
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
It is well known that fruit nuts contain wide variety of flavonoids and various proteins, consumption of which has been associated with the reduced risk of chronic diseases. Cystatins, a family of cysteine proteinase inhibitors, ubiquitously present in all cells serve various important and critical physiological functions. In this study a phytocystatin with molecular mass of 63.4 kDa was purified to homogeneity by a three-step process including ammonium sulfate fractionation (50–70%), acetone precipitation, and gel filtration chromatography on Sephacryl S100-HR column. The purified inhibitor migrated as single band under native and SDS-PAGE. The Ki values for purified inhibitor with papain, ficin, and bromelain were found to be 45.45, 83.33, and 90.9 nM, respectively, suggesting higher affinity of the inhibitor for papain as compared to ficin and bromelain. Phytocystatin was stable in broad pH and temperature range. Purified cystatin appeared to be antigenic as observed in western blot analysis. ITC assay data show a binding stoichiometry of 0.870 ± 0.03 sites for cystatin and papain interaction which indicated that cystatin is surrounded by nearly one papain molecule. FTIR, UV, and fluorescence studies showed significant conformational changes on cystatin–papain complex formation. Purified cystatin was found to possess 36.8% α-helical content as observed by CD spectroscopy.
Public Interest Statement
Plant cystatins or phytocystatins are the second most studied class of protease inhibitors. They are cysteine (thiol) protease inhibitors that interact with the active site of target proteases and block access to their protein substrates. Since the publication of a first full paper reporting their existence, phytocystatins have shown wide application in pharmaceutical, agricultural, and industrial field. These plant derived inhibitors are also gaining wide consideration in drug designing and engineering GM crops. Owing to the great significance and importance of these inhibitors, it is essential to explore and isolate them from various plant sources. In the present study, a phytocystatin has been isolated and purified from almond which is a source of high-quality protein with great nutritive value. Almond cystatin was purified to homogeneity by a three-step process including ammonium sulfate precipitation and gel filtration chromatography. Purified inhibitor was further characterized using various biophysical and biochemical techniques.
Competing Interests
The authors declare no competing interest.
1. Introduction
Proper functioning of the living system requires an appropriate equilibrium between free cysteine proteases and their complexes with inhibitors. Proteases are omnipresent in all micro and macro-organism and involved in health and welfare by performing an essential role in survival and maintenance of the organism (Dunaevsky, Elpidina, Vinokurov, & Belozersky, Citation2005; Haq, Atif, & Khan, Citation2004; Hartl, Giri, Kaur, & Baldwin, Citation2011; Joanitti, Freitas, & Silva, Citation2006; Lawrence & Koundal, Citation2002; Mosolov & Valueva, Citation2008; Neurath, Citation1989; Supuran, Scozzafava, & Clare, Citation2002; Valueva & Mosolov, Citation2004). In spite of many advantages of these proteases, their activity has to be closely regulated and controlled to avoid the excess activity of these proteases, as it may possibly damage its host organism. This controlling task is carried out by PIs (protease inhibitors) (Rawlings, Tolle, & Barrett, Citation2004). Cystatins (PIs) are the natural regulators of cysteine protease activities. Phytocystatins are plant proteinaceous inhibitors and have been identified in both monocot and dicot species (Lin et al., Citation2006). These small proteins specifically inhibit the action of cysteine proteases in general and of the papain class in particular. The cystatin superfamily comprises three classes of inhibitors namely stefins, cystatins, and kininogens (Abe, Kondo, Watanabe, Emori, & Arai, Citation1991), exclusively based on the studies on cystatins of animal origin. In plants they are named as phytocystatin and are particularly well characterized in some crops such as potato (Nissen et al., Citation2009), taro (Yang & Yeh, Citation2005), sesame (Shyu, Chou, Yiu, Lin, & Tzen, Citation2004), strawberry (Martinez et al., Citation2005) turmeric (Chan, Abu Bakar, Mahmood, Ho, & Shaharuddin, Citation2014), kiwifruit cortex and seed (Rassam & Laing, Citation2004), and Chinese cabbage (Hong et al., Citation2012). However, in potato tuber cysteine proteinase inhibitors, prominent structural differences have been found, which include an 80 kDa cystatin with eight cystatin domains (Gruden et al., Citation1997; Waldron, Wegrich, Ann Owens Merlo, & Walsh, Citation1993).
Although the primary sequences of phytocystatins are more similar to the type II cystatins of animals, they are assigned to an independent family (Margis, Reis, & Villeret, Citation1998). On the basis of molecular weight, they have been divided into three distinct groups (Chu, Liu, Wu, Yeh, & Cheng, Citation2011; Margis-Pinheiro, Zolet, Loss, Pasquali, & Margis, Citation2008; Wang et al., Citation2008). Most phytocystatins are 12–16 kDa in size, contain no disulfide bonds, and show significant amino acid sequence identity among them and to the cystatins family of animal origin (Barrett, Citation1987). The group two phytocystatins are approximately or greater than 23 kDa, such as those found in taro (Yang & Yeh, Citation2005), sesame (Shyu et al., Citation2004), and strawberry (Martinez et al., Citation2005). They have a highly conserved N-terminal region, which is similar to first group and are tailed by a repetitive peptide at the C-terminus, in which variation is possibly caused by gene duplication (Christeller, Citation2005). The third group of phytocystatins, is found in potato (Waldron et al., Citation1993) and tomato (Wu & Haard, Citation2000), and includes high molecular weight multi-cystatin domains.
Almonds are one of the most popular tree nuts on a worldwide basis and rank number one in tree nut production. Almonds are a good source of high-quality protein and contain 16–22% protein on a dry weight basis (Sathe et al., Citation2002). It has been reported that almonds, when incorporated in the diet, reduce colon cancer risk in rats (Davis & Iwahashi, Citation2001). It also increases HDL cholesterol and reduces LDL cholesterol levels in humans (Hyson, Schneeman, & Davis, Citation2002). Almonds also have interesting biological properties such as sedative, anti-inflammatory, anti-hyperlipidemic, anti-tumoral, and antioxidant activities (Esfahlan, Jamei, & Esfahlan, Citation2010).
Considerable amount of work has been done on seed cystatins of herbaceous crops (e.g. cereals and legumes), while little is known about cystatins from other plant sources, especially as those of large-seeded trees. These plants are particularly found in tropical forest and their seeds have relatively high water contents at the time of shedding, and these seeds remain in the soil for long time before germination. It may be expected that such seeds display defense barriers towards pests and pathogens as well as characteristic mechanisms for protein turnover regulation (Pernas, Sánchez-Monge, Gómez, & Salcedo, Citation1998). Cystatins and endogenous cysteine proteases show different expression patterns during seed maturation and germination, which recommends that cystatins are involved in the regulation of protein turnover during such processes (Abe, Abe, Kuroda, & Arai, Citation1992; Belenghi et al., Citation2003; Fernandes, Campos, Do Val, & Xavier-Filho, Citation1991). On the other hand, the effects of cystatins on exogenous proteases, speculate its defensive function, such as those of insect pests and nematodes, together with their inducible expression by wounding or methyl jasmonate (Botella et al., Citation1996; Zhao et al., Citation1996).
Plant cystatins have important roles to play in plant system owing to their cysteine proteinase inhibitory activity. Earlier numerous spectroscopic and kinetic studies have been carried out to understand the mechanism of inhibition of cysteine protease by cystatins. In the current study, a phytocystatin from almond has been purified with better yield and fold purification by slightly modifying the method reported earlier (Siddiqui, Sohail, Bhat, Rehman, & Bano, Citation2015). Detailed physiochemical characterization was further carried out using various biophysical techniques. Isothermal titration calorimetry along with kinetic studies and other spectroscopic techniques further helped to explain its interaction with papain.
2. Materials and methods
2.1. Materials
Papain, Sephacryl S-100HR, cysteine, casein, coommassie brilliant blue G-250, were purchased from Sigma Chemical Co, USA. Molecular weight markers were obtained from Genei, India. All other chemicals used were of analytical grade.
2.2. Purification of almond cystatin
Phytocystatin from almond was purified using a slightly modified method of Wu and Haard (Citation2000). Overnight soaked almonds (100 gm) in 300 mL of extraction buffer (50 mM sodium phosphate buffer pH 7.5, 3 mM EDTA, and 0.15 M sodium chloride) were homogenized and centrifuged at 8000 rpm for 15 min in a sigma cooling centrifuge. Pellets were discarded. The supernatant was collected and fractionated by 50–70% ammonium sulfate saturation. The precipitate was collected by centrifugation at 10,000 rpm for 30 min at 4°C and dissolved in minimum amount of 50 mM sodium phosphate buffer pH 7.5. The precipitated protein was then extensively dialyzed against sodium phosphate buffer (pH 7.5) at 4°C to remove ammonium sulfate. Dialyzed sample was treated with chilled acetone in the ratio of 1:1 and centrifuged at 8,000 rpm for 20 min. The supernatant was collected and applied on Sephacryl S-100HR gel-filtration column. Fractions of 5 mL were collected. Fractions having maximum protein peak and cysteine proteinase inhibitory activity were pooled and concentrated for further study. The purification of almond cystatin was repeated several times to obtain the fresh inhibitor for each experiment.
2.3. Papain inhibitory activity assay and protein estimation
The inhibitory activity of cystatin was assessed by its ability to inhibit the caesinolytic activity of papain by the method of Kunitz (Citation1947). Assay involved pre-activation of papain in 0.2 mM cysteine and 0.15 mM EDTA at pH 7.5 in 50 mM sodium phosphate buffer. The activated papain was incubated with 0.3 mL of the purified inhibitor for 30 min at 37°C. Final reaction volume was 1 mL. The reaction was initiated by addition of 1 mL of 2% casein solution to the reaction mixture and incubated for 30 min at 37°C.The reaction was terminated by adding 1 mL of trichloroacetic acid (10%) and particulate matter was removed by centrifugation at 2,500 rpm for 10 min. The soluble peptides were analyzed by the method of Lowry, Rosebrough, Farr, and Randall (Citation1951). The protein content was quantitated by Folins phenol reagent by the method of Lowry et al. (Citation1951). A control was also prepared that was devoid of the purified phytocystatin.
Inhibitory activities of almond cystatin at different concentration (0–50 μg) were also analyzed for other cysteine proteases like ficin, bromelain, and serine proteases (trypsin, chymotrypsin).
2.4. Carbohydrate and thiol group estimation
Carbohydrate content was estimated by the method of DuBois, Gilles, Hamilton, Rebers, and Smith (Citation1956), while thiol group was determined by Ellman’s method (Citation1959). Glucose and dithionitrobenzoate (DTNB) were used as standard for these methods, respectively.
2.5. Stability of almond cystatin
The thermal stability of almond cystatin (50 μg) in 50 mM phosphate buffer, pH 7.5 was investigated at different temperatures (4–90°C) for 60 min. These samples were rapidly cooled in ice cold water bath and the remaining inhibitory activity was determined against 50 μg of papain by the routine papain assay. pH stability of the almond cystatin was investigated by incubating the samples from pH 3–12 at 4°C for 4 h. Sodium acetate buffer (50 mM) was used for pH 3–6 (adjusting pH with 1 N HCl or NaOH). Sodium phosphate buffer (50 mM) was used for pH 7–8 and Tris-HCl buffer (50 mM) was used for pH 9–12. The remaining inhibitor activity of the purified cystatin was analyzed with casein as a substrate after adjusting the pH to 7.5 with 50 mM phosphate buffer.
2.6. Immunological properties
About 300 μg of purified almond cystatin was emulsified in equal volume of Freund’s complete adjuvant and injected subcutaneously in healthy male albino rabbit. Two weeks later 150 μg of purified almond cystatin was injected along with Freund’s incomplete adjuvant to raise antiserum. The injection was repeated every week and the rabbit was bled every second week. The collected blood was allowed to stand and coagulate at room temperature for 3 h. De-complementation of the anti-sera was performed by incubation at 56°C for 30 min. The de-complemented sera were then stored at −20°C. Immunodiffusion and cross-reactivity was performed as reported earlier (Rashid, Sharma, & Bano, Citation2006). Chick pea cystatin (Bhat, Sohail, Siddiqui, & Bano, Citation2014) and buffalo heart cystatin (Sohail et al., Citation2015) used were also isolated and purified in our laboratory.
2.7. Direct binding ELISA
Antibodies generated against specific cystatin antigen in the sera of almond cystatin immunized rabbits were measured by the technique of direct binding ELISA as reported earlier by Rashid et al. (Citation2006).
2.8. Western blot analysis
Increasing concentration (30–50 μg) of purified protein was run on 10% SDS-PAGE and then transferred to a nitrocellulose membrane for 2 h. The membrane was then blocked by 5% milk in PBS and incubated for 1 h. Membrane was further incubated overnight in the presence of primary antibody. After washing with PBST, membrane was incubated with goat anti-rabbit IgG at 1∶5000 dilution for 2 h (Santa Cruz, CA, USA). Blot was further developed using ECL system.
2.9. Stoichiometry of cystatin–papain complex
Stoichiometry of cystatin–papain complex was determined by titration of papain activity against various concentrations of almond cystatin (0.1–1.2 μM) with casein as a substrate in 50 mM phosphate buffer, pH 7.5 containing 20 mM EDTA, and 50 mM L-cysteine as activating agent. The inhibitory activity of cystatin was assessed by its ability to inhibit caseinolytic activity of papain by the method of Kunitz (Citation1947).
2.10. Inhibition constant (Ki) determination
Inhibition constant of the purified cystatin binding to papain was determined by titration of a fixed amount of papain (0.06 nM) against different almond cystatin concentrations (0.06, 0.12, 0.18, 0.24, and 0.30 nM) using various concentration of casein as substrate (0.132, 0.263, 0.526, and 0.789 nM). The remaining papain activity was assayed in 50 mM sodium phosphate buffer, pH 7.5. Lineweaver–Burk 1/v versus 1/[s] was plotted, the km was calculated for each concentration of inhibitor and secondary plot was obtained to determine Ki for the inhibitor.
2.10.1. Determination of dissociation rate constant k−1
The condition for maximal association between the papain and almond cystatin was accomplished before the reaction was made to shift towards dissociation by increasing the concentration of substrate in excess. Excess substrate will bind to the free enzyme. Substrate induced dissociation was observed with an identical enzyme–inhibitor complex incubated for 30 min at 37°C. Excess substrate 6% of the casein was added to the mixture at different time intervals (0, 5, 10, 15, 20, 25, and 30 min), and then the residual activity was assayed.
2.10.2. Determination of association rate constant k+1
The association rate constant was determined by the relation (Abrahamson, Barrett, Salvesen, & Grubb, Citation1986).(1)
(1)
Half-life of the complexes were calculated by rearranging the equation(2)
(2)
2.11. Electrophoresis
2.11.1. Native PAGE
Native PAGE was run to check the homogeneity of the purified cystatin. Cystatin homogeneity was checked by native gel (10% polyacrylamide) using the method of Laemmli (Citation1970). Crude sample with total protein was run along with purified cystatin to check the purity of sample. Different samples (protein obtained from different fraction having maximum inhibitory activity against papain) containing 40 μg of protein mixed with equal volume of sample buffer (62.5 mM Tris HCl pH 6, 8, 10% (v/v) glycerol and 0.001% bromophenol) as tracking dye was applied to the wells. Electrophoresis was performed at 100 V in the electrophoresis buffer containing 192 mM glycine and 25 mM Tris-HCl pH 6.8 until the tracking dye reached the bottom of the gel. The gels were stained with 0.1% Coomassie Brilliant Blue R-250.
2.11.2. Two-dimensional electrophoresis
The purified almond cystatin was analyzed by two-dimensional gel electrophoresis essentially as reported (Fountoulakis & Gasser, Citation2003). Protein sample dissolved in sample buffer was centrifuged and applied on IPG pH 3–10 (Bio Rad, USA) nonlinear strips. Focusing started at 200 V and the voltage was gradually increased to 5,000 V at 3 V/min and kept constant for a further 24 h (approximately 180,000 kVh totally). After focusing, the IPG strips were equilibrated for 15 min in reducing solution containing 30% v/v glycerol, 6 M urea, 1% dithiothreitol (DTT), followed by equilibration in alkylating buffer consisting of 30% v/v glycerol, 6 M urea, and 4% iodoacetamide. Afterward, the IPG strip was transferred onto a linear 10% SDS polyacrylamide gel and electrophoresis was performed in presence of protein molecular weight marker at a constant voltage of 100 V. Gels were stained with Coomassie brilliant blue R-250.
2.11.3. Molecular weight determination by SDS-PAGE
The molecular weight of purified almond cystatin under denaturing condition was determined by SDS-PAGE (reducing and non-reducing) by the procedure of Weber and Osborn (Citation1969). The mobilities of marker proteins determined under identical conditions were plotted against the logarithms of molecular weight. The analysis of the data indicated a linear relationship between log M and relative mobility (Rm) and the plot was used in calculating the molecular weight of purified cystatin.
2.12. Molecular weight determination by gel filtration chromatography
The molecular weight of purified almond cystatin was computed from its elution volume on a Sephacryl S 100-HR column (60 × 1.7 cm). The column was calibrated by determining the elution volume of some marker proteins–phosphorylase b (97.4 kDa), bovine serum albumin (68 kDa), ovalbumin (45 kDa), soyabean trypsin inhibitor (20.1 KDa), lysozyme (14.3 kDa) cytochrome c (12 kDa). These data were analyzed according to the theoretical treatment by the method of Andrews (Citation1964). The linear plot between Ve/Vo and log M was used for calculating the molecular weight of almond cystatin where Ve is the elution volume of the protein and Vo is the void volume of the column determined using blue dextran.
2.13. Spectral analysis
2.13.1. UV–vis absorption spectroscopy
UV–vis absorption studies were employed to obtain the spectra of cystatin (80 μg/3 ml) in the absence and presence of activated papain. Spectra were obtained using Shimadzu spectrophotometer using a cuvette of 1.0-cm path length from 250 to 310 nm. Appropriate controls of the solvent buffer were run and corrections were made wherever necessary. Results are mean of three experiments.
2.13.2. Fluorescence spectroscopy
The emission spectra of thiol protease inhibitor (almond cystatin) (40 μg/1.5 ml), papain and cystatin–papain complex was taken after excitation at 280 nm on a Shimadzu spectrofluorometer at 25°C. Emission wavelength range was of 300–400 nm. Cells with 1-cm path length were used and samples were continuously stirred during measurements. Corrected emission spectra were recorded with an excitation and emission band width of 10 nm. Appropriate controls were run and corrections were made wherever necessary.
2.13.3. Circular dichroism measurement
CD measurements were carried out on a JASCO-J-815 spectropolarimeter equipped with a Peltier-type temperature controller. Far-UV CD spectra of cystatin in the absence and presence of papain was taken at protein concentration of 20 μM in 0.1-cm path length cell. All the spectra were corrected for the appropriate blanks. The observed ellipticity is converted to mean residual ellipticity [θ] in degrees square centimeter per decimole using the following equation.(3)
(3)
where θobs is the observed ellipticity in mdeg, n is the total number of amino acid residues in the protein, c is the molar concentration of the protein, and l is the path length in centimeters. Helical content of cystatin was calculated from the mean residue ellipticity (MRE) values at 222 nm using the following equation as described by Chen, Yang, and Martinez (Citation1972).(4)
(4)
2.13.4. Fourier transform infrared spectroscopy
FTIR spectroscopy is a measurement of wavelength and intensity of the absorption of IR radiation by a sample. Infrared spectroscopy was performed to see the secondary structure present in almond cystatin. The FTIR measurement was carried out on Perkin Elmer-FTIR (PerkinElmer Spectrum-100, USA) in the range of 1520–1740 cm−1 at RT. FTIR spectra of cystatin alone and cystatin with papain was recorded. Each spectrum was the average of five scans. The changes in the peak frequency and intensity were then assigned to the conformational change within the cystatin.
2.14. Isothermal titration calorimetric measurements
The almond cystatin was titrated with papain using a VP-ITC titration microcalorimeter (MicroCal Inc., Northampton, MA) to find out the energetics of the binding. Prior to the titration experiment, all samples were degassed in a thermovac. The sample cell of the calorimeter was loaded with cystatin solution (20 μM) in sodium phosphate buffer (50 mM, pH 7.5) and reference cell with 50 mM sodium phosphate buffer pH 7.5. Multiple injections of 10 μl of papain solution (2.0 mM) were administered into the sample cell that contained cystatin. Each injection was administered over 20 s, with an interval of 180 s between successive injections. The reference power and stirring speed were set at 13 μcal s−1 and 307 rpm, respectively. The binding enthalpy change (∆H), entropy change (∆S), and the binding stoichiometry (N) were permitted to float during the least-square minimization process and taken as the best-fit values.
2.14.1. Statistical analysis
Every experiment was performed at least three times in order to check the reproducibility of the results. Data have been expressed as mean ± SEM. Statistical analysis was performed with one-way ANOVA software. Data were considered significant at p ≤ 0.05.
3. Results and discussion
3.1. Purification of almond cystatin
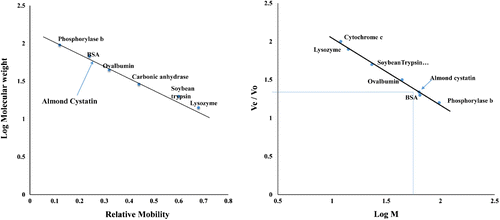
Numerous inhibitors of cysteine proteases have been described and isolated from a variety of sources (Kouzuma et al., Citation1996; Wu & Haard, Citation2000). In this study, purification of almond cystatin was achieved using a three-step procedure that included ammonium sulfate fractionation, acetone precipitation, and gel filtration chromatography (Sephacryl S-100HR). The fractions corresponding to protein peak with significant inhibitory activity were pooled and lyophilized for further analyses. This procedure is very efficient and simple and gives a percent yield and fold purification of 52.7% and 302.4, respectively, which is better than other reported procedures (Khan & Bano, Citation2009; Sadaf, Shahid, & Bilqees, Citation2005; Sumbul & Bano, Citation2006) (Table ). The method reported here is the updated and improved form of our previously reported procedure (Siddiqui et al., Citation2015) giving high yield and fold purification. Purified cystatin was found to be homogenous in native PAGE as well as two-dimensional gel electrophoresis (Figure ). It was found to be monomeric in nature showing a single band on SDS-PAGE under non-reducing and reducing conditions with a molecular mass of 63.4 kDa as reported earlier by our team (Siddiqui et al., Citation2015). Molecular weight was calculated by plotting log M against the relative mobility of different protein markers using Weber and Osborn method (Figure (A)). Molecular weight was further confirmed by passing almond cystatin and different protein markers through gel filtrations column (Figure (B)). Cysteine protease inhibitors purified from various other plant sources are usually small molecular mass protein (molecular weight less than 50 kDa) but some high molecular weight dimeric and tetrameric protease inhibitors from potato (Waldron et al., Citation1993) and tomato (Bolter, Citation1993; Jacinto, Fernandes, Machado, & Siqueira-Júnior, Citation1998) have also been isolated. Some animal cystatin with high molecular weight have also been isolated, e.g. goat lung (Khan & Bano, Citation2009), skin of Atlantic salmon (Synnes, Citation1998), goat brain (Sumbul & Bano, Citation2006).
Table 1. Purification of phytocystatin from almond
Figure 1. (A) Native-PAGE of total protein (lane 1) and purified almond cystatin (lane 2). (B) Native PAGE of purified almond cystatin. Each lane contains protein obtained from different fraction having maximum inhibitory activity against papain. (C) 2D gel electrophoresis of purified inhibitor with protein molecular weight markers.
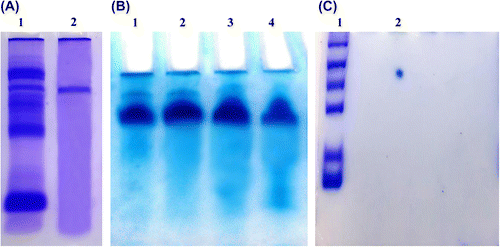
Figure 2. (A) Log M of marker proteins was plotted against relative mobility (Rm) of molecular weight markers for determination of molecular mass of almond cystatin. Molecular weight of standard marker proteins is A, Phosphorylase b (97.4 kDa); B, Bovine Serum Albumin (66 kDa); C, Ovalbumin (43 kDa); D, Carbonic Anhydrase (29 kDa); E, Soyabean Trypsin Inhibitor (20.1 kDa); F, Lysozyme (14.3 kDa). (B) Molecular weight determination of purified almond cystatin using Sephacryl S-100 HR gel filtration chromatography. Purified almond cystatin was applied on a column of Sephacryl S-100 HR (60 × 1.7 cm) and eluted with 50 mM sodium phosphate buffer, pH 7.5 at a flow rate of 15 ml h−1. The molecular weight markers used were, cytochrome c (12 kDa), lysozyme (14.3 kDa) soyabean trypsin inhibitor (20 kDa), ovalbumin (45 kDa), BSA (68 kDa), and phosphorylase b (97.4 kDa). Arrow shows the position of almond cystatin elution.
3.2. Carbohydrate and thiol content of purified almond cystatin
Carbohydrate content in almond cystatin was only 0.04% with no sulfhydryl content which is in accordance with the results reported previously (Amin, Ali Khan, Jahan Rizvi, & Bano, Citation2011; Wu & Haard, Citation2000).
3.3. Interaction of cystatin with different proteases
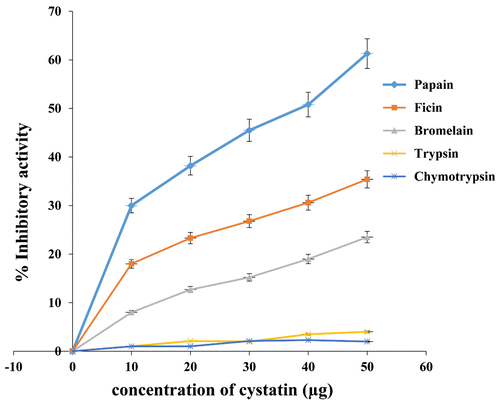
Inhibitory activities of almond cystatin were also analyzed for other cysteine proteases like ficin, bromelain, and serine proteases (trypsin, chymotrypsin). Purified cystatin showed inhibitory activity against cysteine proteases only (papain, ficin, bromelain). However, it was found to be inactive towards serine proteases like trypsin and chymotrypsin (Figure ). The data again confirm it to be a thiol protease inhibitor.
Figure 3. Inhibitory activity of almond cystatin for different proteases.
3.4. Stoichiometry of cystatin interaction with papain
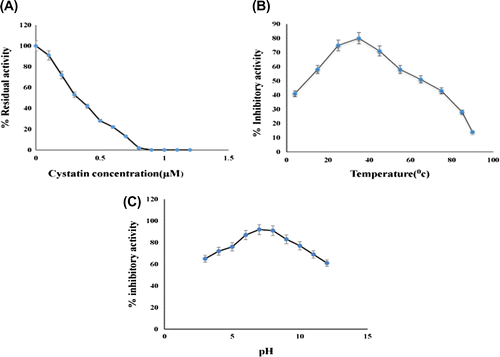
Protein inhibitory assay shows that on increasing papain and casein concentrations, inhibition occurs up to certain level, while beyond 0.6 μM of papain concentration, O.D. remains almost same. As evident from the plot with cystatin concentration on X-axis and % residual activity on Y-axis, it can be inferred that the amount of purified almond cystatin, required to inhibit 0.6 μM of papain activity completely was calculated to be 0.8 μM approximately (Figure (A)). These data indicate that 1 mol of almond cystatin contains almost one binding site for papain.
Figure 4. (A) Stoichiometric titration of papain with almond cystatin. Residual activity of papain was determined using casein as a substrate. (B) Effect of temperature on inhibitory activity of almond cystatin. 50 μg of cystatin in 50 mM sodium phosphate buffer, pH 7.5, was incubated at various temperatures for 60 min. Remaining % inhibitory activity was analyzed against 50 μg of papain. (C) Effect of pH on inhibitory activity of almond cystatin. 50 μg of cystatin was incubated in 50 mM sodium acetate buffer pH 3–6, sodium phosphate buffer, pH 7–8 and Tris-HCl buffer pH 9–12 at 37°C. Remaining % inhibitory activity was analysed against 50 μg of papain 37°C.
3.5. Stability of cystatin
The stability of almond cystatin was assessed by incubating the purified cystatin for 60 min at a wide range of temperatures varying between 4 and 90°C. As seen in Figure (B), with increasing temperature, the inhibitory activity of cystatin gradually increases up to 35°C. On further increasing the temperature, considerable decrease in the activity was observed with minimum activity at 90°C.
Cystatin stability studies carried out at different pH systems for a period of 4 h shows that the protease inhibitor had stability over a wide range of pH. From the results (Figure (C)), it is clear that the activity of the inhibitor gradually increases from pH 3 and reaches its maximum at pH 7.5 and after that gradually decreases up to pH 12. At the acidic (pH 3.0) and alkaline (pH 12.0) conditions, the inhibitory activities were sharply declined. Further at high alkaline and high acidic conditions the protease inhibitor was found to be unstable. These results of temperature and pH are comparable with other reported cystatins (Sharma, Rashid, & Bano, Citation2006).
3.6. Immunological properties
Purified cystatin as an antigen causes good immune response and resulting serum gives a titer value of 35481.33 as determined by the direct binding ELISA. The high antibody titer suggests that the cystatin used as antigen is of high molecular weight range (Lehninger, Nelson, Farr, & Cox, Citation2000; Rashid, Sharma, & Bano, Citation2005).
The antibodies raised against purified cystatin gave a reaction of identity with the cystatin as indicated by a single precipitin line on immunodiffusion suggesting that the wells contained both immunogenically and homogeneously pure almond cystatin (Figure (A)).
Figure 5. (A) Ouchterlony immunodiffusion: Antibodies carrying antiserum against almond cystatin was raised in rabbits. For the immunodiffusion study, the antiserum is allowed to react with cystatin on agarose plates. The central well contained the antiserum, whereas the surrounding three wells contained almond cystatin (A) 30 μg, (B) 40 μg, and (C) 50 μg. (B)Western immunoblot of the purified almond cystatin showed prominent antigenic bands. Lane a, b, c, d, e, f shows different concentrations of purified cystatin used.

Purified fraction was subjected to western blotting after SDS-PAGE. The western blot analysis revealed an antigenic polypeptide in the lane b (63 kDa) (Figure (B)). The western blot analysis of the purified almond cystatin with sera of rabbit immunized with almond cystatin showed prominent reactivity. Purified fractions from the Sephacryl S-100HR column reacted positively in indirect ELISA.
3.7. Kinetic studies
To determine the Ki value of the cystatin it was observed that inhibition of substrate hydrolysis occurred at very low concentration of protease inhibitor and the Ki calculated was found to be 45.45 nM under the assay conditions for papain. Ki for ficin and bromelain was 83.33 nM and 90.9 nM, respectively. The Ki value was the lowest for papain; hence it has the highest affinity for the inhibitor. Lineweaver–Burk plot shows that almond cystatin is an effective non-competitive inhibitor of papain (Figure ). Other phytocystatins such as corn, barley seeds, soybean, and tomato (Abe, Abe, Iwabuchi, Domoto, & Arai, Citation1994; Fernandes et al., Citation1991; Wu & Haard, Citation2000; Zhao et al., Citation1996) were also reported to be non-competitive inhibitors against papain. Ki of almond cystatin binding with papain was comparative with other phytocystatin from corn cystatin I and II (37 and 6.5 nM, respectively) (Abe et al., Citation1994), tomato cystatin (4.7 nM) (Jacinto et al., Citation1998), cowpea cystatin (6.1 nM) (Fernandes et al., Citation1991), two soybean cystatins induced by methyl jasmonate and wounding (57 and 21 nM) (Zhao et al., Citation1996), and oryzacystatin I and II (30 and 830 nM, respectively) (Kondo et al., Citation1990). Ki value was also comparable with some animal cystatin, e.g. goat pancreas (Priyadarshini & Bano, Citation2010), goat brain (Sumbul & Bano, Citation2006). Association rate constants (k+1) was calculated for almond cystatin assuming that papain and the cystatin react in such a way that dissociation rate is low enough to neglect the reverse reaction during the initial part of the reaction. The k+1 values were calculated 3.74 × 103 ± 05 for papain. The reaction was shifted towards first-order kinetics by adding excess substrate for the determination of dissociation rate constant (k–1). It was found to be 1.70 × 10−4 ± 04 for papain. k+1 and k−1 for ficin and bromelain are given in Table .
Figure 6. Determination of inhibition constant Ki with papain. Papain was used at a final concentration of 0.06 nM with increasing amount of inhibitor (0.06–0.30 nM) and measuring the residual activity using casein as substrate.
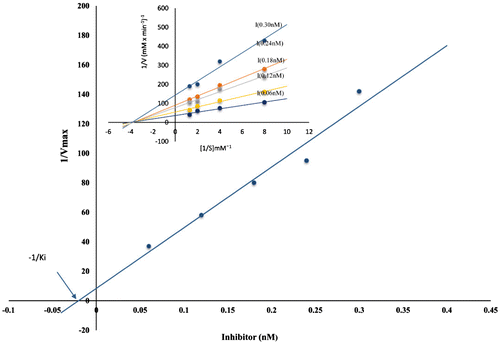
Table 2. Kinetic constants obtained on interaction of almond cystatin with proteases—papain, ficin, and bromelain
Lineweaver–Burk plot shows that the purified cystatin is a non-competitive inhibitor of papain. Almond cystatin is found to be effective inhibitor of papain as indicated by their low Ki value. This finding is supported by the results reported earlier (Abe et al., Citation1994; Fernandes et al., Citation1991; Kondo et al., Citation1990; Sadaf et al., Citation2005). The constant value of Ki with increasing substrate concentration suggests that the inhibition is non-competitive. Tight binding inhibitors have high association rate constants and low dissociation rate constant. The inhibitors with low Ki and k–1, while high k+1 suggest that enzyme–inhibitor complex is stable and complex formation is very fast. Hence, our results give comprehensive information that the cystatin is a potent inhibitors of papain.
The half-life values of almond cystatin calculated were found to be 4.07 × 103 s, 1.11 × 103 s, and 9.76 × 102 s for papain, ficin, and bromelain, respectively. It indicates that the papain–cystatin complex is more stable than the either of the ficin or bromelain–cystatin complexes. Similar trends have been seen for the inhibitors purified from the brain of Capra hircus and goat pancreas (Khaki et al., Citation2016; Priyadarshini & Bano, Citation2010).
3.8. Spectroscopic analysis
Interaction of cystatin with papain was studied using UV–vis spectroscopy. As seen in Figure (A), almond cystatin gave typical protein absorption with maxima at 280 nm. On interaction with papain, spectra indicate that the environment of several aromatic amino acid residues in the protein have been perturbed. These changes are indicative of alterations in conformation of either one or both the proteins. Such changes have also been reported earlier (Amin et al., Citation2011).
Figure 7. (A) Ultraviolet absorption spectra for almond cystatin bound to papain. For complex formation with papain, almond cystatin concentration used was 40 μg/1.5 ml. (B) Fluorescence spectra of cystatin in complex with papain. A fluorescence spectrum of almond cystatin, papain, and papain–cystatin complex was measured at excitation wavelength 280 nm and emission wavelength 300–400 nm. The concentration of almond cystatin was 40 μg/1.5 ml. (C) Far-UV CD spectrum of almond cystatin 20 μm was recorded between 200 and 250 nm using 10-mm path length. (D) FTIR spectra of cystatin alone (2 mM) prepared in sodium phosphate buffer (50 mM, pH 7.5), and cystatin with papain (0.5 mM) was prepared in sodium phosphate buffer (50 mM, pH 7.5). Average of five scan was taken. The spectrum of bound form cystatin was obtained by subtraction of the spectrum of papain from the cystatin + papain complex.
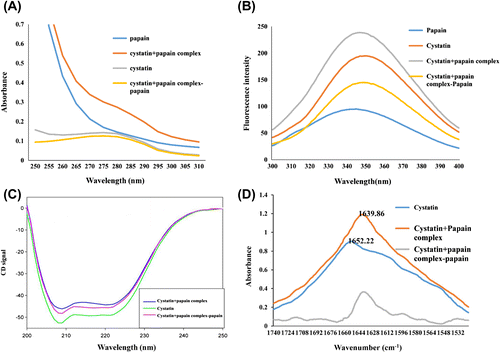
The fluorescence emission spectra of the complex of purified almond cystatin with papain resulted in conformational changes of the proteins (Figure (B)). Fluorescence emission spectra showed maxima at 345 nm for cystatin. Complex formation leads to increase in intensity and a blue shift of 5 nm. This suggests that the interaction of the two proteins occurs in a manner leading to the quenching of absorbing groups to the polar environment due to conformational change or local interactions affecting chromophoric groups of two proteins in the complex (Friefelder, Citation1982).
In Figure (A) and (B), final spectra (cystatin + papain complex − papain) obtained after subtracting the spectra of papain from cystatin–papain complex shows the overall change in cystatin after complex formation.
3.9. CD spectroscopy
The possible overall structure of cystatin–papain complex was monitored by CD spectroscopy, and the results are presented in Figure (C). The far-UV CD spectra of cystatin in the absence of papain showed two negative bands at 208 and 222 nm, a characteristic of the typical α-helix protein. After complex formation, loss in the secondary structure was observed. The α-helical content was calculated from the ellipticity values at 222 nm using equation given by Chen et al. (Citation1972) The α-helical content of native cystatin was found to be 36.8%, while cystatin–papain complex has helical content 32.7%. There is a decrease of 4.1% in helicity values for cystatin–papain complex. This suggests that cystatin alone has greater secondary structural integrity (α-helix) and stability as compared to cystatin–papain complex. This finding is comparative to other previously reported animal cystatins (caprine brain cystatin and goat lung cystatin) (Khaki et al., Citation2016; Khan & Bano, Citation2009). In Figure (C), final CD spectra of cystatin + papain complex − papain obtained after subtracting the spectra of papain from cystatin–papain complex shows the overall ellipticity change in cystatin after complex formation.
3.10. Fourier transform infrared spectroscopy
FTIR spectroscopy provides information about the secondary structure content of almond cystatin. In the IR spectra of proteins, the secondary structure is most clearly reflected by the amide I and amide II bands. The absorption associated with the Amide I band leads to stretching vibrations of the C=O (80%) bond of the amide, absorption associated with the Amide II band leads primarily to bending vibrations of the N–H (60%) bond (Krimm & Bandekar, Citation1986; Pelton & McLean, Citation2000). Because both the C=O and the N–H bonds are involved in the hydrogen bonding that takes place between the different elements of secondary structure, the locations of both the Amide I and Amide II bands are sensitive to the secondary structure content of a protein (Byler & Susi, Citation1986; Surewicz & Mantsch, Citation1988). The amide I band absorbs near 1,650 cm−1 (mainly C=O stretch) and the amide II band absorbs near 1,550 cm−1 (C–N stretching coupled with N–H bending modes). It has also been reported that, for a native protein, the amide I component for the alpha helical structure locates at 1,656 ± 2 cm−1, the band components for beta sheet structure should locate between 1,622 and 1,642 cm−1 (lower wave number beta sheet bands) and between 1,690 and 1,698 cm−1 (higher wave number beta sheet bands) (Dong et al., Citation1997).
Data of cystatin alone and cystatin–papain complex show significant shift in the peak intensity from 1652.22 to 1639.86 cm−1 (Figure (D)) and significant changes in the structure of cystatin from that of α helix to β structure (lower wavenumber β-sheet bands), after co-incubating cystatin with papain for 30 min. This result is consistent with the result of UV, fluorescence, and CD spectroscopy. The result was very comparative with cystatin purified from Phaseolus mungo (Urd) (Sharma, Rashid, & Bano, Citation2005). In Figure (D), final absorbance spectra (cystatin + papain complex − papain) obtained after subtracting the spectra of papain from cystatin–papain complex shows the overall change in the conformation of cystatin after complex formation.
3.11. Isothermal titration calorimetry
ITC experiment was employed to measure the binding thermodynamics and stoichiometry between cystatin and papain. The isothermogram for this titration is shown in Figure , and the derived parameters are shown in Table . The best fits for the titration was obtained using a single site of binding model with the lowest χ2-value. Enthalpy change (∆H) of the binding was negative, whereas entropy change (∆S) of the binding was positive which indicated the exothermic and entropically driven nature of binding. The negative value of enthalpy is directly related to weak van der Waals interactions during complex formation (Færgeman, Sigurskjold, Kragelund, Andersen, & Knudsen, Citation1996). The binding stoichiometry of cystatin and papain is 0.870 ± 0.03 sites which indicate that cystatin is surrounded by nearly one papain molecule. The results of ITC were in agreement with that of kinetic parameters.
Figure 8. Isothermal titration calorimetry of almond cystatin titrated with papain.
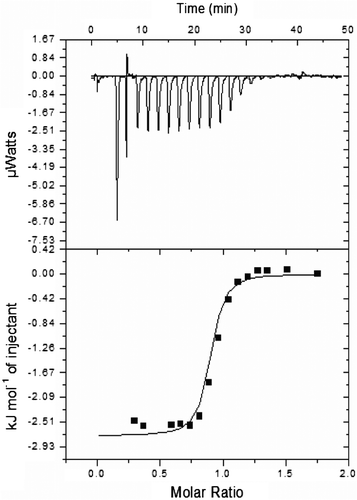
Table 3. Binding and thermodynamic parameters of cystatin with papain
4. Conclusion
Protease inhibitors are of great importance in medicine as proteolysis has significant role in almost every biological process. The potential for the natural inhibitors in agriculture is enormous, awaiting full-scale exploration. Recombinant PIs have proved to be of particular interest as promising models for studying the ecological impacts of insect-resistant transgenic plants, co-evolutionary processes in plant–insect systems, and recombinant protein-mediated pleiotropic effects in transgenic plants. The highly specific nature of the inhibitor–protease interaction makes it a valuable tool in medicine, agriculture, and food technology. An active area of research in the development of transgenic plant is the one which express protein inhibitors with improved features. With such a vast potential and application there is an emergent need for identification of a high yielding protein-rich source. Almond was found to be a high yielding cystatin-rich source. A better knowledge of cysteine protease-mediated processes in plant system with a thorough perception of cysteine protease regulation in plant cells and the design of potent cystatin inhibitory variants, should pave the way, to metabolic pathways involving recombinant cystatins in forthcoming years. Chances for further exploration of improved tolerance in cystatin-expressing plants to different abiotic stress are likely to happen. Adding to the pool of already purified cystatins, almond cystatin shows a good association with other purified plant cystatins in terms of its physicochemical properties. Data shed some light on the mechanism of interaction of almond cystatin with papain. Considering all the data obtained for the purified almond cystatin it can be placed in the third group of plant cystatins (Wang et al., Citation2008) with respect to its high molecular weight (Jacinto et al., Citation1998; Wu & Haard, Citation2000). Plants are immobile organisms and have a variety of parasite-defense systems (Habib & Fazili, Citation2007; Lawrence & Koundal, Citation2002), and it is likely that the seed cystatins are involved in such systems. Further research into such regulatory and defensive mechanisms will facilitate the development of transgenic crops.
Acknowledgment
Facilities provided by Aligarh Muslim University are gratefully acknowledged. We are grateful to SAP-DRS-III and UGC-BSR program for their generous research support.
Additional information
Funding
Notes on contributors
Azad Alam Siddiqui
Bilqees Bano is an active member of the Department of Biochemistry, Faculty of Life Sciences Aligarh Muslim University, Aligarh. She has received several fellowships (at pre-PhD level from CSIR-JRF, SRF; at post-PhD level from CSIR-PDF Award and Pool Officer Fellowship; Commonwealth Academic Staff Fellowship—UK; Indo-US Fulbright fellowship—USA; NIH Fellowship Award—USA). She has 34 years’ research experience in the fields of proteins, enzymes, and clinical biochemistry. Bano’s current interest is in cysteine protease inhibitors with particular reference to their amyloid formation and evaluation of potential candidates for their anti-amyloidogenic properties.
References
- Abe, M., Abe, K., Iwabuchi, K., Domoto, C., & Arai, S. (1994). Corn cystatin I expressed in Escherichia coli: Investigation of its inhibitory profile and occurrence in corn kernels. Journal of Biochemistry, 116, 488–492.
- Abe, M., Abe, K., Kuroda, M., & Arai, S. (1992). Corn Kernel cysteine proteinase inhibitor as a novel cystatin superfamily member of plant origin. Molecular cloning and expression studies. European Journal of Biochemistry, 209, 933–937. 10.1111/ejb.1992.209.issue-3
- Abe, K., Kondo, H., Watanabe, H., Emori, Y., & Arai, S. (1991). Oryzacystatins as the first well-defined cystatins of plant origin and their target proteinases in rice seeds. Biomedica Biochimica Acta, 50, 637–641.
- Abrahamson, M., Barrett, A. J., Salvesen, G., & Grubb, A. (1986). Isolation of six cysteine proteinase inhibitors from human urine. Their physicochemical and enzyme kinetic properties and concentrations in biological fluids. The Journal of Biological Chemistry, 261, 11282–11289.
- Amin, F., Ali Khan, A., Jahan Rizvi, S., & Bano, B. (2011). Purification and characterizaion of buffalo brain cystatin. Protein and Peptide Letters, 18, 210–218.10.2174/092986611794475011
- Andrews, P. (1964). Estimation of the molecular weights of proteins by Sephadex gel-filtration. Biochemical Journal, 91, 222–233.10.1042/bj0910222
- Barrett, A. J. (1987). The cystatins: A new class of peptidase inhibitors. Trends in Biochemical Sciences, 12, 193–196. doi:10.1016/0968-0004(87)90092-2
- Belenghi, B., Acconcia, F., Trovato, M., Perazzolli, M., Bocedi, A., Polticelli, F., … Delledonne, M. (2003). AtCYS1, a cystatin from Arabidopsis thaliana, suppresses hypersensitive cell death. European Journal of Biochemistry, 270, 2593–2604.10.1046/j.1432-1033.2003.03630.x
- Bhat, S. A., Sohail, A., Siddiqui, A. A., & Bano, B. (2014). Effect of non-enzymatic glycation on cystatin: A spectroscopic study. Journal of Fluorescence, 24, 1107–1117. doi:10.1007/s10895-014-1391-2
- Bolter, C. J. (1993). Methyl jasmonate induces papain inhibitor(s) in tomato leaves. Plant Physiology, 103, 1347–1353.10.1104/pp.103.4.1347
- Botella, M. A., Xu, Y., Prabha, T. N., Zhao, Y., Narasimhan, M. L., Wilson, K. A., … Hasegawa, P. M. (1996). Differential expression of soybean cysteine proteinase inhibitor genes during development and in response to wounding and methyl jasmonate. Plant Physiology, 112, 1201–1210.10.1104/pp.112.3.1201
- Byler, D. M., & Susi, H. (1986). Examination of the secondary structure of proteins by deconvolved FTIR spectra. Biopolymers, 25, 469–487. doi:10.1002/bip.360250307
- Chan, S.-N., Abu Bakar, N., Mahmood, M., Ho, C.-L., & Shaharuddin, N. A. (2014). Molecular cloning and characterization of novel phytocystatin gene from turmeric, Curcuma longa. BioMed Research International, 2014, 973790. doi:10.1155/2014/973790
- Chen, Y. H., Yang, J. T., & Martinez, H. M. (1972). Determination of the secondary structures of proteins by circular dichroism and optical rotatory dispersion. Biochemistry, 11, 4120–4131.10.1021/bi00772a015
- Christeller, J. T. (2005). Evolutionary mechanisms acting on proteinase inhibitor variability. FEBS Journal, 272, 5710–5722. doi:10.1111/j.1742-4658.2005.04975.x
- Chu, M.-H., Liu, K.-L., Wu, H.-Y., Yeh, K.-W., & Cheng, Y.-S. (2011). Crystal structure of tarocystatin–papain complex: Implications for the inhibition property of group-2 phytocystatins. Planta, 234, 243–254. doi:10.1007/s00425-011-1398-8
- Davis, P. A., & Iwahashi, C. K. (2001). Whole almonds and almond fractions reduce aberrant crypt foci in a rat model of colon carcinogenesis. Cancer Letters, 165, 27–33.10.1016/S0304-3835(01)00425-6
- Dong, A., Kendrick, B., Kreilgård, L., Matsuura, J., Manning, M. C., & Carpenter, J. F. (1997). Spectroscopic study of secondary structure and thermal denaturation of recombinant human factor XIII in aqueous solution. Archives of Biochemistry and Biophysics, 347, 213–220. doi:10.1006/abbi.1997.0349
- DuBois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 28, 350–356. doi:10.1021/ac60111a017
- Dunaevsky, Y. E., Elpidina, E. N., Vinokurov, K. S., & Belozersky, M. A. (2005). Protease inhibitors in improvement of plant resistance to pathogens and insects. Molecular Biology, 39, 608–613. doi:10.1007/s11008-005-0076-y
- Ellman, G. L. (1959). Tissue sulfhydryl groups. Archives of Biochemistry and Biophysics, 82, 70–77. doi:10.1016/0003-9861(59)90090-6
- Esfahlan, A. J., Jamei, R., & Esfahlan, R. J. (2010). The importance of almond (Prunus amygdalus L.) and its by-products. Food Chemistry, 120, 349–360. doi:10.1016/j.foodchem.2009.09.063
- Færgeman, N. J., Sigurskjold, B. W., Kragelund, B. B., Andersen, K. V., & Knudsen, J. (1996). Thermodynamics of ligand binding to acyl-coenzyme a binding protein studied by titration calorimetry †. Biochemistry, 35, 14118–14126. doi:10.1021/bi960545z
- Fernandes, K. V. S., Campos, F. A. P., Do Val, R. R., & Xavier-Filho, J. (1991). The expression of papain inhibitors during development of cowpea seeds. Plant Science, 74, 179–184. doi:10.1016/0168-9452(91)90044-9
- Fountoulakis, M., & Gasser, R. (2003). Proteomic analysis of the cell envelope fraction of Escherichia coli. Amino Acids, 24, 19–41. doi:10.1007/s00726-002-0339-z
- Friefelder, D. (1982). Physical biochemistry, applications to biochemistry and molecular biology (2nd ed.). New York, NY: WH Freeman and Company.
- Gruden, K., Štrukelj, B., Ravnikar, M., Poljšak-Prijatelj, M., Mavrič, I., Brzin, J., … Kregar, I. (1997). Potato cysteine proteinase inhibitor gene family: Molecular cloning, characterisation and immunocytochemical localisation studies. Plant Molecular Biology, 34, 317–323.10.1023/A:1005853026333
- Habib, H., & Fazili, K. M. (2007). Plant protease inhibitors: A defense strategy in plants. Biotechnology and Molecular Biology Reviews, 2, 068–085.
- Haq, S. K., Atif, S. M., & Khan, R. H. (2004). Protein proteinase inhibitor genes in combat against insects, pests, and pathogens: Natural and engineered phytoprotection. Archives of Biochemistry and Biophysics, 431, 145–159. doi:10.1016/j.abb.2004.07.022
- Hartl, M., Giri, A. P., Kaur, H., & Baldwin, I. T. (2011). The multiple functions of plant serine protease inhibitors. Plant Signaling & Behavior, 6, 1009–1011. doi:10.4161/psb.6.7.15504
- Hong, J. K., Je, J., Song, C., Hwang, J. E., Lee, Y.-H., & Lim, C. O. (2012). Biochemical analysis of a Chinese cabbage phytocystatin-1. Genes & Genomics, 34, 13–18. doi:10.1007/s13258-011-0150-x
- Hyson, D. A., Schneeman, B. O., & Davis, P. A. (2002). Almonds and almond oil have similar effects on plasma lipids and LDL oxidation in healthy men and women. Journal of Nutrition, 132, 703–707.
- Jacinto, T., Fernandes, K. V. S., Machado, O. L. T., & Siqueira-Júnior, C. L. (1998). Leaves of transgenic tomato plants overexpressing prosystemin accumulate high levels of cystatin. Plant Science, 138, 35–42. doi:10.1016/S0168-9452(98)00153-8
- Joanitti, G. A., Freitas, S. M., & Silva, L. P. (2006). Proteinaceous protease inhibitors: Structural features and multiple functional faces. Current Enzyme Inhibition, 2, 199–217, doi:10.2174/157340806777934801
- Khaki, P. S. S., Feroz, A., Amin, F., Rehman, M. T., Bhat, W. F., & Bano, B. (2016). Structural and functional studies on a variant of cystatin purified from brain of Capra hircus. Journal of Biomolecular Structure and Dynamics, 1–17, doi:10.1080/07391102.2016.1191375
- Khan, M. S., & Bano, B. (2009). Purification, characterization and kinetics of thiol protease inhibitor from goat (Capra hircus) lung. Biochemistry (Moscow), 74, 781–788. 10.1134/S0006297909070116
- Kondo, H., Abe, K., Nishimura, I., Watanabe, H., Emori, Y., & Arai, S. (1990). Two distinct cystatin species in rice seeds with different specificities against cysteine proteinases. Molecular cloning, expression, and biochemical studies on oryzacystatin-II. The Journal of Biological Chemistry, 265, 15832–15837.
- Kouzuma, Y., Kawano, K., Kimura, M., Yamasaki, N., Kadowaki, T., & Yamamoto, K. (1996). Purification, characterization, and sequencing of two cysteine proteinase inhibitors, sea and scb, from sunflower (Helianthus annuus) seeds. Journal of Biochemistry, 119, 1106–1113.10.1093/oxfordjournals.jbchem.a021355
- Krimm, S., & Bandekar, J. (1986). Vibrational spectroscopy and conformation of peptides, polypeptides, and proteins. Advances in Protein Chemistry, 38, 181–364.10.1016/S0065-3233(08)60528-8
- Kunitz, M. (1947). Crystalline soybean trypsin inhibitor: II. general properties. The Journal of General Physiology, 30, 291–310.10.1085/jgp.30.4.291
- Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227, 680–685. doi:10.1038/227680a0
- Lawrence, P. K., & Koundal, K. R. (2002). Plant protease inhibitors in control of phytophagous insects. Electronic Journal of Biotechnology, 5, 5–6. doi:10.2225/vol5-issue1-fulltext-3
- Lehninger, A. L., Nelson, D. L., Farr, A. L., & Cox, M. M. (2000). Principles of biochemistry (2nd ed., pp. 558–560). New York, NY: Worth Publishers.
- Lin, Y.-H., Li, H.-T., Huang, Y.-C., Hsieh, Y.-C., Guan, H.-H., Liu, M.-Y., … Chen, C.-J. (2006). Purification, crystallization and preliminary X-ray crystallographic analysis of rice Bowman-Birk inhibitor from Oryza sativa. Acta Crystallographica. Section F, Structural Biology and Crystallization Communications, 62, 522–524. http://doi.org/10.1107/S1744309106014795
- Lowry, O. H., Rosebrough, N. J., Farr, A. L., & Randall, R. J. (1951). Protein measurement with the Folin phenol reagent. The Journal of Biological Chemistry, 193, 265–275.
- Margis, R., Reis, E. M., & Villeret, V. (1998). Structural and phylogenetic relationships among plant and animal cystatins. Archives of Biochemistry and Biophysics, 359, 24–30. doi:10.1006/abbi.1998.0875
- Margis-Pinheiro, M., Zolet, A. C. T., Loss, G., Pasquali, G., & Margis, R. (2008). Molecular evolution and diversification of plant cysteine proteinase inhibitors: New insights after the poplar genome. Molecular Phylogenetics and Evolution, 49, 349–355. doi:10.1016/j.ympev.2008.04.025
- Martinez, M., Abraham, Z., Gambardella, M., Echaide, M., Carbonero, P., & Diaz, I. (2005). The strawberry gene Cyf1 encodes a phytocystatin with antifungal properties. Journal of Experimental Botany, 56, 1821–1829. doi:10.1093/jxb/eri172
- Mosolov, V. V., & Valueva, T. A. (2008). Proteinase inhibitors in plant biotechnology: A review. Applied Biochemistry and Microbiology, 44, 233–240. doi:10.1134/S0003683808030010
- Neurath, H. (1989). Proteolytic processing and physiological regulation. Trends in Biochemical Sciences, 14, 268–271.10.1016/0968-0004(89)90061-3
- Nissen, M. S., Kumar, G. N. M., Youn, B., Knowles, D. B., Lam, K. S., Ballinger, W. J., … Kang, C. (2009). Characterization of solanum tuberosum multicystatin and its structural comparison with other cystatins. The Plant Cell Online, 21, 861–875. doi:10.1105/tpc.108.064717
- Pelton, J. T., & McLean, L. R. (2000). Spectroscopic methods for analysis of protein secondary structure. Analytical Biochemistry, 277, 167–176. doi:10.1006/abio.1999.4320
- Pernas, M., Sánchez-Monge, R., Gómez, L., & Salcedo, G. (1998). A chestnut seed cystatin differentially effective against cysteine proteinases from closely related pests. Plant Molecular Biology, 38, 1235–1242.10.1023/A:1006154829118
- Priyadarshini, M., & Bano, B. (2010). Cystatin like thiol proteinase inhibitor from pancreas of Capra hircus: Purification and detailed biochemical characterization. Amino Acids, 38, 1001–1010. doi:10.1007/s00726-009-0308-x
- Rashid, F., Sharma, S., & Bano, B. (2005). Detailed biochemical characterization of human placental cystatin (HPC). Placenta, 27, 822–831.
- Rashid, F., Sharma, S., & Bano, B. (2006). Detailed biochemical characterization of human placental cystatin (HPC). Placenta, 27, 822–831. doi:10.1016/j.placenta.2005.09.005
- Rassam, M., & Laing, W. A. (2004). Purification and characterization of phytocystatins from kiwifruit cortex and seeds. Phytochemistry, 65, 19–30.10.1016/j.phytochem.2003.09.019
- Rawlings, N. D., Tolle, D. P., & Barrett, A. J. (2004). MEROPS: The peptidase database. Nucleic Acids Research, 32, 160D–164. http://doi.org/10.1016/j.cbpb.2005.08.00710.1093/nar/gkh071
- Sadaf, Z., Shahid, P. B., & Bilqees, B. (2005). Isolation, characterization and kinetics of goat cystatins. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 142, 361–368. http://doi.org/10.1016/j.cbpb.2005.08.007
- Sathe, S. K., Wolf, W. J., Roux, K. H., Teuber, S. S., Venkatachalam, M., & Sze-Tao, K. W. C. (2002). Biochemical characterization of amandin, the major storage protein in almond ( Prunus dulcis L.). Journal of Agricultural and Food Chemistry, 50, 4333–4341.10.1021/jf020007v
- Sharma, S., Rashid, F., & Bano, B. (2005). Biochemical and biophysical changes induced by fungicide sodium diethyl dithiocarbamate (SDD), in phytocystatin purified from phaseolus mungo (Urd): A commonly used indian legume. Journal of Agricultural and Food Chemistry, 53, 6027–6034. doi:10.1021/jf050580p
- Sharma, S., Rashid, F., & Bano, B. (2006). Unfolding during urea denaturation of a low molecular weight phytocystatin (thiol protease inhibitor) purified from phaseolus mungo (Urd). Protein and Peptide Letters, 13, 323–329.10.2174/092986606775974447
- Shyu, D. J. H., Chou, W.-M., Yiu, T.-J., Lin, C. P. C., & Tzen, J. T. C. (2004). Cloning, functional expression, and characterization of cystatin in sesame seed. Journal of Agricultural and Food Chemistry, 52, 1350–1356. doi:10.1021/jf034989v
- Siddiqui, A. A., Sohail, A., Bhat, S. A., Rehman, M. T., & Bano, B. (2015). Non-enzymatic glycation of almond cystatin leads to conformational changes and altered activity. Protein and Peptide Letters, 22, 449–459.10.2174/0929866522666150326105704
- Sohail, A., Bhat, S. A., Siddiqui, A. A., Zaman, M., Khan, R. H., & Bano, B. (2015). Conformational transitions induced by in vitro macromolecular crowding lead to the amyloidogenesis of buffalo heart cystatin. Journal of Molecular Recognition, 28, 699–709. doi:10.1002/jmr.2484
- Sumbul, S., & Bano, B. (2006). Purification and characterization of high molecular mass and low molecular mass cystatin from goat brain. Neurochemical Research, 31, 1327–1336. doi:10.1007/s11064-006-9175-y
- Supuran, C. T., Scozzafava, A., & Clare, B. W. (2002). Bacterial protease inhibitors. Medicinal Research Reviews, 22, 329–372. doi:10.1002/med.10007
- Surewicz, W. K., & Mantsch, H. H. (1988). New insight into protein secondary structure from resolution-enhanced infrared spectra. Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology, 952, 115–130.10.1016/0167-4838(88)90107-0
- Synnes, M. (1998). Purification and characterization of two cysteine proteinase inhibitors from the skin of Atlantic salmon (Salmo Salar L.). Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 121, 257–264. doi:10.1016/S0305-0491(98)10098-6
- Valueva, T. A., & Mosolov, V. V. (2004). Role of inhibitors of proteolytic enzymes in plant defense against phytopathogenic microorganisms. Biochemistry (Moscow), 69, 1305–1309. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/1562738410.1007/s10541-005-0015-5
- Waldron, C., Wegrich, L. M., Ann Owens Merlo, P., & Walsh, T. A. (1993). Characterization of a genomic sequence coding for potato multicystatin, an eight-domain cysteine proteinase inhibitor. Plant Molecular Biology, 23, 801–812.10.1007/BF00021535
- Wang, K.-M., Kumar, S., Cheng, Y.-S., Venkatagiri, S., Yang, A.-H., & Yeh, K.-W. (2008). Characterization of inhibitory mechanism and antifungal activity between group-1 and group-2 phytocystatins from taro (Colocasia esculenta). FEBS Journal, 275, 4980–4989. doi:10.1111/j.1742-4658.2008.06631.x
- Weber, K., & Osborn, M. (1969). The reliability of molecular weight determinations by dodecyl sulfate-polyacrylamide gel electrophoresis. The Journal of Biological Chemistry, 244, 4406–4412.
- Wu, J., & Haard, N. F. (2000). Purification and characterization of a cystatin from the leaves of methyl jasmonate treated tomato plants. Comparative Biochemistry and Physiology. Toxicology & Pharmacology, 127, 209–220.
- Yang, A. H., & Yeh, K. W. (2005). Molecular cloning, recombinant gene expression, and antifungal activity of cystatin from taro (Colocasia esculenta cv. Kaosiung no. 1). Planta, 221, 493–501. doi:10.1007/s00425-004-1462-8
- Zhao, Y., Botella, M. A., Subramanian, L., Niu, X., Nielsen, S. S., Bressan, R. A., & Hasegawa, P. M. (1996). Two wound-inducible soybean cysteine proteinase inhibitors have greater insect digestive proteinase inhibitory activities than a constitutive homolog. Plant Physiology, 111, 1299–1306.10.1104/pp.111.4.1299