
ABSTRACT
This work is the first attempt to assess the biodiversity of the Hydrozoa in the Archipiélago de Bocas del Toro (Panamá, Caribbean Sea) using morphology and molecular taxonomy, and to produce field identification tools to help future identification and monitoring efforts in the area.
We sampled, identified, vouchered, and barcoded 112 specimens of Hydrozoa from shallow coastal waters (0–22 m depth) in the Archipiélago de Bocas del Toro. The specimens belong to 70 taxa, of which 53 were identified at the species level, and 17 were identified at the genus or family level. We produced 64 sequences of the large ribosomal subunit of the mitochondrial RNA (mt lsu-rRNA, 16S), the genetic marker generally used for barcoding Hydrozoa. We updated the local checklist that now comprises 118 species, and produced 87 detailed taxon identification tables that display species descriptions augmented with pictures, geographic distribution (worldwide and in Bocas del Toro), GenBank accession numbers for the 16S mitochondrial gene, and a synopsis of the families they belong to.
Introduction
Hydrozoa are an inconspicuous and often overlooked class of the phylum Cnidaria [Citation1]. Most hydrozoans have a complex life cycle characterized by a progression of three life stages: a short-living larva (the planula), generally metamorphosing into a benthic colonial stage (the polyp), and a pelagic sexual stage (the medusa stage) asexually budded off from the polyp [Citation2]. The life cycle can be shortened into a bi-phasic cycle, by reduction or complete suppression of either the polyp or the medusa stage [Citation3]. With more than 3,700 described species [Citation4] hydrozoans are structurally and functionally important members of benthic and planktonic communities [Citation5; 1]. Work on Hydrozoa has been hampered by the scarcity of taxonomic expertise, which has dramatically declined over the past two decades [Citation6,Citation7]. Also, because polyps and medusae require different expertise and each follow their own identification rules, linking both to a single species has proven difficult and has further hampered cohesive taxonomic revisions. In the last 10 years molecular tools have contributed significantly to hydrozoan taxonomy and have shown that selected gene sequences may be necessary, in combination with traditional taxonomy, to correctly identify cryptic species and disentangle taxonomic confusion [Citation8–Citation12].
Hydrozoa from the Atlantic/Caribbean coast of Panama are abundant but scarcely studied. The Archipiélago de Bocas del Toro, Panamá is located on the NW Caribbean shore of Panama, close to the Costa Rican border. It consists of more than 68 small islands and mangrove keys and is characterized by diverse ecosystems, from mangrove dominated shallow water to coral reefs and sea grass meadows (see for a map). To date, 79 nominal species have been reported in the Bocas del Toro region [Citation13]. However, proper descriptions and species identification tools are lacking, inadequate, or scattered in old and hard to access articles, thus hindering the ability of nonexperts to identify species of interest in this area.
Figure 1. Map of the Bocas del Toro archipelago, with sampling localities.
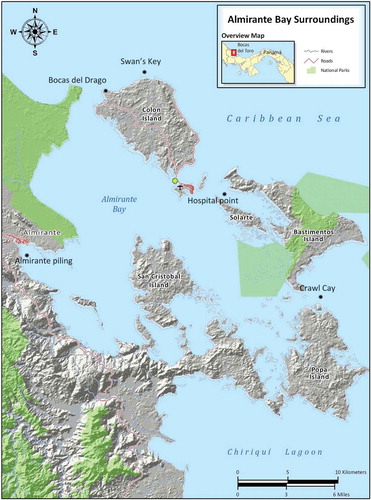
This work is a result of the Taxonomic Training workshop held in July 2015 in Bocas del Toro, Panama, organized by Smithsonian Tropical Research Institute as part of a National Science Foundation Advancing Revisionary Taxonomy and Systematics (ARTS) grant. During the workshop, in which 14 students participated, we collected shallow water Hydrozoa around the Archipiélago de Bocas del Toro, Panamá. Collections targeted both pelagic medusa with plankton tows and benthic polyps with snorkeling. In this paper, we present an updated checklist of hydrozoan biodiversity in the Archipiélago de Bocas del Toro,, augmented by [I] a DNA barcoding database consisting of sequences of a ~ 600bp gene fragment of the mitochondrial large ribosomal RNA subunit (mt lsu-rRNA, 16S), and [II] taxon identification tables. The 16S gene has been successfully used for taxonomic revisions and is considered the Hydrozoa “barcoding” molecule [Citation10,Citation11,Citation14–Citation17]. Both the barcoding data and the taxonomic identification sheets were produced to aid future biodiversity inventory efforts in the region.
Material and methods
Collection
Hydrozoa were collected during the Tropical Taxonomy Training course on systematics and biology of Hydrozoa (Cnidaria) held at the Smithsonian Tropical Research in Bocas del Toro, Panama from 7/7/2015 to 7/21/2015. A total of 16 people (two instructors and 14 students) were in the field during every sampling effort. A total of 11 sites were sampled. Localities sampled were: Bocas del Toro Marine Station docks/weather station, Punta Hospital, Crawl Cay, Bocatorito Bay vicinity, San Cristobal, Vicinity of Manuguar Cay, Punta Caracol, Swans Cay, Bastimento vicinity of “Casa Verde”, Bocas del Drago, Pandora, Almirante (Quary’s point) ( and ). Polyps were collected by snorkeling (0–8 m depth) or SCUBA diving (18–22 m depth), and medusae by plankton towing using a net with 280µm mesh size. Plankton tows were carried out near the Bocas del Toro Station docks and weather station.
Table 1. Samples collected during the 2015 Hydrozoa workshop held in Bocas del Toro, Panama. Date of collections, sample ID, species identified, type of fixative used for preservation (ethanol or formalin), GenBank accession number, and location within the Bocas del Toro archipelago are reported. In the table N = No, Y = Yes.
Polyps and medusae were sorted in the laboratory and identified to the lowest taxonomic level using appropriate taxonomic literature [e.g. 16–20]. When possible, polyps from each colony where divided in two vials and preserved in both formalin (for morphological analyses) and 99% ethanol (for genetic analyses). Vouchered specimens were deposited at the Museum of Universidad the Panamá, Panama City, Panama. When little tissue was available, specimens were preserved in ethanol only and used for molecular analyses. DNA samples are currently at the University of Texas A&M at Galveston. Pictures of live specimens featured colony, polyps, medusae (or equivalent reproductive structures), and other morphological structures useful for identification.
Barcoding
Genomic DNA was extracted using standard techniques followed by ethanol precipitation [Citation8]. A ~ 600bp fragment of the large ribosomal subunit of the mitochondrial RNA (lsu-rRNA, 16S) was amplified using SHA and SHB primers [Citation23], Polymerase Chain Reaction (PCR), and conditions as described in Miglietta [Citation15]. The PCR product was run on a 2% agarose gel stained with SybrGreen I nucleic acid gel stain (Sigma-Aldrich) to assay its quantity and quality (i.e. accessory bands). PCR products were purified using exoSAP-it (Affymetrix), following manufacturer’s instruction and used as a template for double stranded sequencing with the amplification primers. The purified DNA was sequenced at the Texas A&M Corpus Christi Genomics Core Laboratory. Sequences were analyzed in Geneious R9 (http://www.geneious.com [Citation24]) and deposited in Genbank (accession numbers MH361321 to MH361381). For definition of anatomical terms used in the ID tables, see online taxonomic glossary for Hydrozoa at https://stricollections.org/portal/misc/glossarycover.php [Citation25].
Results
We collected, vouchered, and fixed in ethanol and/or formalin 112 specimens of Hydrozoa (). We identified a total of 53 species. An additional 17 taxa could be identified at the genus or family level only due to the lack of fertile structures, small size of the colony, or, in the case of planktonic specimens, early age of the medusae ( for a complete species list).
Table 2. List of species found in Bocas del Toro during the Hydrozoa Taxonomy course in 2015. For each species we report voucher name(s), number of barcoding sequences (mt lsu-rRNA, 16S) produced per species, and whether their taxonomic description is in the Taxon Identification Tables. A total of 53 taxa were identified at the species level. At the bottom, 17 taxa identified at the genus or higher taxonomic level.
We sequenced the 16S gene for 64 out of 112 specimens belonging to 44 species. Some of the 112 specimens did not yield sufficient DNA for Polymerase Chain Reaction and thus could not be barcoded. For some species, however, multiple sequences were produced. All sequences were ~ 600bp in length and were deposited in GenBank (accession numbers MH361321-MH361332, MH361334-MH361359, MH361361-MH361381, MH374630).
Morphological, ecological, and barcoding data where assembled in Taxon Identification Tables. We produced 87 tables featuring 28 families (13 in the order Anthoathecata; 12 in the order Leptothecata, 2 in the order Trachymedusae and 1 in the Order Limnomedusae) and 55 species (Appendix 1). These 55 species represent the most common Hydrozoa found in Bocas del Toro during the workshop. For the most speciose taxa we supply identification keys (to genera and/or to species). One of the most abundant families found in the area was the Campanulariidae, with 3 genera and 9 species. Because the polyps of the three genera sampled (Clytia, Gastroblasta, and Obelia) can be easily confused we also supply identification tables for each genus.
Discussion
Check list of Bocas del Toro Hydrozoa
A comprehensive inventory of the Hydrozoa from the Caribbean coast of Panama was previously produced by Calder and Kirkendale [Citation13], who gathered three different collections acquired in 1969, 2002, and 2004, mostly from the Bocas del Toro region. They recorded 79 nominal species (of which were 17 identified at the genus or family level only) belonging to 22 families. We found 53 species, of which only 31 were in common with Calder and Kirkendale [Citation13]. We merged our own inventory with that of Calder and Kirkendale [Citation13],to produce an updated checklist of the Hydrozoa of Bocas del Toro that now comprises 118 taxa, of which 86 identified at the species level and 32 identified at the genus or higher level (). Given such a small geographical area (250 km2), this is an impressive number. For comparison, 118 species constitutes about ¼ of the total known Hydrozoa species from the Mediterranean Sea [Citation26], and more than a half of the species known from the Arctic [Citation27] or the Antarctic [Citation28]. Seven families and 12 genera are also reported for the first time in Bocas del Toro. Genera added to the checklist are: Amphynema, Cytaeis, Codonorchis, Rhizogeton, Thecocodium, Turritopsoides (?), Gastroblasta, Egmundella, Pteroclava, Cubaia, Lyriope, and Persa. Families new to the Bocas del Toro region are: Pandeidae, Cytaeididae, Cladocorynidae, Ptilocodidae, Olindiidae, Geryoniidae, Rhopalonematidae. Of the new species added to the list, of particular interest is Thecocodium sp. The genus Thecocodium was never reported in the Caribbean and was only recently recorded for the first time in the Atlantic Ocean [Citation29]. Thecocodium sp. (specimen BTH 15.64) presents unique morphological features and may represent a new species. Another species of interest is a species of the genus Coryne (specimen 15.93) found in Swans Key. The colony could not be identified at the species level; however in GenBank its 16S sequence showed 100% identity with Coryne japonica (AY512540) from New Zealand. C. japonica has been reported from the Pacific Ocean but never from the Atlantic Ocean. Our record from Bocas del Toro is the first in the Atlantic Ocean and may represent an introduced species.
Table 3. Updated checklist of the Hydrozoa of Bocas del Toro. The list includes species reported in Calder and Kirkendale [Citation13],and this paper. For the species in this paper, the voucher number is reported. At the bottom, taxa identified at the genus or higher taxonomic level.
Taxon identification sheets
Taxon Identification Sheets collate the taxonomic description of 56 species found during the Hydrozoa Taxonomy course and confidently identified at the species level. They also include a brief synopsis of the 28 families they belong to (see Tables 1–88 in Supplementary Materials. For each family we included authorship, corresponding Order, number of species (worldwide and in Bocas del Toro), morphologically similar taxa that could be mistakenly identified as member of the family of interest, and their key diagnostic characters. The species identification sheets include author, diagnostic characters of the colony and their reproductive structures (medusae, eumedusoids, or fixed gonophores), species ecology, species distribution in Bocas del Toro, number of specimens collected, pictures and, when available, GenBank accession numbers for their 16S sequence(s). These species identification sheets assemble in simple format information that can be used for a correct identification. The morphological description, pictures, and link to their 16S barcoding sequence represent a comprehensive display of information that integrates traditional and modern taxonomy into a practical tool to aid identification of the most commons species of Hydrozoa in the shallow waters of the Archipiélago de Bocas del Toro, Panamá.
Conclusive remarks
Knowledge on the local biodiversity is an essential pre-requisite for the monitoring and management of environmental assets and ecosystem health worldwide. The present inventory of the marine hydrozoan fauna in the Bocas del Toro shallow water is far to be exhaustive due to the inherent limitation of our sampling efforts, based mostly on snorkeling and more rarely on SCUBA diving collections. However, the high number of recorded taxa suggests that the Caribbean Sea should be considered a region of high hydrozoan diversity. Paradoxically, taxonomy is a science at brink of extinction. The ARTS courses have been devoted not only to increase knowledge on local biodiversity, but towards the conservation and promotion of biodiversity expertise. More generally, training a new generation of taxonomists is a current challenge and a mandatory urge to understanding ecosystem functioning in face of local and global changes, and to address the needs of sustainability of humankind activities.
Author contributions
MPM and S. Piraino designed the experiments; S. Pruski produced the barcoding sequences, all authors collected samples and contributed the Taxon Identification Tables: MPM wrote the paper.
Supplemental Material
Download MS Power Point (18.9 MB)Acknowledgments
The authors would like to thank the National Science Foundation for funding. We would also like to thank the staff at the Smithsonian Tropical Research Institute’s Bocas del Toro Research Station for their support and help during the 2015 taxonomy course, the Panamanian Ministry of the Environment (MiAmbiente) for permission to conduct this research, and two anonymous reviewers for their useful comments.
Associate Editor: Federico Brown
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplementary material
Supplemental data can be accessed here.
Additional information
Funding
Related Research Data
References
- Boero F, Bouillon J, Gravili C, et al. Gelatinous plankton: irregularities rule the world (sometimes). Mar Ecol Prog Ser. 2008;356:299.
- Boero F, Bouillon J, Piraino S, et al. Asexual reproduction in the Hydrozoa. In: Hughes RN, editors. Reproductive biology of invertebrates - progress in asexual reproduction. Vol. 11. New Delhi: Oxford & IBH Publishing; 2002. p. 141.
- Bouillon J, Gravili C, Pagés F, et al. An introduction to Hydrozoa. Mémoires Du Muséum National d’Histoire Naturelle. 2006;194.
- World Hydrozoa Schuchert, P Database. 2018. Available from: http://www.marinespecies.org/hydrozoa
- Gili J-M.Coma R. 1998. Benthic suspension feeders: their paramount role in littoral marine food webs. Trends Ecol Evol 13 (8):316 –321.
- Tautz D, Arctander P, Minelli A, et al. A plea for DNA taxonomy. Trends Ecol Evol. 2003;18:70–74.
- Boero F. Light after dark: the partnership for enhancing expertise in taxonomy. Trends Ecol Evol. 2001;16:266.
- Miglietta MP, Cunningham CW. Evolution of life cycle, colony morphology, and host specificity in the family Hydractiniidae (Hydrozoa, Cnidaria). Evolution. 2012;66:3876–3901.
- Miglietta MP, Lessios HA. A silent invasion. Biol Invasions. 2009;11:825–834.
- Miglietta MP, Odegard D, Faure B, et al. Barcoding techniques help tracking the evolutionary history of the introduced species Pennaria disticha (Hydrozoa, Cnidaria). PLoS One. 2015;10:e0144762.
- Schuchert P. Species boundaries in the hydrozoan genus Coryne. Mol Phylogenet Evol. 2005;36:194–199.
- Miglietta MP, Schuchert P, Cunningham CW. Reconciling genealogical and morphological species in a worldwide study of the family Hydractiniidae (Cnidaria, Hydrozoa). Zool Scr. 2009;38:403–430.
- Calder DR, Kirkendale L. Hydroids (Cnidaria, Hydrozoa) from shallow-water environments along the Caribbean coast of Panama. Caribbean J Sci. 2005;41:476–491.
- Moura CJ, Harris DJ, Cunha MR, et al. DNA barcoding reveals cryptic diversity in marine hydroids (Cnidaria, Hydrozoa) from coastal and deep-sea environments. Zool Scr. 2008;37:93–108.
- Miglietta MP. Turritopsis fascicularis Fraser, 1943 (Cnidaria: hydrozoa): redescription and discussion of its phylogenetic position within the genus. Zootaxa. 2016;4097:426–433.
- Miglietta MP, Piraino S, Kubota S, et al. Species in the genus Turritopsis (Cnidaria, Hydrozoa): a molecular evaluation. J Zool Systematics Evol Res. 2007;45:11–19.
- Miglietta MP, Faucci A, Santini F. Speciation in the sea: overview of the symposium and discussion of future directions. Integr Comp Biol. 2011;51:449–455.
- Calder DR. Shallow-water hydroids of Bermuda: the Thecatae, exclusive of Plumularioidea (No. 154). Toronto, Ontario; Royal Ontario Museum; 1990.
- Galea HR. Additional shallow-water thecate hydroids (Cnidaria: hydrozoa) from Guadeloupe and Les Saintes, French Lesser Antilles. Zootaxa. 2010;2570(1):1–40.
- Galea HR. New additions to the shallow-water hydroids (Cnidaria: hydrozoa) from the French Lesser Antilles: martinique. Zootaxa. 2013;3686(1):1–50.
- Galea HR. On a collection of shallow-water hydroids (Cnidaria: hydrozoa) from Guadeloupe and Les Saintes, French Lesser Antilles. Zootaxa. 2008;1878:1–54.
- Calder DR. Shallow-water hydroids of Bermuda: superfamily Plumularioidea (No. 161). Toronto, Ontario: Royal Ontario Museum; 1997.
- Cunningham CW, Buss LW. Molecular evidence for multiple episodes of paedomorphosis in the family Hydractiniidae. Biochem Syst Ecol. 1993;21:57–69.
- Kearse M, Moir R, Wilson A, et al. Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28:1647–1649.
- Collin R, Fredericq S, Freshwater DW, et al. TaxaGloss - A glossary and translation tool for biodiversity studies. Biodivers Data J. 2016;e10732.
- Bouillon J, Medel MD, Pagès F, et al. Fauna of the Mediterranean hydrozoa. Scientia Marina. 2004;68.
- Ronowicz M, Kukliński P, Mapstone GM, et al. Trends in the diversity, distribution and life history strategy of Arctic Hydrozoa (Cnidaria). PloS One. 2015;10(3):e0120204.
- Mercado Casares B, Soto Àngel JJ, Peña Cantero ÁL, et al. Towards a better understanding of Southern Ocean biogeography: new evidence from benthic hydroids. Polar Biol. 2017;40:1975.
- Kubota SMeldonian S. First occurrence of a rare thecocodium medusa (anthomedusae, ptilocodiidae) from riviera beach, florida, usa. Biogeography. 2016;18:77–78.