
Abstract
The Mesozoic chondrichthyan fossil record from northern South America is scarce and poorly known containing only a few reports of lamniform and ptychodont species from the Upper Cretaceous of Colombia and Venezuela. Here, we report the discovery of two associated (found together) dorsal fin spines of a hybodont shark from the Valanginian deposits of the Rosablanca Formation (Andes of Colombia). This hybodont along with the presence of ammonites and other mollusks, turtles, plesiosaurs and ichthyosaurs support a marine depositional environment for this geological unit. Although both spines described here are fragmentary and without diagnostic characters to allow identification to generic level, their finding represents the first record of this shark group from northernmost part of South America and northwestern margin of Gondwana. This new record expands the known distribution of this group of sharks during Lower Cretaceous.
Introduction
Hybodontiform sharks belong to an extinct chondrichthyan lineage with a fossil record that spanned from the Paleozoic to end of the Mesozoic.[Citation1] Their remains are frequently found in Mesozoic rocks around the world [Citation1,2] especially in Triassic and Jurassic rocks where hybodonts are the dominant selachian group in both marine and non-marine environments.[Citation3] This dominant clade became restricted at the end of the Jurassic when neoselachians started displacing hybodonts.[Citation3] However, after the neoselachians radiation, hybodonts survived in restricted marine and fluvial ecosystem until the end of the Cretaceous, when the group became extinct.[Citation1] The Mesozoic hybodont paleodiversity has been estimated by Cappetta [Citation1] to represent at least seven families and 27 genera. However, there are still gaps in our knowledge of hybodonts, whose interrelationships are mostly reconstructed on the basis of isolated teeth [Citation2] and few articulated specimens.[Citation4]
The fossil record of Mesozoic chondrichthyans from northern South America is poorly known with only few remains of lamniforms and ptychodonts from the Upper Cretaceous of Colombia and Venezuela.[Citation5–11] The Mesozoic fossil record of hybodonts in South America includes the following occurrences: several articulated and semi-articulated specimens from the Lower Cretaceous of Brazil (e.g. Tribodus limae Brito and Ferreira 1989 [Citation12]), isolated teeth, cephalic and dorsal fin spines, and dermal denticles from different strata of the Triassic and Jurassic of Argentina [Citation13,14]; Upper Jurassic-? Lower Cretaceous of Uruguay [Citation15] and Upper Jurassic and Lower Cretaceous of Brazil.[Citation16–21] Here, we describe two dorsal fin spines of a hybodontiform from the Lower Cretaceous of Colombia (Rosablanca Formation), representing the first record of this shark group from the northernmost part of South America and shedding additional lights to the chondrichthyan paleodiversity of the northwestern margin of Gondwana during Lower Cretaceous time.
Materials and methods
The dorsal fin spines described here were found in a limestone layer of the Lower Cretaceous Rosablanca Formation (Figure ) (El Alto locality: 6° 49′ 30.21″ N, and 73° 15′ 9.43″ W), a geological unit widely exposed in Zapatoca Town area (Santander Department), northeastern Cordillera of Colombia (Figure ). The Rosablanca Formation has been interpreted as shallow marine deposits [Citation22] of late Valanginian age, based on ammonites.[Citation23,24] The presence of others fossil vertebrate such as turtles, plesiosaurs, ichthyosaurs, bony fish teeth, invertebrates such as ammonites, bivalves, oysters and gastropods support that the deposition of the Rosablanca Formation occurred under marine conditions.[Citation23,24] The specimens referred here (IGMp-880644; IGMp-880645) are housed in the paleontological collection of the Museo Geológico Nacional “José Royo y Gómez” of the Colombian Geological Survey (Servicio Geológico Colombiano, IGMp), in Bogotá, Colombia. The systematic classification follows Cappetta [Citation1] and the hybodont fin spine terminology follows Maisey [Citation25] and Rees and Underwood [Citation3]. Taxonomic identification is based on an extensive bibliographical review and anatomical comparison studies with fossil specimens from the following collections: Natural History Museum of Basel, Switzerland; Palaeontological Institute and Museum at the University of Zurich Switzerland; René Kindlimann (RK) private collection with public access, Uster, Switzerland.
Figure 1. Geographical location map (A), and stratigraphic column of the Rosablanca Formation (B). South American map redrawn from Google Maps, Colombia map modified from [24], art illustration of the shark by Jorge Blanco.
![Figure 1. Geographical location map (A), and stratigraphic column of the Rosablanca Formation (B). South American map redrawn from Google Maps, Colombia map modified from [24], art illustration of the shark by Jorge Blanco.](/cms/asset/92dbbaf0-175d-4129-9d84-ee702b346e0b/tneo_a_1191749_f0001_oc.gif)
Systematic paleontology
Class Chondrichthyes Huxley, 1880 [Citation26]
Subclass Elasmobranchii Bonaparte, 1838 [Citation27]
Order Hybodontiformes Maisey, 1975 [Citation28]
Family Hybodontidae Owen, 1846 [Citation29]
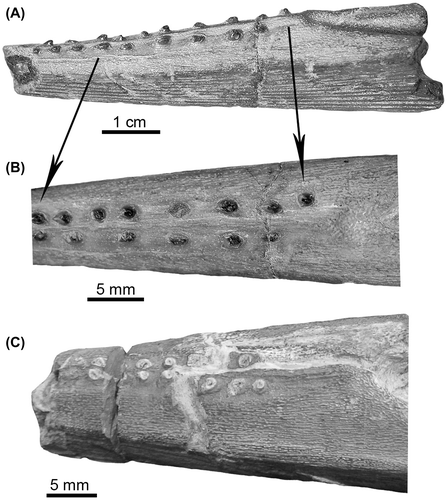
Gen. et sp. indet. (Figure (A)–(I))
Figure 2. Hybodont fin spines from the Rosablanca Formation. (A–F) Specimen IGMp-880644; (G–I) Specimen IGMp-880645.

Referred material. Two incomplete and fragmented dorsal spines of indeterminate position. Both specimens were found in close proximity in the same horizon (less than 2 m of separation between each other), suggesting that both could belong to the same individual; however, the alternative hypothesis of different individuals is also plausible.
Description. The most complete spine (IGMp-880644) has a robust shape with a length of 129 mm (Figure (A)–(F)). In cross section, the spine is approximately oval (Figure (E)), while the tip part of the right side below the basal opening and the unornamented base of this specimen are missing (Figure (A)–(D)); the anterior side of the spine is slightly straight. On the posterior side, there are two parallel rows of small hooks, without an evident groove between them (Figure (D)–(F)). The basal opening is long and wide, having an oval shape in its upper section. Complete longitudinal ribs are not well preserved due to potential erosional effects; however, some remains of these can be observed on the anterior part of both lateral surfaces (Figure (B) and (C)). Specimen IGMp-880645 corresponds to a small spine fragment that includes the upper section of the basal opening (Figure (H) and (I)) and an eroded and broken long fragment (~120 mm) contained whitin in a calcareous matrix (Figure (G)). The inadequate preservation of those fragments does not permit the observation of longitudinal ribs or rows of small hooks. The insertion area of the spines cannot be observed in any of the two specimens describe here (IGMp-880644; IGMp-880645). For this reason, their assignation to an anterior or posterior position is not possible.
Discussion and conclusion
Hybodont fin spines are unique in several aspects, allowing us to distinguish them from fin spines of other xenacanthoid, ctenacanthoid, holocephalan, among others chondrichthyans.[Citation25,28,30] Some of the most important hybodont fin spine characters can be found in Maisey [Citation25,28] and Cione et al. [Citation13], including (a) spines are elongated and gently curved posteriorly showing a typical oval shape in cross section; (b) posterior face strongly convex with denticles always present and arranged in a pair of longitudinal rows; (c) spine deeply penetrating myotome with a wide and long basal opening representing approximately 60% of the spine total length; and (d) several longitudinal ridges on the anterior and lateral faces, but not present in Asteracanthus Agassiz, 1837.[Citation31] Specimen IGMp-880644 although incomplete could represent at least 50% of the entire spine (Figure (A)). Its wide and long basal opening, the two rows with hooks on the posterior side and the presence of longitudinal ribs (Figure (A)–(F)), allow us to refer this to a hybodont shark. Although the other two eroded and broken spine fragments assigned to IGMp-880645 preserve no diagnostic characters, they were found associated with the specimen IGMp-880644, likely belonging both to the same individual. Usually the identification of hybodont genera based on isolated fin spines could be difficult, especially if these are not in association with teeth. Also, as mentioned Rees [Citation2], diagnosis of some Mesozoic hybodonts genera, on the basis of isolated teeth, could sometimes be difficult and entire dentitions are often needed.
Among hybodonts from the Cretaceous of South America, only Tribodus limae is represented by articulated and semi-articulated specimens with fins specimens and teeth in association,[Citation12,32] while its fine spines do not reach more than 125 mm in total length.[Citation12,13] The size of the latter does not coincide with the large size of the specimens from Colombia. Other species as Planohybodus marki Pinheiro et al. [Citation21], Planohybodus sp. and Parvodus sp., also reported from the Early Cretaceous of Brazil, are solely known by isolated teeth.[Citation20,21]
One of the best preserved characters of the Colombian specimen IGMp-880644 is the two parallel rows of small hooks (without an evident groove between them) on the posterior side. The two hook rows of the Colombian specimens appear quite similar to those present on a fin spine of Planohybodus peterboroughensis Rees and Underwood [Citation3], from the Middle Jurassic of England and deposited at the René Kindlimann private collection (RK/5-255) (Figure (A) and (B)). The fin spines of the P. peterboroughensis (holotype P. 8386) illustrated by Rees and Underwood [Citation3] exhibit two alternating rows of small hooks on their posterior side; however, they are missing the basal-most hook rows. Two parallel hook rows with also are present in the indeterminate hybodont fin spine illustrated by Cione et al. [Citation13] from the Triassic–Jurassic of Argentina. This character has not been observed in fin spines of other hybodont genera that we have used for comparison. However, as has been suggested by Maisey [Citation25], there is a considerable variation in the hook row distribution on the fin spines of hybodont sharks, whereas current knowledge about hybodont fin spine morphology and its importance in taxonomy remains scarce.
Figure 3. Parallel rows of hooks in Hybodont fin spines Middle Jurassic of England (A–B) and Lower Cretaceous of Colombia (C).
Due to the fragmentary condition and the absence of clearly diagnostic characters from the specimens described here from Rosablanca Formation, we refrain from any taxonomic identification to the generic level or below. We expect that future field expeditions and findings will offer more precise systematic information. Nevertheless, the presence of these hybodont remains in the Valanginian of Colombia expands the known, Lower Cretaceous, distribution of this shark group to cover the northwestern margin of Gondwana.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
Funded by the cooperation agreement number 0010 between the Colombian Geological Survey, ISAGEN, and the Smithsonian Tropical Research Institute. Additional funding was supplied by an NSERC Discovery Grant and Canada Research Chair to HCEL and a Quebec Centre for Biodiversity Science grant to TYD.
Acknowledgments
We thank C. Padilla and M. Parra, for the hospitality and opportunity to prepare the fossil material and the Centro de Investigaciones Paleontológicas, Villa de Leyva, Colombia and J. Arenas for registration of specimens at the Museo Geológico ‘José Royo y Gómez’. Thanks to Dr. C. Jaramillo and the team of paleontologists at the Paleosogamoso project, as well as to A. Bustamente and C. Rosero for facilitate contacts and logistics during the fieldwork. Specials thanks to R. Rueda for her hospitality during our fieldwork in Zapatoca. The authors wish to thank René Kindlimann for their generous and important counseling and collaboration. Special thanks to Thodoris Argyriou for his invaluable suggestions and comments. We greatly appreciate comments and suggestions from three anonymous reviewers.
Associate Editor: Miguel Pinto.
References
- Chondrichthyes H. Cappetta: Mesozoic and Cenozoic Elasmobranchii: teeth. Munich: Verlag Dr. Friedrich Pfeil; 2012.
- Rees J. Interrelationships of Mesozoic hybodont sharks as indicated by dental morphology-preliminary results. Acta Geol. Pol. 2008;58:217–221.
- Rees J, Underwood CJ. Hybodont sharks of the English Bathonian and Callovian (middle Jurassic). Palaeontology. 2008;51:117–147.10.1111/pala.2008.51.issue-1
- Maisey JG. Hamiltonichthys mapesi, g. & sp. nov. (Chondrichthyes; Elasmobranchii), from the Upper Pennsylvanian of Kansas. Am. Mus. Novit. 1989;2931:1–42.
- Reinhart RH. A new shark of the family ptychodontide from South America. Univ. Calif. Publ. Geol. Sci. 1951;28:195–202.
- Moody JM, Maisey JG. New Cretaceous marine vertebrate assemblages from North–Western Venezuela and their significance. J. Vertebr. Paleontol. 1994;14:1–8.10.1080/02724634.1994.10011534
- Brito PMM, Janvier P. A Ptychodontid (Chondrichthyes, Elasmobranchii) from the Upper Cretaceous of South America. Geodiversitas. 2002;24:785–790.
- Carrillo-Briceño JD. Presencia del genero Ptychodus (Elasmobranchii: Ptychodontiade) en el Cretácico superior de los Andes de Trujillo Venezuela [Ocurrence of Ptychodus genus (Elasmobranchii:Ptychodontidae) from the upper Cretaceous of Trujillo, Andes, Venezuela]. Geominas. 2009;37:207–210.
- Carrillo-Briceño JD. Presencia de Ptychodus mortoni (Elasmobranchii: Ptychodontidae) en el Cretácico Superior de Venezuela [Ocurrence of Ptychodus mortoni (Elasmobranchii: Ptychodontidae) from the upper Cretaceous of Venezuela]. Rev. Geog. Am. Cent. 2012;46:145–150.
- Carrillo-Briceño JD, Spencer LG. The first tooth set of Ptychodus atcoensis (Elasmobranchii: Ptychodontidae), from the Cretaceous of Venezuela. Swiss J. Palaeontol. 2013;132:69–75.10.1007/s13358-013-0053-3
- Carrillo-Briceño JD, Ayala R, Chávez-Aponte EO, et al. Registro de Serratolamna serrata (Elasmobranchii: Serratolamnidae) en el Cretácico superior (Maestrichtiense) de los Andes Venezolanos [Record of Serratolamna serrata (Elasmobranchii: Serratolamnidae) from the upper Cretaceous (Maastrichtian) of Venezuelan Andes]. Geominas. 2008;36:160–163.
- Brito PMM, Ferreira PLN. The first Hybodont shark, Tribodus limae n. gen., n. sp., from the Lower Cretaceous of Chapada do Araripe (North-East Brazil). An. Acad. Braz. Ciênc. 1989;61:53–57.
- Cione AL, Acosta Hospitaleche C, Mennucci JA, et al. The first shark from the Triassic-Jurassic of South America. Neu. Jb. Geol. Paläontol. 2002, 1:9–18.
- Johns MJ, Albanesi GL, Voldman GG. Freshwater shark teeth (Family Lonchidiidae) from the middle-upper Triassic (Ladinian-Carnian) Paramillo formation in the Mendoza Precordillera, Argentina. J. Vertebr. Paleontol. 2014;34:512–523.10.1080/02724634.2013.803976
- Soto M, Perea D, Toriño P. New remains of Priohybodus arambourgi (Hybodontiformes: Hybodontidae) from Late Jurassic–?earliest Cretaceous deposits in Uruguay. Cretaceous Res. 2012;35:118–123.10.1016/j.cretres.2011.12.001
- Woodward AS. Notes on some vertebrate fossils from the Province of Bahia, Brazil, collected by Joseph Mawson Esq., F.G.S. Ann. Mag. Nat. Hist. 1888;2:132–136.10.1080/00222938809460891
- Lane JA, Maisey JG. Pectoral anatomy of Tribodus limae (Elasmobranchii: Hybodontiformes) from the Lower Cretaceous of northeastern Brazil. J. Vertebr. Paleontol. 2009;29:25–38.10.1080/02724634.2009.10010359
- Pinheiro FL, Figueiredo AEQ, Fortier DC, et al. Fauna de vertebrados eocretácicos de um afloramento da Bacia de Lima Campos, Ceará, Brasil [Lower Cretaceous vertebrates from the Bacia de Lima Campos, Ceará, Brazil]. Rev. Bras. Paleontolog. 2011;14:189–198.10.4072/rbp
- Silva MC, Carvalho MSS, Barreto AMF, et al. Paleoictiofauna da Formação Aliança (Jurássico Superior), Bacia de Jatobá, Nordeste do Brasil [Fossil fish from the Aliança Formation (upper Jurassic), Bacia de Jatobá, Northwestern of Brazil]. In: Carvalho IS, Srivastava NK, Strohschoen O Jr, Lana CC, editors. Paleontologia: Cenários de Vida. Rio de Janeiro: Editora Interciência; 2011. p. 595–608.
- Cupello CD, Bermúdez-Rochas DD, Martill DM, et al. The Hybodontiformes (Chondrichthyes: Elasmobranchii) from the Missão Velha Formation (?Lower Cretaceous) of the Araripe Basin, North-East Brazil. C. R. Palevol. 2012;11:41–47.10.1016/j.crpv.2011.09.005
- Pinheiro FL, de Figueiredo AEQ, Dentzien-Dias PC, et al. Planohybodus marki sp. nov., a new fresh-water hybodontid shark from the Early Cretaceous of northeastern Brazil. Cretaceous Res. 2013;41:210–216.10.1016/j.cretres.2012.12.005
- Guzman G. Los Grifeidos infracretacicos Aetostreon couloni y Ceratrostreon boussingaulti, de la Formación Rosablanca, como indicadores de oscilaciones marinas [Lower Cretaceous ostreids Aetostreon couloni and Ceratrostreon boussingaulti of the Rosablanca Formation, as indicators of marine fluctuations]. In: Etayo S, editor. Proyecto Cretácico, Publicacion especial del Ingeominas. Bogota: Ingeominas; 1985. p. 1–16.
- Cadena EA, Gaffney E. Notoemys zapatocaensis, a new sidenecked turtle (Pleurodira: Platychelyidae) from the Early Cretaceous of Colombia. Am. Mus. Novit. 2005;3470:1–19.
- Cadena EA. First record of eucryptodiran turtles from the early Cretaceous (Valanginian), at the northernmost part of South America. S. Am. J. Herpetol. 2011;6:49–53.10.2994/057.006.0107
- Maisey JG. Growth and form of finspines in hybodont sharks. Palaeontology. 1978;21:657–666.
- Huxley TH. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proc. Zool. Soc. London. 1880;43:649–662.
- Bonaparte CL. Selachorum tabula analytica. Nuovi Ann. Sci. Nat. Bologna. 1838;1:1–15.
- Maisey JG. The interrelationships of phalacanthous selachians. Neu. Jb. Geol. P-M. 1975;1975:553–567.
- Owen R. Lectures on the comparative anatomy and physiology of the vertebrate animals, delivered at the Royal College of Surgeons of England in 1844 and 1846. Part 1. Fishes. London: Longman; 1846.
- Maisey JG. The anatomy and interrelationships of Mesozoic hybodont sharks. Am. Mus. Novit. 1982;2724:1–48.
- Agassiz L. Recherches sur les poisons fossils [Research on the fossil fish]. Neuchatel. 1833–1843.
- Brito PMM. Nouvelles données sur l’anatomie et la position systématique de Tribodus limae Brito & Ferreira, 1989 (Chondrichthyes, Elasmobranchii) du Crétacé inférieur de la Chapada do Araripe (NE Brésil) [New data on the anatomy and systematic position of Tribodus limae Brito & Ferreira, 1989 (Chondrichthyes, Elasmobranchii) from the lower Cretaaceous of the Chapada do Araripe (NW of Brazil)]. Géobios. 1992;25:143–150.