625 g of plant powder in 100 ml of distilled water for 24 h. These extracts were then filtered using Whatman paper No. 1.
Methanolic extracts were prepared by macerating each plant powder (1,000 g) in 6 L methanol for two days, and then filtered using Whatman paper No. 1. The evaporation of solvent was carried out using a rotary evaporator (Buchi R-200) under vacuum at 40°C. The extracts obtained were placed in an oven at 40°C for 24 h to remove residual solvent.
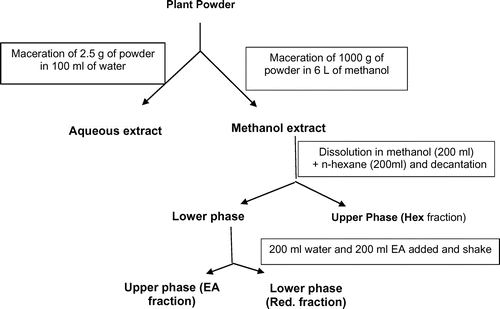
The methanol extracts were successively and separately partitioned using n-hexane and ethyl acetate. For this, 80 g of each methanol extract were dissolved in 200 ml of methanol. To this solution, 200 ml of hexane were added and the mixture was gently shaken and the two phases were separated using separating funnel. The upper phase, the hexane fraction, was kept aside while to the lower portion, 200 ml of water and 200 ml of ethyl acetate were added. After shaking, the upper phase constituted the ethyl acetate fraction while the lower phase was the residual fraction. The solvents were evaporated under vacuum at 40°C to give the hexane fraction (Hex), the ethyl acetate fraction (EA), and the residual fraction (Res) (Figure ).
Figure 1. General procedure of plant extracts and fractions preparation.
2.3. Treatment and seed germination
Stock solutions of methanolic extracts and their fractions were prepared by dissolving separately 10 mg of each extract in 800 μl of Tween 80 (surfactant) and the total volume adjusted to 8 ml with sterile distilled water for a final concentration of 1.25 mg/ml. Tested concentrations (0.625, 0.312, and 0.156 mg/ml) were obtained by serial dilution of the stock solutions. Positive control consisted of a 0.025 μg/ml naphthalene acetic acid (NAA) from a stock solution of 10 μg/ml in sterile distilled water, while distilled water was used as negative control.
The tomato seed germination experiment was carried out using 90 mm Petri dishes containing two layers of Whatman papers No. 2. Ten milliliters of tested solutions were used to moisten this paper. Seed germination was monitored by looking for a visible protrusion of the radicle from day 1 of experiment onwards. All sets of treatments and controls were prepared in triplicate of 10 seeds each and the experiment was repeated three times. The experiment was carried out at 25 ± 2°C and 12 h photoperiod. The germination percentage was calculated using the following formula:
2.4. Evaluation of physiological growth parameters
The measurements of the diameter, length of stems and roots were made on the fifteenth day of culture. The diameter of the plants was measured using a micrometer brand. The stem and root length was measured using a straight edge in cm.
The vigor index was calculated by the formula of Patel and Manhad (Citation2014).
The data from the repeated experiments were pooled before being subjected to ANOVA and treatment means separated by Duncan’s Multiple Range Test at 5% probability level.
3. Results and discussion
3.1. Results
3.1.1. Effect of extract/fraction on the germination kinetics of tomato seeds
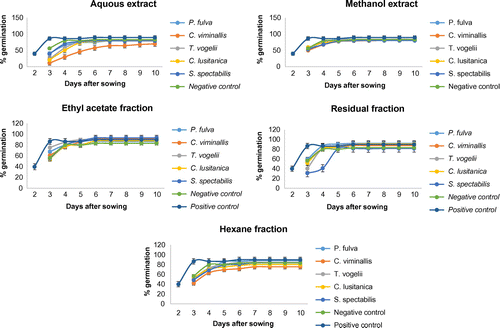
The kinetics of tomato seed germination in the presence or absence of plant extracts are presented in Figure . The extracts or fractions used did not have any effect on the time taken for the seeds to germinate. However, the positive control accelerated seed germination, with a germination time of 2 days, instead of 3 days observed in the negative control or extract/fraction-treated groups. Moreover, the percentage of germination generally increased with time up to day 6. The plant extracts and their fractions significantly (p < 0.05) and differentially affected the percentage of seed germination. The aqueous extracts of T. vogelii, C. lusitanica, and C. viminalis exerted the highest inhibition on seed germination. Globally, treated seeds responded differentially and gradually to germination. Forty percent of seeds quickly responded to the treatment with NAA and the number of germinated seeds increased with time up to 90%; with up to 10% of seed not germinating. Also, considering NAA and negative control, the germination pattern was the same in each experiment with up to 90 and 83% of seeds germinating, respectively.
Figure 2. Germination kinetics of tomato seeds in presence and absence of plant extract.
3.1.2. Effects of plant extracts and fractions on the tomato seedling diameter
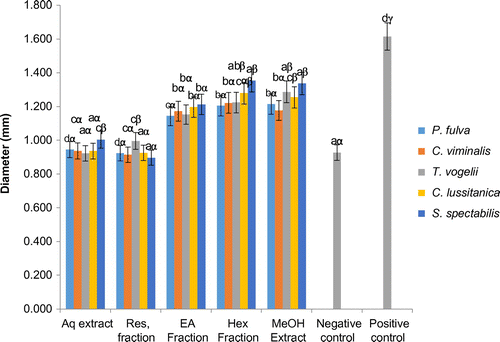
For all plant species, the methanol extract, hexane and ethyl acetate fractions induced significant increases in diameters of seedlings, while the negative control, aqueous extract and the residual fraction had comparable effects (p > 0.05) (Figure ). However, the positive control significantly increased (p < 0.05) diameter of seedlings compared to all the above. There was a significant increase in the diameter of seedlings by the methanol extract and hexane fractions of C. lusitanica and S. spectabilis.
Figure 3. Effects of plants extracts and fractions on the tomato seedling diameter at the end of the experiment (10 days).
a, b, c: for the same plant, extracts or fractions carrying different letters induced significant differences in seedlings diameters (Waller-duncan test, p < 0.05).
α, β, γ: for the same type of extracts or fractions, plant species carrying different letters induced significant differences in seedlings diameters (Waller-duncan test, p < 0.05).
3.1.3. Effects of plant extracts and fractions on the tomato seedling stem lenght
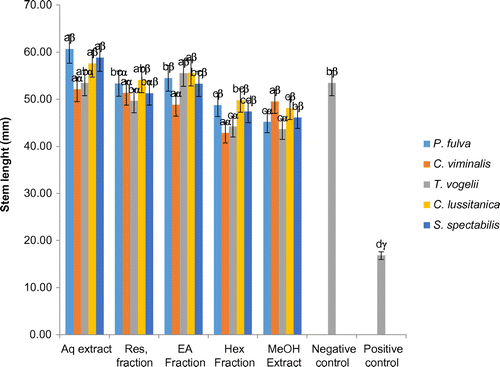
The activation or inhibition effect of the plant extracts and fractions on the seedling stem length is illustrated in Figure . Generally, the aqueous extracts which could not increase the diameter of seedlings were the most effective on the stem length. Although the methanol extracts generally induced a reduction in the stem length of seedlings relative to the negative control, the treatment of tomato seedlings with ethyl acetate fractions of P. fulva, T. vogelii, and C. lusitanica and residual fraction of P. fulva and C. lusitanica residue fractions generally led to increases in the stem length of seedlings.
Figure 4. Effects of plants extracts and fractions on the tomato seedling plant stem lenght at the end of the experiment (10 days).
a, b, c: for the same plant, extracts or fractions carrying different letters induced significant differences in seedlings stem lenght (Waller-duncan test, p < 0.05).
α, β, γ: for the same type of extracts or fractions, plant species carrying different letters induced significant differences in seedlings stem lenght (Waller-duncan test, p < 0.05).
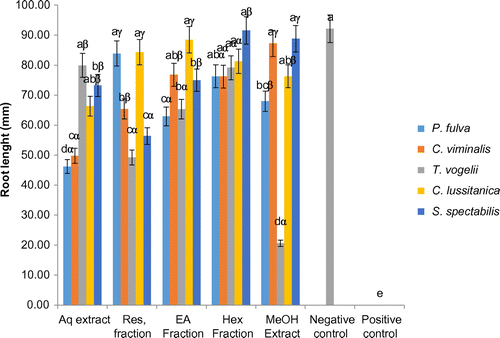
3.1.4. Effect of extracts and fractions on the seedling root lengths
The effect of the plant extracts/fractions on the root length of tomato seedlings after germination is presented in Figure . Treatments generally reduced the root length compared to negative control. For the methanol extracts, only those from C. viminalis and S. spectabilis preserved the root length, to a dimension comparable to that of the negative control. Hexane fractions, especially from T. vogelii significantly led to increases in the root length, though to a value below that of the negative control. The aqueous extracts from T. vogelii had the strongest activity on root length, very close to the negative control and significantly (p < 0.05) higher than those of T. vogelii methanol extract and fractions, but comparable to the effect of the Hexane fraction. The methanol extract of T. vogelii induced the strongest reduction in the root length but less than the positive control which completely inhibited the formation of roots.
Figure 5. Effects of plants extracts and fractions on the tomato seeding plant root lenght at the end of the experiment (10 days).
a, b, c: for the same plant, extracts or fractions carrying different letters induced significant differences in seedlings root lenght (Waller-duncan test, p < 0.05).
α, β, γ: for the same type of extracts or fractions, plant species carrying different letters induced significant differences in seedlings root lenght (Waller-duncan test, p < 0.05).
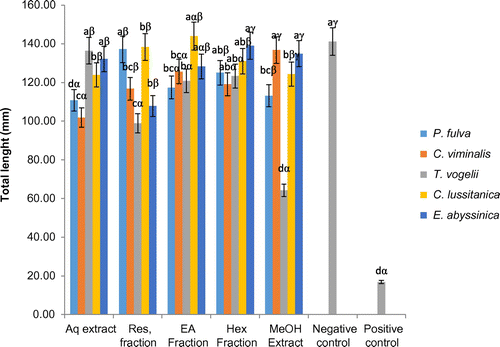
3.1.5. Effect of extracts and fractions on tomato seedling total length
The seedlings treated with the aqueous extracts of T. vogelii and S. spectabilis as well as the methanol extracts of C. viminalis and S. spectabilis did not show significant (p > 0.05) variation in total length compared to the negative control (Figure ). The resulting seedling lengths were close to 140 mm, while those of seedlings treated with the other extracts were significantly reduced. The fractions from T. vogelii, P. fulva and C. lusitanica methanol extracts had reduced total seedlings length compared to the control.
Figure 6. Effects of plants extracts and fractions on the tomato seedling plant total lenght at the end of the experiment (10 days).
a, b, c: for the same plant, extracts or fractions carrying different letters induced significant differences in seedlings total lenght (Waller-duncan test, p < 0.05).
α, β, γ: for the same type of extracts or fractions, plant species carrying different letters induced significant differences in seedlings total lenght (Waller-duncan test, p < 0.05).
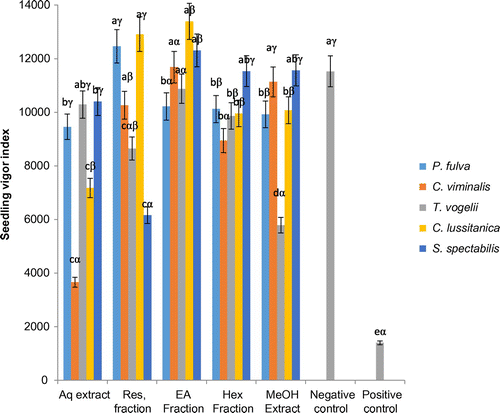
3.1.6. Effects of plants extracts and fractions on tomato seedling vigor index
The plant extracts and fractions exerted variable effects on tomato seedling plant vigor index (Figure ). Apart from T. vogelii, the vigor index of seedlings treated with methanol extracts were significantly (p < 0.05) higher than those treated with their corresponding aqueous extracts. The aqueous and methanol extracts from S. spectabilis were the most effective, inducing seedling vigor index of about 10,200 and 12,100 units, respectively. The methanol extract of S. spectabilis induced vigor index comparable to that of the negative control, while the rest of the extracts induced a reduction in the vigor index. Treatment of seedlings with ethyl acetate fractions led to significant increases in the vigor index.
Figure 7. Effects of plants extracts and fractions on the tomato seedling plant vigor index at the end of the experiment (10 days).
a, b, c: for the same plant, extracts or fractions carrying different letters induced significant differences in seedlings vigor index (Waller-duncan test, p < 0.05).
α, β, γ: for the same type of extracts or fractions, plant species carrying different letters induced significant differences in seedlings vigor index (Waller-duncan test, p < 0.05).
3.2. Discussion
Some plant extracts, particularly C. viminalis and S. spectabilis reduced the overall seed germination percentage indicating the presence of potential allelopatic inhibitory substances in such extracts. Reduction in germination is the most obvious allelopathic effect observed in bioassays (Alagesa Boopathi, Citation2011; Samreen, Hussain, & Sher, Citation2009) and in environment (ZhongQun, Junan, HaoRu, & Zhi, Citation2012). Thus, previous studies showed that different leaf aqueous extracts of C. arizonica showed significant inhibitory effects on germination percentage of tomato seeds (Aliloo, Citation2012). Likewise, germination efficiency of tomato seeds decreased after treatment with Eucalyptus camaldulensis extracts (Fikreyesus, Kebebew, Nebiyu, Zeleke, & Bogale, Citation2011).
In this study, solvent polarity effect was highly pronounced and the reduction in germination percentage was intensified with increasing extract polarity, suggesting an increase in active principles with polarity. The ethyl acetate fraction, less polar than aqueous and methanol fractions exhibited the highest stimulatory activities leading to a higher germination percentage than the negative control. Kiran, Lalitha, and Raveesha (Citation2011) showed that extract obtained using polar solvent could have phytotoxic effect on tomato seeds while Al-Wakeel, Gabr, Hamid, and Abu-El-Soud (Citation2007) suggested that it may possess allelochemicals with inhibitory effects on seed germination. This may be the case for the ethyl acetate fractions of the tested plants of this study, since it is known that allelochemicals present in some leaf extracts can prevent growth of embryo or cause its death (Anjana & Pramod, Citation2010). Different other mechanisms may explain the observed reduction in seed germination, including obstruction of water imbibition by the seeds (Tawaha & Turk, Citation2003), alteration in the synthesis or activities of gibberellic acid (Olofsdotter, Citation2001).
| • | The result on the seedling diameter indicates that stem-widening chemical compounds are concentrated in the hexane fractions, as well as in the ethyl acetate fractions to a lesser extent. Considering this active extract and its active fractions, S. spectabilis, C. lusitanica, and T. vogelii are more likely to induce such an effect. The same groups of compounds could be responsible for the stem-lengthening effects of the ethyl acetate fractions of P. fulva, T. vogelii, and C. lusitanica as well as P. fulva and C. lusitanica residue fractions. The increase in shoot length by these extracts/fractions corroborates the work of Perelló, Gruhlke, and Slusarenko (Citation2013), who demonstrated the same effect for garlic juice treatment on wheat cultivar as a function of garlic juice concentration tested. In a previous study, aqueous leaf extracts of C. lusitanica was shown to significantly reduce both germination and radicle growth of chickpea, maize, pea, and teff (Lisanework & Michelsen, Citation1993). | ||||
Overall, no plant extract accelerated seedling growth in length since all the values were less than or comparable to that of the negative control. Apart from the aqueous extract of T. vogelii and the MeOH extracts of C. viminalis and S. spectabilis, the rest of the extracts may possess plant length-inhibiting factors. Distilled water and methanol extracts are expected to contain polar substances, known to reduce the growth of seedling by inhibiting cell division or cell enlargement in the meristematic areas of shoot and root of plants and may also interfere with the rearrangement of microtubules during cell division (Singh, Pandey, & Singh, Citation2009). Consequently, these extracts, together with their residual fractions, caused the plants to have shorter shoots and roots, and thinner diameters. This may explain why the hexane fractions containing non-polar substances were generally the best fractions regarding increase in shoot and root lengths, and diameters. These fractions therefore have non polar growth-accelerating compounds (Sornchai, Saithong, Srichompoo, Unartngam, & Iamtham, Citation2014), probably in appreciable amounts to provoke visible length increase.
All the extracts, apart from the MeOH extract of S. spectabilis which induced comparable vigor index to that of the negative control, induced a reduction in the vigor indices. This shows that the vigor index-reducing extracts may contain inhibiting factors. It could be observed that most of the vigor index inhibitors were distributed, as a function of the plant extract, in the residue and hexane fraction, while the ethyl acetate fraction contained mostly vigor index activators.
4. Conclusion
The overall results of the present study show that the MeOH extracts of C. viminalis, T. vogelii, S. spectabilis, C. lusitanica, and P. fulva, as well as their ethyl acetate fractions, might contain compounds that can differentially affect positively or negatively tomato seed germination as well as seedling length and vigor index. Extract fractions could also contain both stimulatory and inhibitory allelochemicals, as related to the parameters measured. The stimulatory allelochemicals present in the ethyl acetate extract may have good biotechnological application potential in agriculture. It would be very interesting to identify such compounds for further investigations.
Additional information
Funding
Notes on contributors
Kuiate Jules-Roger
Our main research domain is the physiology of growth and development of the plant, ripening and senescence of fruits. Events occurring in the field greatly influence these processes. The conventional cultivation methods of plant like tomato require the use of important quantities of chemical inputs, which can impair the nutritional virtues of fruits and harm the environment. Thus, an integrated management of the production of fruits with known health-promoting properties should involve pre-harvest treatments based on biological material. In this regard, many of our research activities in the past years have been focused on bioregulators/biostimulants, emerging notions that refer to any biological organism, its powder or extracts that can induce positive alterations in plant growth and development parameters. The use of allelochemicals in the cultivation of tomatoes is one of pragmatic approaches to reduce biotic and abiotic stresses effects, enhance growth and yield, and preserve fruit quality.
References
- Afzal, I., Munir, F., Ayub, C., Basra, S., Hameed, A., & Nawaz, A. (2009). Changes in antioxidant enzymes, germination capacity and vigour of tomato seeds in response of priming with polyamines. Seed Science and Technology, 37, 765–770.10.15258/sst
- Alagesa Boopathi, C. (2011). Allelopathic effects of Andrographis paniculata Nees on germination of Sesamum indicum L. Asian Journal of Experimental Biological Sciences, 2, 147–150.
- Aliloo, A. (2012). Allelopathic potentials of Cupressus arizonica leaves extracts on seed germination and seedling growth of Lolium perenne and Poa pratensis. International Journal of Agriculture and Crop Sciences, 4, 1371–1375.
- Al-Wakeel, S. A. M., Gabr, M. A., Hamid, A. A., & Abu-El-Soud, W. M. (2007). Allelopathic effects of Acacia nilotica leaf residue on Pisum sativum L. Allelopathy Journal, 19, 411–422.
- Anjana, D., & Pramod, K. J. (2010). Seed Germination responses of the medicinal herb Centella asiatica. Brazilian Journal of Plant Physiology, 22, 143–150.
- Badmus, A., & Afolayan, A. (2012). Allelopathic potential of Arctotis arctotoides (L.f.) O. Hoffm aqueous extracts on the germination and seedling growth of some vegetables. African Journal of Biotechnology, 11, 10711–10716.
- Dayan, F. E., Romagni, J. G., & Duke, S. O. (2000). Investigating the mode of action of natural phytotoxins. Journal of Chemical Ecology, 26, 2079–2094.10.1023/A:1005512331061
- FAOSTAT. (2013). Annuaires statistiques de la FAO – L’alimentation et l’agriculture dans le monde. Retrieved consulted on the 18 March 2016 from http://www.factfish.com/statistic-country/cameroon/tomatoes,+production+quantity
- Fikreyesus, S., Kebebew, Z., Nebiyu, A., Zeleke, N., & Bogale, S. (2011). Allelopathic Effects of Eucalyptus camaldulensis dehnh. on germination and growth of tomato. American-Eurasian Journal of Agricultural & Environmental, 11, 600–608.
- Kiran, B., Lalitha, V., & Raveesha, K. A. (2011). In vitro evaluation of aqueous seed extract of Psoralea corylifolia L. on seed germination and seedling vigor of maize. Journal of Applied Pharmaceutical Science, 1, 128–130.
- Kuzyakov, Y., Hill, P. W., & Jones, D. L. (2007). Root exudate components change litter decomposition in a simulated rhizosphere depending on temperature. Plant Soil, 290, 293–305.10.1007/s11104-006-9162-8
- Lisanework, N., & Michelsen, A. (1993). Allelopathy in agroforestry systems: The effects of leaf extracts of Cupressus lusitanica and three Eucalyptus spp. on four Ethiopian crops. Agroforestry Systems, 21, 63–74.10.1007/BF00704926
- Olofsdotter, M. (2001). Rice—A step toward use of allelopathy. Agronomy Journal, 93, 3–8.10.2134/agronj2001.9313
- Patel, R. G., & Manhad, A. U. (2014). Effect of Gibberillins on seed germination of Tithonia rotundifolia Blake. International Journal of Innovative Research in Science, Engineering and Technology, 3, 10680–10684.
- Perelló, A., Gruhlke, M., & Slusarenko, A. (2013). Effect of garlic extract on seed germination, seedling health, and vigor of pathogen-infested wheat. Journal of Pharmacy Practice and Research, 53, 317–323.
- Ruilong, W., Yang, X., Song, Y., Zhang, M., Su, Y., & Rensen, Z. (2011). Allelopathic potential of Tephrosia vogelii Hook. f.: Laboratory and field evaluation. Allelopathy Journal, 28, 53–62.
- Samreen, U., Hussain, F., & Sher, Z. (2009). Allelopathic potential of Calotropis procera Ait. Pakistan Journal of Plant Science, 15, 7–14.
- Singh, N. B., Pandey, B. N., & Singh, A. (2009). Allelopathic effects of Cyperus rotundus extract in vitro and ex-vitro on banana. Acta Physiologiae Plantarum, 31, 633–638.10.1007/s11738-009-0274-7
- Sornchai, A., Saithong, N., Srichompoo, Y., Unartngam, A., & Iamtham, S. (2014). Effect of Spirulina maxima aqueous extract on seed germination and seedling growth of mung bean, Vignaradiate and rice, Oryza sativa var. Japonica.. Journal of the International Society for Southeast Asian Agricultural Sciences, 20, 77–84.
- Tawaha, A., & Turk, M. (2003). Allelopathic effects of black mustard (Brassica nigra) on germination and growth of wild barley (Hordeum spontaneum). Journal of Agronomy and Crop Science, 189, 298–303.10.1046/j.1439-037X.2003.00047.x
- Tonfack, L. B., Bernadac, A., Youmbi, E., Mbouapouognigni, V. P., Ngueguim, M., & Akoa, A. (2009). Impact of organic and inorganic fertilizers on tomato vigor, yield and fruit composition under tropical andosol soil conditions. Fruits, 64, 167–177.10.1051/fruits/2009012
- ZhongQun, H., Junan, Z., HaoRu, T., & Zhi, H. (2012). Different vegetables crops in response to Allelopathic of hot pepper root exudates. World Applied Sciences Journal, 19, 1289–1294.