
Abstract
Bryde’s whales aggregate in the Galápagos Archipelago in an area off the north coast of Isla San Cristóbal during periods of upwelling. Seasonally, these whales are locally abundant off San Cristóbal between May and November, except in years when El Niño conditions occur. Field surveys were performed in summers 2013, 2014, and 2016 around San Cristóbal to search for the presence of Bryde’s whales, and to document their spatial relationship with 20 °C isotherm topography. Our surveys found that most sightings of these whales occurred in locations within 3–6 km of shore (water depths 70–300 m), where the 20 °C isotherm depth shoaled to an average of 31 m below the surface. The presence of Blue whales (Balaenoptera musculus), as well as Bryde’s whales was documented during 2016 upwelling. Mother-calf pairs of Bryde’s and Humpback whales were also documented off San Cristóbal in summers 2013 and 2016 and 2014, respectively. We suggest that our three summers of sightings of Bryde’s whales in an area of upwelling indicates that Isla San Cristóbal is important habitat for individuals and mother/calf pairs alike.
Introduction
The unique oceanographic characteristics around the Galápagos Islands provide rich habitat for several migratory and resident cetacean species. Nearshore environments and upwelling areas in particular create favorable conditions for large baleen whales, such as Bryde’s whales (Balaenoptera brydei) and Blue whales [Citation1]. Bryde’s whales are a small- to medium-size baleen whale that has been reported inhabiting a wide range of tropical/subtropical environments. In the Galápagos Archipelago, Bryde’s whales are most commonly seen around the western islands, where rich primary productivity is stimulated by topographically induced upwelling. However, they are also frequently observed along the northern coast of the eastern-most island of San Cristóbal [Citation2]. In Galápagos, Bryde’s whales are typically solitary but they have also been observed in groups of two to three individuals [Citation3]. O’Hern et al. [Citation4] reported seeing Bryde’s whales in summer 2010 while surveying for marine mammals and tuna-dolphin-bird aggregations along the north coast of Isla San Cristóbal in the Galápagos Marine Reserve (GMR). During this study, Bryde’s whales were observed more frequently along the north coast of San Cristóbal (nine sightings) than along the south coast (two sightings), even though most of the island margin area from SW to NE along the north coast is orographically blocked from the SE Trade Winds that promote wind-induced upwelling elsewhere in Galápagos.
The aim of this study was to gain a better understanding of the relationship between sightings of Bryde’s whales and patterns of upwelling and 20 °C isotherm topography along the northwest coast of Isla San Cristóbal. In cooperation with the Oceanographic Institute of the Ecuador Navy (INOCAR), surveys were conducted along the north coast of San Cristóbal in summers 2013, 2014, and 2016. Given that higher prey availability is associated with upwelling, more whales should be expected during periods when cold, nutrient-rich, prey-filled waters are closest to the surface.
Of particular interest was determining whether Bryde’s whales were present in May 2016, since Denkinger et al. [Citation2] had reported these whales were not seen in Galápagos until after two years following strong El Niño events of 1981–1982 and 1997–1998 in the ETP region. ENSO (El Niño Southern Oscillation) neutral conditions characterized the surveys conducted as part of this study, although La Niña was concluding in the Eastern Tropical Pacific (ETP) just a few months before surveys in May 2013 [Citation5]. Opposite (ENSO) conditions had been present in fall-winter 2013–2014 before surveys in July 2014, and again in fall-winter 2015–2016 before surveys in May 2016 [Citation6].
Methods
Near-surface temperature was surveyed and the presence/absence of Bryde’s whales was noted along the north coast of Isla San Cristóbal in summers 2013, 2014, and 2016. Surveys were conducted from an Ecuador Navy small boat in May 2013, a fishing boat in July 2014, and from the Navy vessel LAE Sirius in May 2016. Each summer, near-surface temperature, salinity, and depth were profiled using an internally recording SeaBird 19Plus SeaCat CTD. Profiling depth was increased each field season: the CTD was lowered to reach the 20 °C isotherm to a maximum depth of 30 m in 2013, 50 m in 2014, and 100 m in 2016. CTD cast depth was increased each year to reach the 20 °C isotherm at all locations in the multi-year survey area.
While underway and stopped on station to gather CTD profiles, whales on the surface were observed using Bushnell 7 × 35 binoculars. When spouts were sighted, digital photos of the whales on the surface were taken with a Canon EOS digital camera with 75–300 mm image-stabilized telephoto lens. GPS position, local time, and frequency of each sighting were recorded. While vessels were underway, whales were not approached closer than 100 m, although occasionally, whales closed this distance while the vessel was stopped. Thus, photo identification techniques used were non-invasive; the goal was to facilitate data gathering on the whales in conjunction with physical oceanographic data collection. ArcGIS and ArcMap software were used to create the summary maps of station locations and whale sightings reported in this paper.
Results
In May 2013, four Bryde’s whales were seen at two locations off the north coast of Isla San Cristóbal near an offshore dormant volcanic tuff cone called Leon Dormido, also known as Kicker Rock, and two groups of others at two other locations to the northeast of this feature (Figure ). Sightings ranged 3–6 km offshore; water depths, where whales were seen ranged 70–300 m. Surface temperatures of three CTD casts conducted within the area of whale sightings ranged 21.2–21.9 °C; surface salinity varied very little (35.06–35.07) at all three locations as did subsurface salinity at the depth of the 20 °C isotherm (35.01–35.08). The wind-mixed layer in May 2013 extended only a few meters deep resulting in a similarly shallow thermocline. The depth of the 20 °C isotherm, which marks the base of the sharpest part of the vertical temperature gradient in the upper 100 m, ranged 16–25 m (Figure ). In all, six whales or groups of whales were seen along a transect distance of 25 km. Where whales were sighted, the average depth of the 20 °C isotherm was 23 m (range 17–29 m: Table ).
Figure 1. CTDs conducted around Isla San Cristóbal along with sightings of whales in summers 2013, 2014, and 2016. Source: The Authors.
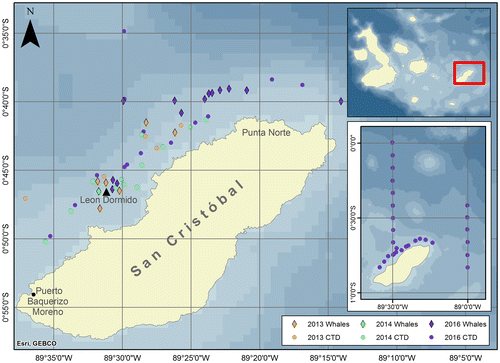
Figure 2. Temperature profiles in the upper 30 m in the area along the north coast of Isla San Cristóbal where Byrde’s whales were sighted in May 2013.
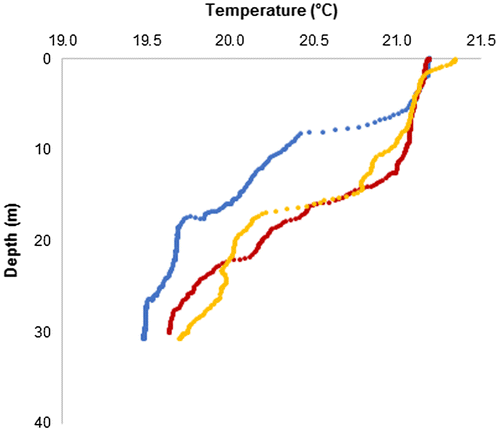
Table 1. Summary of whale sightings for summers 2013, 2014, and 2016 and corresponding oceanographic data collected near sightings.
In July 2014, just one Bryde’s whale was seen. This individual was sighted near Leon Dormido, close to where Bryde’s whales had been seen in 2013 although the total transect searched distance was 28 km. A humpback whale (Megaptera novaeanaliae) with its calf was also sighted in July 2014, in very shallow water (20 m), inshore and SW of Leon Dormido. After the 2013–2014 ENSO events, surface temperatures along the north coast were uniformly warmer than in the previous summer, and the depth of the 20 °C isotherm had a much wider range than in 2013, (17–44 m); where the Bryde’s whale was seen, 20 °C was at 22 m (Figure ). Surface salinity along a SW–NE transect was fresher than in summer 2013; at the four SW CTD stations (Figure ), surface salinity ranged 34.72–34.87, while surface salinity at NE CTDs ranged 34.84–35.02. However, at the depth of the 20 °C isotherm, salinity was similar at all eight CTD stations, ranging from 35.02 to 35.05 in July 2014 (Table ).
Figure 3. Section plot of temperature in the upper 50 m off the north coast of Isla San Cristóbal in July 2014, starting offshore of Puerto Baquerizo Moreno in the SW (left side of graph) to Bahia Sardinella in the NE (right side of graph).
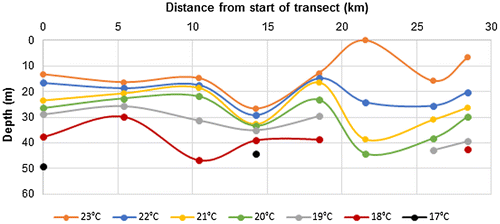
In May 2016, Bryde’s whales were again seen near Leon Dormido, and others were seen in association with blue whales during surveys near the NE tip of the island offshore of Punta Norte (Figure ). In total, 12 individuals or groups of whales were sighted at eight locations in May 2016, along a transect length of 47 km off the north shore that was searched on three separate dates, and one individual was seen in deeper water to the north. The range in depth of the 20 °C isotherm where whales were sighted in May 2016 was 8–51 m (Table ). Surface salinity all along the north coast was high and similar to May 2013 with values ranging from 34.9 to 35.2. However, a transect plot of isotherm depths along the north shore (Figure (a)) illustrates that during summer 2016, thermocline depth decreased dramatically moving towards the NE tip of the island. The depth of the 20 °C isotherm ranged from 50 to 60 m offshore of Puerto Baquerizo Moreno moving toward Leon Dormido, but this shoaled to 36–49 m around Leon Dormido and then decreased to just 8–11 m off Punta Norte. As in previous summers, salinity of the 20 °C subsurface water remained relatively constant at 35.0–35.1 (Table ).
Figure 4. Section plots of temperature in the upper 50 m May 2016, (A) from SW (left) to NE (right) along the north coast; (B) along 89.5°W (N to S), (C) along 89.0°W (N to S).
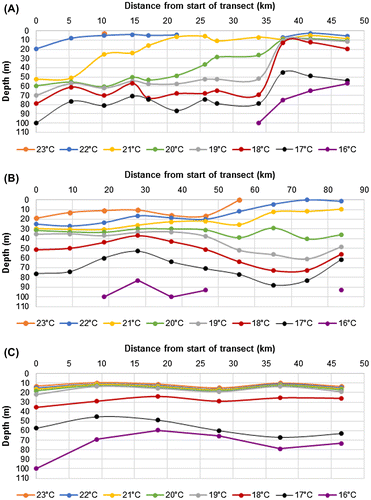
Moving from N to S along a line of 10 CTD stations conducted along 89.5°W from the 0°S to 00° 40′S (Figure (b)), the depth of the 20 °C isotherm ranged 28–42 m with an average depth of 30 m. On a similar transect south along 89.0°W between 00° 25′S to 00° 50′W, the depth of the 20 °C isotherm ranged 12–19 m with an average depth of 15.4 m (Figure (c)). A wire angle of >15° on the line to the CTD at depth indicated that the subsurface eastward-flowing Cromwell Current was quite strong below a depth of about 30 m to the north of 00° 20′S Latitude along 89.5°W Longitude, and to the north of 00° 35′S Latitude along 89.0°W (Figure ).
Discussion
The GMR lies at the confluence of two westward-flowing ocean currents and an eastward-flowing countercurrent. This confluence creates a doming (upwelling) of cold, nutrient-rich mid-water that, along with an input of iron from the islands themselves, enhances the primary production of this ETP region [Citation7]. The South Equatorial Current (SEC, or Panama Current) flows west at the surface between 5°N and 5°S, and the Peru Current (PC, or Humboldt Current) flows west to the south of the SEC. The South Equatorial Countercurrent (SECC, or Cromwell Current) flows east at a depth of 30–100 m in the confluence between the SEC and the PC. Where the SECC meets the westernmost islands in the GMR, this undercurrent shoals to create high-nutrient near-surface conditions west of Isla Fernandina. From there, the SECC bifurcates to flow along the north side and along the south side of the GMR [Citation8]. In the north, the SECC is topographically steered by shoaling bathymetry to upwell on to the continental margin near Isla San Cristobal. It is this topographically steered upwelling of the SECC off San Cristobal that creates favorable habitat for whales there.
Examining the topography of the 20 °C isotherm has been previously used to describe marine mammal distribution patterns in the ETP [Citation10]. The depth of the subsurface 20 °C isotherm and salinity of 35.0–35.1 that is a proxy for the core of the SECC [Citation9] usually shoals to 31 m of the surface along the north and east side of Isla San Cristobal. Seasonally and interannually, the depth of the 20 °C isotherm off Isla San Cristóbal can be much deeper. In September 2015, in CTD casts taken along 89.0° W during the height of the 2015–2016 ENSO event, the depth of the 20 °C isotherm at each of three locations at 00°S, 00° 30′S, and 00°59′S was 55–60 m (INOCAR, Hydrographic Division unpublished data archives of annual CTD surveys by B/O Orion).
Our surveys found that, in general, years when the 20 °C isotherm was closer to the surface were years when more Bryde’s whales were spotted around Isla San Cristóbal. In summers 2013 and 2016, when depth of the 20 °C isotherm off the north coast there averaged 20 and 33 m, respectively, Bryde’s whales were observed frequently. In contrast, in 2014, when the depth of the 20 °C isotherm from Leon Dormido and to the northeast generally ranged 30–44 m, only one whale was observed. These results suggest that whales are attracted to the area during periods of upwelling, where it is likely that increased productivity on fine scales of space (i.e. station-to-station distances of a few kilometers corresponding to small vertical differences in 20 °C depth) is providing an important food source for these cetaceans. It is believed these neritic whales capture their food by looking for aggregations of fish, shrimp, and other potential prey. Other members of the genus Balaenoptera have also been found to forage in coastal upwelling environments in the Pacific Ocean [Citation11][Citation12].
Seasonally, upwelling along the north coast of Isla San Cristóbal occurs between May and November, during the so-called garua season [Citation13]. This is the time of year when Bryde’s whales are most commonly seen by the Galápagos National Park naturalists that accompany the tour boats and dive boats that make day trips along the north shore (O. Ricaurte, personal communication). This anecdotal evidence of increased Bryde’s whale sightings around San Cristóbal during summer months matches the results of the surveys presented here.
During the course of this study, several other marine mammal species were spotted, including Humpback whales (Megaptera novaeangliae), Blue whales (Balaenoptera musculus), Galápagos sea lions (Zalophus wollebaeki), and several pods of dolphins. Some of these marine mammals were observed very close to shore in water depths of 20 m or less (i.e. most of the sea lions, some of the dolphin sightings, and the mother-calf pair of Humpback whales in 2014). Galápagos National Park rangers also spotted Orcas (Orcinus orca) in our survey area during 2015, when the rangers were escorting boat charters by scuba divers transiting along the north coast of San Cristóbal (O. Ricaurte, personal communication).
Because whales were abundant off San Cristobal just 3–4 months after the strong 2015–2016 ENSO event had ended indicates that Isla San Cristóbal is a fragile but vital ecosystem for Bryde’s whales during multiple life stages. The presence of mother-calf pairs of Bryde’s whales off the north coast of Isla San Cristóbal was noted during surveys in May 2013 and 2016 (Figure (b)), so it is likely that this location is important to the reproductive biology of this species. Because of their elusive nature, little is known about where Bryde’s whales sighted in Galápagos might breed, or about their seasonal or interannual migration patterns. To better understand how Bryde’s whales are using the habitat around San Cristóbal, it is critical that wide-scale oceanographic data be collected several times a year in addition to more accurate information on the whales themselves through using either mark-recapture techniques or photo identification methods. But because Bryde’s whales do not fluke up when they dive to forage, continued/extended photography will have to focus on differences in coloration of the back in the area adjacent to the dorsal fin, and/or to nicks or other wear in the fin itself. Nevertheless, collecting dorsal fin photographs for photo identification, if possible in coordination with tagging animals for satellite telemetry, could be particularly valuable in determining if individuals display interannual site fidelity and characterizing how this population of Bryde’s whales uses this habitat. With extreme events like this most recent ENSO expected in higher frequency as a result of climate change [Citation14,15], now, more than ever, it is vital to obtain a base level of understanding of this environment and its transient inhabitants in order to map future changes both locally and ocean-wide.
Figure 5. Telephoto photography of whales seen off Isla San Cristóbal in 2013 and 2016: (A) Typical sighting of a Bryde’s whale on the surface (2013); (B) A mother-calf Bryde’s whale pair photographed just off Leon Dormido in 2016; (C) A Blue whale close to shore in 2016 (beach is visible at top of photo).

Geolocation information
Data for this study were collected around the coast of Isla San Cristóbal, Galápagos, Ecuador. Approximate coordinates are: 89°45′0″ W to 89°0′0″ W and 1°0′0″ S to 0°0′0″.
Author contribution
DB is the corresponding author and was principal investigator for Galápagos fieldwork 2013, 2014, and 2016. SD and LM were graduate students at Texas A&M who participated in Galápagos in 2013/2014 and 2014, respectively. MN was an undergraduate student volunteer from USF Quito for 2014 fieldwork. AG and ZL were undergraduate Texas A&M students and REU participants for 2016 fieldwork. MS was a Lieutenant in Ecuadorian Navy and Captain of the LAE Sirius for Galápagos fieldwork 2016.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
This work was supported by Texas A and M University [grant number 02-241588-15022].
Acknowledgments
We thank Julia O’Hern and Michelle Sculley for sharing their survey data from summer 2010 with us, and we thank Galápagos National Park Ranger Onivid Ricaurte for sharing with us his observations about seasonal occurrence of Bryde’s whales and killer whales off Isla San Cristobal. We also thank the Director of INOCAR (Humberto Gomez Proano) for his support of CTD survey work from Ecuador Navy ships. We especially thank the INOCAR cadets who in 2013, 2014, and 2016 made the oceanographic data collection during surveys. Finally, we thank Bruce Mate and Tom Jefferson for confirming that Figure (c) is that of a Blue whale.
Associate Editor: Juan M. Guayasamin.
Notes
Research was conducted at Isla San Cristóbal, Galápagos. We thank the Director of INOCAR, Humberto Gomez Proaño, for his support of CTD survey work from Ecuadorian Navy ships, and we also thank the College of Geosciences at Texas A&M University for travel support of U.S. participants.
References
- Alava JJ. Carbon productivity and flux in the marine ecosystems of the Galápagos Marine Reserve based on cetacean abundances and trophic indices. Rev Biol Mar Oceanogr. 2009;44:109–122.
- Denkinger J, Ona D, Alarcon G, et al. From whaling to whale watching: cetacean presence and species diversity in the Galápagos Marine Reserve. In: Walsh SJ, Mena CF, editors. Science and Conservation in the Galápagos Islands. New York (NY): Springer; 2013. p. 217–235.10.1007/978-1-4614-5794-7
- Palacios DM. Marine mammal research in the Galápagos Islands: the 1993–94 Odyssey expedition. Puerto Ayora, Galápagos, Ecuador: Galápagos National Park Service and Charles Darwin Research Station; 1999.
- O’Hern JE, Sculley MJ, Smith KJ, et al. Marine mammal distribution in Ecuador: Surveys of opportunity as a means of monitoring relative abundance. Latin Am J Aquatic Mammals. In press.
- Historical El Niño La Niña Watch [Internet]. Pasadena (CA): NASA Jet Propulsion Laboratory, California Institute of Technology; 2016 [ cited 2016 August 24]. Available from https://sealevel.jpl.nasa.gov/science/elninopdo/elnino/
- ENSO: Recent Evolution, Current Status, and Predictions [Internet]. College Park (MD): NOAA Climate Prediction Center/NCEP; 2016 [ cited 2016 August 24]. Available from https://www.cpc.ncep.noaa.gov/products/analysis_monitoring/lanina/enso_evolution-status-fcsts-web.pdf
- Palacios DM. Factors influencing the island-mass effect of the Galápagos Archipelago. Geophys Res Ltrs. 2002;29:2134–2137.
- Pak H, Zaneveld JRV. The Cromwell Current on the east side of the Galápagos Islands. J Geophys Res. 1973;78(33):7845–7859.10.1029/JC078i033p07845
- Fiedler PC. Comparison of objective descriptions of thermocline. Limnol Oceanogr: Methods. 2010;8:313–325.10.4319/lom.2010.8.313
- Reilly SB. Seasonal changes in distribution and habitat differences among dolphins in the eastern tropical Pacific. Mar Ecol Prog Ser. 1990;66:1–11.10.3354/meps066001
- Reilly SB, Thayer VG. Blue whale (Balaenoptera musculus) distribution in the Eastern Tropical Pacific. Mar Mam Sci. 1990;6(4):265–277.10.1111/mms.1990.6.issue-4
- Croll DA, Marinovic B, Benson S, et al. From wind to whales: trophic links in a coastal upwelling system. Mar Ecol Prog Ser. 2005;289:117–130.10.3354/meps289117
- Houvenaghel GT. Oceanographic conditions in the Galápagos Archipelago and their relationships with life on the islands. In: Boje R, Tomczak M, editors. Upwelling Ecosystems. Berlin: Springer-Verlag; 1978. p. 181–200.10.1007/978-3-642-66985-9
- Cai W, Borlace S, Lengaigne M, et al. Increasing frequency of extreme El Nino events due to greenhouse warming. Nat Clim Change. 2014;4:111–116.10.1038/nclimate2100
- Gergis JL, Fowler AM. A history of ENSO events since A.D. 1525: implications for future climate change. Clim Change. 2009;92(3–4):343–387.10.1007/s10584-008-9476-z