
Abstract
Considering a new sample of calls of Bokermannohyla oxente, we revisited its call description and acoustic diagnosis. Four males were recorded in Piatã (Bahia, Brazil). Calls were composed of a single note emitted in sequences which could be long (up to 25 s). Notes were composed by a discrete weak/short initial prefix slightly separated from a second strong/long part; besides fundamental (= dominant), 2−3 other harmonically related bands were found. No prefix or harmonic structure were previously reported to B. oxente. Our record extends about 50 km the distribution of the species to west. The call of B. oxente is particular by having the weak/short initial prefix. Bokermannohyla juiju and B. oxente share the differential feature of releasing calls at high rates and in long sequences. Bokermannohyla itapoty also seems to have a higher dominant frequency than B. oxente.
Considerando-se uma nova amostra de cantos de B. oxente, revisitamos sua descrição de canto e diagnose acústica. Quatro machos foram gravados em Piatã (Bahia, Brasil). O canto é composto de uma única nota emitida em sequências que podem ser longas (até 25 segundos). As notas são compostas por um prefixo inicial discreto, fraco e curto, ligeiramente separado de uma segunda parte, mais longa e forte; além da fundamental (= dominante) encontramos 2−3 outras bandas harmônicas. A presença do prefixo e de harmônicos ainda não eram conhecidas para B. oxente. Nossa amostra também estende cerca de 50 km a distribuição da espécie para o oeste. O canto de B. oxente é particular por ter o prefixo fraco/curto inicial. Bokermannohyla juiju e B. oxente compartilham a característica diferencial de cantar em taxas elevadas e em sequências longas. Bokermannohyla itapoty também parece ter uma maior frequência dominante do que B. oxente.
Introduction
Among frogs, the advertisement call is a species-specific signal involved in mating recognition with well-established applications in taxonomy.[Citation1,2] Bokermannohyla oxente Lugli and Haddad, 2006 [Citation3] was described from localities in the Chapada Diamantina (Bahia, Brazil) and presently is placed in the B. pseudopseudis species group (sensu Faivovich et al. [Citation4]). Its advertisement call was described along with the species [Citation3] based on analogical recordings of two males. Considering the availability of a new sample of calls of B. oxente, we revisited its call description and acoustic diagnosis and found that an improvement of both is necessary, what we intend to provide here.
Methods
Four males were recorded in Piatã (Chapada Diamantina, Bahia, Brazil; 80 km southwest from type locality) in 4–6 January 2013 between 19:36 and 21:10 h. Calls were recorded with a Marantz PMD 671 (Sennheiser ME67/K6 microphone) or a MicroTrack (ME66/K6 microphone), all set to 44.1 or 48 kHz sampling rate and 16-bit resolution. Call records were performed from close range (100−300 cm) from calling males. Acoustic variables were analyzed with Raven Pro 1.5, 64-bit version [Citation5]; terminology for call features were mostly according to Raven’s manual. A 200-Hz high-pass band filter was applied for call analyses and figuring. Sound figures were obtained with Seewave 1.6.4 [Citation6] R package,[Citation7] v. 3.2.2, settings were Hanning window, 85% overlap, and 256 points resolution.
Lugli and Haddad [Citation3] recognized calls of B. oxente as composed by longer (20–58) and shorter (17–37 ms) notes but considering the overlap which precludes objective classification and that we did not find such variability in our sample, we avoided such differentiation. As to B. juiju Faivovich, Lugli, Lourenco and Haddad, 2009 (call described in [Citation8]), we think that, to B. oxente, taking individual notes as the call is in closer accordance with the mechanistic definition of call in frogs.[Citation9,10] Measured call (= note; = prefix + suffix + interval) features were: (1) prefix and (2) suffix durations; (3) prefix–suffix interval; (4) note interval, (5) peak frequency; (6) lower frequency; (7) higher frequency; and (8) note rate (considering sections of 5 or 10 s of active calling). Descriptive statistics was based in mean values of individual males.
Examined voucher specimens and recorded calls are in the collection of frogs of the Universidade Federal de Uberlândia (AAG-UFU) (Appendix 1). We tried to reanalyze the original records of Lugli and Haddad [Citation3] but they are apparently lost (C.F.B. Haddad pers. comm.). A footage of a B. oxente calling male by Herpetoorg (youtube.com/user/Herpetoorg) is available in: youtube.com/watch?v=40kZ8qP2hrM.
Results
Adult male specimens from our study site (Piatã, Bahia) in Figure .
Figure 1. Adult males of Bokermannohyla oxente from Piatã, Bahia, Brazil. Above AAG-UFU 1684, below AAG-UFU 1685. Both vouchers of call records.

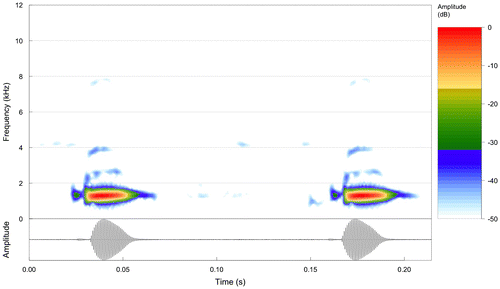
Advertisement call. Table and Figure . Call composed of a single note. Notes emitted in sequences of variable length, which could be very long (up to 220 notes, 25 s). The first 2–5 (introductory) notes could be weaker (lower amplitude) than the remaining ones; after introduction, notes varied just slightly in amplitude regarding one another. Notes were composed by a discrete weak/short prefix slightly separated from a second unpulsed strong/long part, here called suffix; besides fundamental (= dominant), 2–3 other harmonically related bands could be noticed.
Table 1. Call features of Bokermannohyla oxente based on our sample from Piatã (Bahia; N = 4 males, 80 analyzed calls) and on types (Lençóis, Bahia, topotypes, N = 2 males) in Lugli and Haddad.[Citation3]
Figure 2. Above spectrogram and below oscillogram of a sequence of two calls of Bokermannohyla oxente (Piatã, Bahia, Brazil). Notice the weak prefix of each call and the harmonic structure (second and forth harmonics can be seen here). January 6 2013, air and water 22 °C. Record voucher AAG-UFU 1684. Call file Bokerm_oxentePiataBa2aAAGm671.
Discussion
Lugli and Haddad [Citation3] found no prefix or harmonic structure in their call sample of B. oxente, in which they were followed by Taucce et al. [Citation8]. The differences in relation to our sample can be attributed to eventualities such as background noise, large distances of males from microphones, undesired effects resulting from digitalization and/or settings for figuring. The presence of the prefix is particularly problematic because it is very weak and could quite easily be missed during recording or analyses. Despite these differences, we have no reason to doubt our attribution of calls to B. oxente as call duration, dominant frequencies and call rate are in agreement between samples. Accordingly, besides call, our specimens are also in agreement with B. oxente description (data not shown). Our sample from Piatã extends about 50 km the known distribution of the species to the west.
Comparisons of the advertisement call of several species of Bokermannohyla are in Carvalho et al. [Citation11,12] and in Taucce et al. [Citation8]. According to our data, the call of B. oxente is particular by having the weak prefix. Our call redefinition to B. oxente turns it more similar to that of Bokermannohyla juiju and B. itapoty Lugli and Haddad, 2006 [Citation13] than previously thought. Bokermannohyla juiju and B. oxente share the differential feature of releasing calls in long sequences and at high rates, with B. oxente being even faster (4 times). The syntopic B. itapoty also seems to have a higher dominant frequency (1680–3300 Hz,[Citation13] than B. oxente (peak at 1300 Hz).
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
This work was supported by Fapemig; CNPq.
Acknowledgements
Thiago R Carvalho helped in the field works and critically read the draft. The Bioacoustics Research Program, Cornell Laboratory of Ornithology, provided free licenses of Raven Pro. A research grant to AAG by CNPq. Fellowships to TB by FAPEMIG and to LBM by CNPq. Collection permits by ICMBio /SISBIO (#30059).
References
- Duellman WE, Trueb L. Biology of amphibians. New York: McGraw-Hill; 1986, p. 670.
- Angulo A, Reichle S. Acoustic signals, species diagnosis, and species concepts: the case of a new crypitic species of Leptodactylus (Amphibia, Anura, Leptodactylidae) from the Chapare region, Bolivia. Zool J Lin Soc. 2008;152:59–77.10.1111/zoj.2008.152.issue-1
- Lugli L, Haddad CFB. A new species of the Bokermannohyla pseudopseudis group from central Bahia, Brazil (Am phibia, Hylidae). Herpetologica. 2006;62:453–465.
- Faivovich J, Haddad CFB, Garcia PCA, et al. Systematic review of the frog family Hylidae, with special reference to Hylinae: phylogenetic analysis and taxonomic revision. Bull Am Mus Nat Hist. 2005;294:1–240.10.1206/0003-0090(2005)294[0001:SROTFF]2.0.CO;2
- Bioacoustics Research Program. Raven Pro: interactive sound analysis software. Version 1.5; 2012 [cited 2016 Feb 7]. Available from: http://www.birds.cornell.edu/brp/raven/RavenOverview.html
- Sueur J, Aubin T, Simonis C. Seewave, a free modular tool for sound analysis and synthesis. Bioacoustics. 2008;18:213–226.10.1080/09524622.2008.9753600
- R Core Team R Foundation for Statistical Computing. Version 3.1.0; 2014 [cited 2015 Mar 19]. Available from: http://www.R-project.org
- Taucce PPG, Pinheiro PDP, Leite FSF, et al. Advertisement call and morphological variation of the poorly known and endemic Bokermannohyla juiju Faivovich, Lugli, Lourenço and Haddad, 2009 (Anura, Hylidae) from central Bahia, Brazil. Zootaxa. 2015;3915(1):099–110 [cited 2016 Feb 7]. doi:10.11646/zootaxa.3915.1.410.11646/zootaxa.3915.1
- McLister JD, Steven ED, Bogart JD. Comparative contractile dynamics of calling and locomotor muscle in three hylid frogs. J Exper Biol. 1995;198:1527–1538.
- Robillard T, Hobel G, Gerhardt HC. Evolution of advertisement signal in North American hylid frogs: vocalizations as end products of calling behavior. Cladistics. 2006;22:533–545.10.1111/cla.2006.22.issue-6
- Carvalho TR, Giaretta AA, Magrini L. A new species of the Bokermannohyla circumdata group (Anura, Hylidae) from southeastern Brazil, with biocaustic data on seven species of the genus. Zootaxa. 2012;3321:37–55.
- Carvalho TR, Teixeira BFV, Martins LB, et al. New bioacoustic and distributional data on Bokermannohyla sapiranga, Brandão et al., 2012 (Anura, Hylidae): revisiting its diagnosis in comparasion with B. Pseudopseudis (Miranda – Ribeiro, 1937). Zootaxa. 2013;3746:383–392 [cited 2016 Feb 7]. doi:10.11646/zootaxa.3746.2.810.11646/zootaxa.3746.2
- Lugli L, Haddad CFB. A new species of Bokermannohyla (Anura, Hylidae) from central Bahia. Brazil. J. Herpetol. 2006;40:7–15.
Appendix 1.
Examined material
(A) Call recordings from the A. A. Giaretta record files (N = 4 males);
Bokerm_oxentePiataBA1a-cAAGm671.wav;
Bokerm_oxentePiataBA2a-bAAGm671.wav (voucher AAG-UFU 1684);
Bokerm_oxentePiataBA3aLBM_AAGmt.wav;
Bokerm_oxentePiataBA4a-bLBM_AAGmt.wav (voucher AAG-UFU 1685).
Associate Editor: Guayasamin Juan Manuel.