
ABSTRACT
Of the total numbers of microsites present in an environment, only a fraction presents adequate conditions to ensure the regeneration of the forest. This study analyzes the facilitation given by the environment and the microsites in the P. tarapacana regeneration in the High Andes. We worked with 21 forests in Jujuy, Argentina, registering variables of forest structure, soil moisture and the offer of microsites. Pearson correlation analyses were done between environmental variables and forest structure of seedling and trees and shrubs. In addition, a simple ANOVA were performed using 8 categories of microsites as the main factor to analyze soil moisture and the density of seedling. The density of seedling represents 36.8% of the density of trees and shrubs. At greater slopes, the presence of seedling decreases, but no significant differences were found with altitude. A higher seedling density (201.2 and 279.5 ind.ha−1) was found in microsites under the influence of rocks that coincide with higher soil moisture, but not in microsites with litter, with the same moisture content. There was a varied offer of microsites that could be found in the study area, where the density of the seedling was positively related to the offer of microsites under the influence of rocks and was associated with their relative humidity. These results define microsites favorable for regenerating P. tarapacana which can be used to identify the most suitable areas for restoration and preservation of this species.
Resumen
Del total de micrositios presentes en un ambiente, solo una fracción presenta condiciones adecuadas para asegurar la regeneración del bosque. Este estudio analiza la facilitación dada por el ambiente y los micrositios a la regeneración de P. tarapacana en los Altos Andes. Se trabajó con 21 bosques en Jujuy, Argentina, registrándose variables de estructura forestal, humedad de suelo y la oferta de micrositios para la instalación de renovales. Se realizó un análisis de correlación de Pearson entre variables ambientales y de estructura forestal de renovales y de árboles y arbustos. Además, se realizó un ANOVA simple utilizando 8 categorías de micrositios como factor principal para analizar la humedad del suelo y la densidad de renovales. La densidad de renovales representa el 36.8% de la densidad de árboles y arbustos. A mayores pendientes la presencia de renovales disminuye, pero no se hallaron diferencias significativas con la altitud. Se encontró una mayor densidad de plántulas (201.2 y 279.5 ind.ha−1) en micrositios bajo la influencia de rocas que coindicen con una mayor humedad de suelo, pero no así en micrositios con hojarasca, con un mismo contenido de humedad. Existió una oferta variada de micrositios posibles de encontrar en la zona de estudio, donde la densidad de los renovales se relacionó positivamente con la oferta de micrositios bajo la influencia de rocas, y estuvo asociada a la humedad relativa de los mismos. Estos resultados permiten definir los micrositios propicios para la regeneración de P. tarapacana pudiendo ser utilizados para identificar las áreas más adecuadas para la restauración y conservación de esta especie.
Introducción
Los micrositios son sectores del suelo del bosque con un conjunto de características ambientales propias dadas por el suelo, el clima y los factores bióticos [Citation1,Citation2]. Del total de micrositios presentes en un ambiente, sólo una fracción presenta condiciones ambientales adecuadas que aseguran la instalación de plántulas [Citation1,Citation2]. Esta instalación puede estar dada por la cantidad de luz que llega al suelo del bosque [Citation3], la heterogeneidad del suelo mineral [Citation4] y la proximidad a plantas de mayor tamaño, las denominadas plantas nodriza [Citation5,Citation6].
En los ecosistemas alpinos los principales factores de estrés son el largo período de escasez de agua, el alto grado de evapotranspiración potencial (debido al viento y al alto nivel de radiación del suelo), la erosión eólica del suelo, su bajo contenido de nutrientes y la elevada variación térmica [Citation7–9]. A su vez, presentan baja diversidad funcional como consecuencia de las condiciones severas existentes, como pendientes altas, menor capacidad de retención de agua y mayor erosión del suelo [Citation10]. Los sitios abiertos de estos ecosistemas muestran mayores dificultades para la germinación de semillas e instalación de plántulas debido a las mayores amplitudes térmicas, de humedad y de radiación solar [Citation11–14], disminuyendo estas amplitudes en sitios ubicados bajo protecciones dada por el relieve accidentado [Citation5,Citation6] o por plantas del sotobosque [Citation15]. En consecuencia, la importancia de la protección contra la pérdida de agua se ha enfatizado para el éxito de la germinación y establecimiento de las plántulas, ya que la sequía puede ser la principal causa de mortalidad durante el primer año, después de la germinación en la sucesión primaria [Citation11]. Las grandes rocas aumentan el contenido de humedad del suelo en su entorno inmediato y la temperatura del aire justo por encima del suelo se mantiene más cálida [Citation16–19], generando un micrositio adecuado para la instalación de plantas [Citation11], demostrando que la asociación con grandes rocas son clave en la dinámica de los ecosistemas alpinos [Citation20]. También algunas interacciones entre plantas, conocidas éstas como facilitadoras o plantas nodrizas, crean micrositios favorables para la germinación, el establecimiento y la supervivencia de otras plantas [Citation21–23].
Para la persistencia y dispersión de los bosques, es fundamental una adecuada densidad de renovales, la cual puede estar limitada por cambios en la temperatura y la disponibilidad de agua a lo largo de gradientes de altitud, pendiente y humedad del suelo [Citation12,Citation24]. Estos gradientes topográficos influyen en la variación a escala local en la disponibilidad de nutrientes del suelo, la escorrentía, la infiltración y el microclima del suelo [Citation25], modificando el establecimiento, crecimiento y desarrollo de especies vegetales [Citation26] y determinando la estructura forestal [Citation27,Citation28]. A su vez, la competencia por el agua con la vegetación herbácea tiende a ser un factor de estrés para el establecimiento de renovales en ecosistemas de escasos recursos [Citation29,Citation30].
Gran parte de los ecosistemas alpinos se encuentran dominados por una comunidad caracterizada por pastos perennes y arbustos bajos dispersos [Citation31]. Sin embargo, es poco lo que se sabe sobre qué condiciones de micrositios facilitan el establecimiento de plántulas, siendo particularmente importante para evaluar el potencial de expansión de sus diferentes componentes vegetales [Citation32].
Los ecosistemas alpinos ubicados en los Andes sudamericanos, consisten en una matriz de pastizales y matorrales, con la presencia de un género arborescente, Polylepis (Rosaceae: Sanguisorbeae) [Citation33,Citation34]. Este género domina la canopia de bosques y arbustales de montaña desde la cordillera de Mérida en Venezuela hasta las serranías del centro argentino [Citation35–37] ocupando una gran variedad de hábitats. Su distribución puede ser continua o formar parches aislados monoespecificos [Citation38] y distribuidos en laderas de montañas y roquedales [Citation39]. Dentro de la Argentina hay identificadas cinco especies de Polylepis [Citation36,Citation37], de las cuales Polylepis tarapacana Phill. (comúnmente llamada Quewiña o Queñoa de altura) forma bosques dispersos a la mayor altitud ((3.900-) 4.400–5.000 m snm [Citation37,Citation40]. Polylepis tarapacana se adapta a condiciones extremas de sequía y bajas temperaturas, siendo su crecimiento muy sensible a los cambios hidroclimáticos [Citation32,Citation41–43]. En estas formaciones forestales se pueden caracterizar varios micrositios originados por la heterogeneidad del suelo, el relieve accidentado, la presencia de flora (gramíneas y dicotiledóneas), la acumulación de hojarasca y el suelo mineral expuesto por disturbio, siendo estos sectores del suelo lugares posibles de instalación debido al conjunto de condiciones ambientales [Citation1,Citation17,Citation44,Citation45].
El objetivo de este trabajo fue analizar la facilitación dada por el ambiente y los micrositios en la instalación de renovales de P. tarapacana en la provincia fitogeográfica Altoandina. Planteamos la hipótesis de que la mayor densidad de renovales de P. tarapacana corresponde con micrositios protegidos y con suficiente retención hídrica.
Si esto se cumple, esperamos que i) existirá un óptimo en la altitud, exposición o pendiente donde la densidad de los renovales es máxima [Citation46]; ii) un incremento en la densidad de los bosques producirá un incremento en la densidad de renovales [Citation5,Citation6], iii) micrositios que presentan una mayor retención de humedad (aquellos que se encuentran bajo la protección o cercanía de rocas) son aquellos que presentarán una mayor densidad de renovales, y iv) la mayor abundancia de renovales ocurrirá en zonas que presenten la mayor superficie de micrositios propicios para la retención de humedad.
Materiales y métodos
Área de estudio
Se trabajó con bosques de P. tarapacana distribuidos en la zona de la localidad de Coranzulí (23°00ʹ49ʹ’S, 66°21ʹ59ʹ’O), departamento de Susques, Provincia de Jujuy, Argentina (). El área de estudio se ubica dentro de la Provincia Altoandina, Distrito Quichua [Citation47–49], en las altas cumbres de la cordillera de los Andes por encima de los 4300 m snm. Se desarrollan en regiones con 100 a 500 mm de precipitación media anual, y con 13º C de temperatura media en el mes más cálido (enero) y 3°C de temperatura media del mes más frio [Citation49–53].
Figure 1. Distribución de los bosques de P. tarapacana y ubicación de los sitios de muestreo. A: Distribución de Polylepis tarapacana en los Andes sudamericanos (adaptado de [Citation49]) B: Distribución de P. tarapacana en la Provincia de Jujuy, Argentina (adaptado de [Citation49]) C: Sitios de muestreo
![Figure 1. Distribución de los bosques de P. tarapacana y ubicación de los sitios de muestreo. A: Distribución de Polylepis tarapacana en los Andes sudamericanos (adaptado de [Citation49]) B: Distribución de P. tarapacana en la Provincia de Jujuy, Argentina (adaptado de [Citation49]) C: Sitios de muestreo](/cms/asset/856105e3-f38f-4901-b104-8efa20cc5f47/tneo_a_1902251_f0001_oc.jpg)
Medición de variables topográficas
Se seleccionaron 21 bosques que se desarrollan en un gradiente de exposición, de altitud, pendiente y cobertura forestal. El muestreo se desarrolló en un amplio gradiente de variables topográficas y de estructura forestal ().
Table 1. Rangos de las variables altitud, exposición y pendiente en bosques de P. tarapacana en el norte de Jujuy. Altitud: Altitud en m snm; Exp: Exposición de los sitios de muestreo en grados, ExE: Exposición Este (seno de la exposición en grados, oscilan entre −1 (en el oeste) a 1 (en el este); ExN: Exposición Norte (coseno de la exposición en grados, −1 (en el sur) a 1 (en el norte); Pend: Pendiente en grados. Desvest: Desvío Estándar
Con el fin de conocer el óptimo en las variables ambientales, donde la instalación de los renovales es máxima, se utilizó la densidad de renovales por hectárea relacionando estos valores con la altitud, exposición y la pendiente. En todas las parcelas se utilizó un dispositivo de geoposicionamiento global (GPS) para la medición de la altitud, la pendiente se midió utilizando clinómetro y la exposición mediante brújula. La exposición suele medirse en grados presentando valores entre 0 y 360°, la exposición Norte varía entre 315 y 45º (incluyendo al 0), mientras que la exposición sur entre 135 y 215°. Si se promedian los valores por exposición, los promedios presentaran valores similares a 180º en ambas exposiciones. Para evitar estos errores la exposición en grados se convierte mediante la función seno y coseno en dos variables: norte-sur (ExN) y este-oeste (ExE). Los valores del seno oscilan entre −1 (al oeste) y 1 (al este), mientras que los valores del coseno oscilan entre −1 (al sur) y 1 (al norte).
Estructura forestal
Dentro de cada sector de bosque se seleccionó un punto ubicado al azar, el que fue el inicio de cada unidad de muestreo compuesta por tres subunidades rectangulares de superficie variable. Cada punto se ubicó en el terreno utilizando GPS. Cada subunidad presenta dimensiones particulares (en ancho y largo) que se ajustó a un total mínimo de 20 individuos de cada clase de tamaño: 1) árboles – A (definidos como aquellos individuos iguales o mayores a 1.3 m de altura), 2) arbustos – Ar (individuos entre 20 cm y menores a 1.3 m) y 3) renovales – R (menores a 0.2 m) de P. tarapacana. Las superficies de las parcelas se corrigieron por la pendiente del terreno mediante la siguiente formula: Superficie corregida = Superficie × coseno (pendiente en grados) [Citation54].
Se registraron variables de estructura forestal en árboles, arbustos y renovales. A todos los individuos de las parcelas se le midió el diámetro en la base (DAB – cm), la altura (AT – cm) correspondiente al tronco más alto del árbol y se calculó la altura media dominante (AMD – m), con el promedio de los tres arboles más altos de cada parcela () [Citation55], la densidad como número de árboles por hectárea y el área basal (AB – m2.ha−1) como la sumatoria de las áreas de los troncos expresadas por hectárea. Se definió como micrositio a un área de 10 × 10 cm centrada en cada renoval (R) presente en la parcela, y se caracterizó utilizando los siguientes parámetros: 1) Monocotiledóneas-MONO (especies de la familia Poaceae); 2) Dicotiledóneas-DICO (incluyendo ejemplares de P. tarapacana y otras dicotiledóneas del sotobosque); 3) Hojarasca-H (residuos vegetales en superficie); 4) Suelo desnudo-SD (suelo abierto (orgánico o mineral), superficies terrosas o arenosas que no están cubiertas por vegetación); 5) Roca-R (como aquellos afloramientos rocosos que se hallan fijados al suelo); 6) Glera-G (piedras sueltas de diversos tamaños depositadas en superficie o semifijas al suelo; su grano siempre es mayor que el de la fracción arena); 7) Borde de roca-BR (zona de influencia entre las rocas y el suelo o glera circundante) y 8) Suelo bajo protección de roca-SBR (zona cubierto por rocas generando un ambiente sombrío), quedando así 2 categorías de micrositios caracterizados por tipos de plantas y 6 por sustratos. Asimismo, a cada renoval presente en la parcela se le midió la distancia comprendida entre el ápice y el tercer nudo, utilizando como indicador del crecimiento anual en altura el promedio de las 3 distancias [Citation56].
Figure 2. Determinación de la altura total, y diámetro en la base. AT: Altura total, DAB: diámetro en la base
Caracterización de micrositios
Se caracterizó la variabilidad de micrositios presentes en cada uno de los 21 sitios de muestreo, mediante el cálculo de la abundancia porcentual de las ocho categorías de micrositios. Esta determinación se basó en la metodología propuesta por Martínez Pastur et al. [Citation57] y Toro Manríquez et al. [Citation2] en bosques de Nothofagus. En cada una de las parcelas se relevó la abundancia porcentual, mediante 100 parcelas contiguas de 1 m2 cada una, obteniendo así el porcentaje de cada categoría de micrositio en cada parcela. De esta forma este porcentaje se puede relacionar con las variables topográficas y de estructura forestal de cada parcela. Se extrajo una muestra de suelo de por lo menos 100 g de peso a 2 cm de profundidad de cada micrositio (MONO, DICO, H, SD, G, BR y SBR) por parcela. A cada muestra en laboratorio, se le determinó la humedad relativa (HR) mediante secado en estufa a 70°C hasta peso constante.
Análisis estadísticos
Se realizó un análisis de correlación de Pearson entre variables topográficas (Altitud, Exposición Este, Exposición Norte y Pendiente) y de estructura forestal de renovales y de árboles y arbustos. Las variables de estructura forestal analizadas fueron la densidad (ind.ha−1), diámetro en la base (DAB – cm), área basal (AB – m2.ha−1) y, sólo para renovales, el crecimiento en altura (utilizando como indicador el largo promedio de entrenudo en cm.año−1). Asimismo, se realizó un ANOVA simple utilizando las 8 categorías de micrositios como factor principal para analizar la humedad del suelo y la densidad de renovales. Las diferencias de crecimiento y densidad bajo diferentes micrositios se determinaron mediante comparaciones de medias adecuadas al tipo de datos (p < 0.05). En todos los casos se realizaron los test de normalidad mediante la prueba de Kolmogorov-Smirnov, mientras que la homogeneidad de variancias (homocedasticidad) se analizó mediante la prueba de Levene. En caso de incumplimiento de supuestos se aplicaron métodos no paramétricos (Kruskal-Wallis). El porcentaje de ocupación de las 8 categorías microambientales en cada sitio se relacionó con la densidad de renovales de P. tarapacana.
Resultados
Las variables de estructura forestal () presentaron una amplia variación, donde las variables de estructura de renovales indican una baja ocupación en contraste con la estructura de árboles y arbustos (la densidad de renovales representa el 36.8% de la densidad de árboles y arbustos).
Table 2. Rangos de las variables de estructura forestal en renovales y árboles y arbustos para P. tarapacana en la Provincia de Jujuy. Dens: Densidad en ind.ha−1; R: renovales (plantas < 20 cm de altura); Crec: crecimiento de renovales utilizando como indicador el largo promedio de entrenudo (cm); DAB: diámetro en la base (cm); AB: Área basal en m2.ha−1; A+ Ar: árboles y arbustos (plantas ≥ 20 cm de altura); AMD: Altura media dominante como promedio de los 3 árboles más altos (cm); Desvest: Desvío Estándar
El análisis de correlación entre variables ambientales (Altitud, exposición Este (ExE), exposición Norte (ExN) y Pendiente) y estructura forestal de árboles y arbustos (A+ Ar) mostró que a medida que la pendiente del terreno se incrementa, los bosques presentan individuos de menor tamaño (Pendiente y diámetro a la base, coeficiente de correlación CC = −0.5337; p = 0.0127; Pendiente y AMD (altura media dominante) (CC = −0.5030; p = 0.0201). Esto mismo se observó en la densidad de renovales (CC = −0.5550; p = 0.0090), en donde para mayores pendientes la presencia de renovales disminuye (). Sin embargo, no se hallaron diferencias significativas entre densidades de renovales debido al incremento de la altitud (p = 0.15). En cuanto a la estructura forestal, se observó que la densidad de renovales aumenta cuando los árboles que forman el bosque presentan mayores diámetros basales (CC = 0.6385; p = 0.0018) y mayor altura media dominante (AMD) (CC = 0.7023; p = 0.0004). Esta correlación también se observó en el área basal de renovales y el diámetro a la base (DAB) de árboles del bosque (CC = 0.5557; p = 0.0089) y la AMD (CC 0.5982; p = 0.0042) ().
Table 3. Análisis de correlación de Pearson entre variables de estructura forestal de renovales y variables topográficas y de estructura forestal en bosques de P. tarapacana. ExE: Seno de la exposición en grados, presentando valores de −1 (al oeste) y 1 (al este), ExN: Coseno de la exposición en grados, presentando valores de −1 (al sur) y 1 (al norte), Altitud: Altitud en m s.n.m., Pendiente en grados, DAB: diámetro en la base (cm), A+ Ar: árboles y arbustos (plantas ≥ 20 cm de altura); AMD: Altura media dominante como promedio de los 3 árboles más altos (cm), AB: Área basal en m2.ha−1, Dens: Densidad de renovales en ind.ha−1, Crec: Crecimiento de renovales utilizando como indicador el largo promedio de entrenudo en cm, valores en coeficiente de correlación (p-valor)
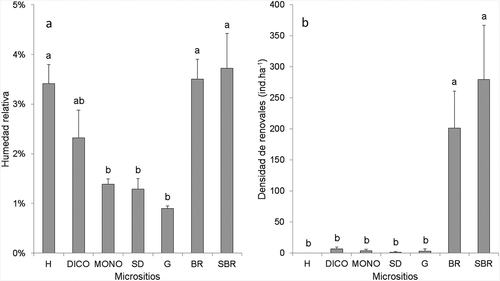
La humedad relativa del suelo varió de acuerdo al micrositio (p = 0.0000), encontrando mayores porcentajes en los micrositios bajo la influencia de rocas, y con alta cobertura. El Borde de roca (BR) y el Suelo bajo protección de roca (SBR) y Hojarasca (H) presentaron valores de 3.5%, 3.7% y 3.4% respectivamente (), y los menores porcentajes de humedad relativa (HR) fueron hallados sitios más expuestos correspondientes a Dicotiledóneas (DICO) (2.3%), Monocotiledóneas (MONO) (1.4%); Suelo desnudo (SD) (1.3%) y Glera (G) (0.9%). La densidad de plántulas de P. tarapacana varió significativamente entre micrositios (p = 0.0000), mostrando un patrón similar a lo observado en el análisis de humedad de suelo. La mayor densidad de renovales se encontró en los micrositios BR y SBR (201.2 y 279.5 ind.ha−1 respectivamente) (), mientras que en el resto de los micrositios presentaron una escasa cantidad de plántulas (6.5 en DICO y 0.0 ind.ha−1 en H).
Figure 3. a) Humedad relativa del suelo en porcentaje hallada en cada uno de los 8 tipos de micrositios en bosques de P. tarapacana en la Provincia de Jujuy b) Densidad de renovales (ind.ha−1) en cada uno de los 8 micrositios en bosques de P. tarapacana en la Provincia de Jujuy. MONO: Monocotiledóneas; DICO: Dicotiledóneas; H: Hojarasca; SD: Suelo desnudo; R: Roca. G: Glera; BR Borde de roca; SBR: Suelo bajo protección de roca; barras de error en desvío estándar, letras diferentes indican diferencias significativas según test de Tuckey
Existe una oferta variada de micrositios posibles de encontrar en la zona de estudio (), siendo Glera (G), Suelo desnudo (SD), BR y Roca (R) los que presentaron los mayores porcentajes (entre 18,2 y 16.1%). La abundancia porcentual de los restantes micrositios disminuye siendo MONO y H los micrositios que presentaron los menores promedios (5.9 y 5.7%) y menores desvíos (3.2 y 2.7).
Table 4. Rangos de abundancia de micrositios en porcentaje presentes en bosques de P. tarapacana en los sitios de muestreo. MONO: Monocotiledóneas; DICO: Dicotiledóneas; H: Hojarasca; SD: Suelo desnudo; R: Roca. G: Glera; BR Borde de roca; SBR: Suelo bajo protección de roca; valores en porcentaje; Desvest: Desvío Estándar
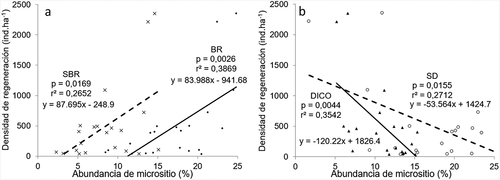
Se observó que la densidad de los renovales varía con la oferta de alguno de los micrositios presentes. Vemos que existe una tendencia positiva (), en donde a mayor oferta de los micrositios Borde de roca (BR) y Suelo bajo protección de roca (SBR) mayor es la densidad de los renovales (BR: coeficiente de correlación: 0.6220; p = 0.0026, SBR: 0.5150 (0.0169)) y una tendencia negativa () donde a mayor oferta de los micrositios Dicotiledóneas (DICO) y Suelo desnudo (SD), menor es la densidad de renovales encontrada (DICO −0.5952 (0.0044); SD −0.5208 (0.0155)).
Figure 4. Abundancia en porcentaje de los micrositios (a) Suelo bajo protección de roca (SBR) y borde de roca (BR) y (b) sotobosque de Dicotiledóneas (DICO) y suelo desnudo (SD) en relación a la densidad (ind.ha−1) de renovales en bosques de P. tarapacana en la Provincia de Jujuy, Argentina. C: coeficiente de correlación, p: probabilidad
Discusión
Con el fin de encontrar un óptimo en las variables topográficas (altitud, exposición o pendiente) donde la instalación de los renovales es máxima, es preciso que los gradientes ambientales analizados, sean los más amplios posibles. Los bosques estudiados abarcaron un amplio gradiente de exposición, presentándose en exposiciones de W a E, mientras que en el gradiente N – S se encontraron sólo en laderas orientadas al Norte. La distribución de los bosques de P. tarapacana se encuentra relacionada con la humedad en una escala geográfica amplia (a lo largo de la Cordillera Occidental), mientras que a escala local su distribución se encuentra influida por la irradiación solar, con árboles claramente restringidos a las exposiciones más cálidas [Citation58]). Las poblaciones de P. tarapacana en el norte de Chile, se encuentran distribuidas preferentemente en exposiciones oeste y noroeste [Citation59,Citation60], siendo también lo observado por Hoch y Körner [Citation56] y Braun [Citation58] donde P. tarapacana, en el volcán Sajama, se desarrolla en altitudes más altas en laderas cálidas y secas (N), disminuyendo progresivamente hacia las laderas del sur, presentando más humedad pero al menos 6 grados más frías con respecto a las exposiciones orientadas al norte. Piña [Citation61] describe formaciones en todo el gradiente de exposiciones para la especie, coincidiendo en parte con los resultados hallados en el presente trabajo. Dado que no observamos bosques de P. tarapacana en laderas sur en el sitio de estudio, sería interesante realizar una búsqueda con una metodología más robusta y a nivel regional para determinar si es un fenómeno local o una característica de la especie.
En cuanto a las pendientes analizadas, contemplan una gran amplitud de situaciones, coincidiendo esto con lo observado en otros sitios de desarrollo de P. tarapacana [Citation62]. Las pendientes donde se desarrolla P. tarapacana se consideran moderada a pronunciada (15 a 20°) [Citation59], mientras que las observadas en este trabajo corresponden a un gradiente mucho mayor, comprendiendo pendientes desde un terreno prácticamente plano (1°) a pendientes abruptas (37°). Los bosques de Polylepis tarapacana analizados en este trabajo, presentaron un gradiente coincidente con el gradiente de altitud estudiado por diversos autores [Citation37,Citation40,Citation59].
Se observó una relación inversa de la pendiente con la densidad de renovales y el tamaño de árboles y arbustos (A+ Ar), registrando baja densidad de renovales y plantas pequeñas en pendientes pronunciadas. En estudios similares se encontraron resultados opuestos, en donde se observó la mayor incorporación de renovales en laderas más pronunciadas, donde existe una mayor disponibilidad de humedad [Citation59]. En bosques de P. tarapacana del volcán Sajama, Hoch y Körner [Citation56] observaron una tendencia similar a los resultados del presente trabajo, donde la densidad de renovales (5 a 50 cm de altura) no presentó diferencias significativas entre las 3 altitudes analizadas (400 ind.ha−1 a los 4360 m snm; 800 ind.ha−1 en 4550 m snm y 500 ind.ha−1 a 4810 m snm). Además, estos autores encontraron numerosas plántulas pequeñas (< a 5 cm de altura) a 4810 m snm pero no en las elevaciones más bajas. La hipótesis planteada -existirá un óptimo en la altitud, exposición o pendiente donde la instalación de los renovales es máxima- se cumple en forma parcial, dado que la instalación de renovales es máxima en bajas pendientes, al no presentar una tendencia significativa la altitud y la exposición.
La estructura forestal presentó una amplia variación en densidad, y altura de los árboles, similar a otros estudios de estructura de bosques de P. tarapacana [Citation59,Citation62]. La correlación positiva entre tamaño de árboles (diámetro en la base y la altura) y la densidad de renovales, implica que, en bosques con árboles más grandes y frondosos, mayor será la densidad de renovales debido a un mayor aporte de semillas [Citation63]. En este estudio la densidad de renovales no se relacionó con la mayor densidad, pero sí con el tamaño de árboles y arbustos. Resultados similares fueron encontrados por Hertel y Wesche [Citation64] en bosques de altura de Polylepis pepei B.B. Simpson y Polylepis besseri Hieron. donde la mayor densidad de renovales se presentó en bosques compuestos por arboles de mayor porte, mientras que la incorporación fue prácticamente nula en sitios donde se observaron árboles de menor tamaño. En base a esto, se rechaza la hipótesis de que un incremento en la densidad de los bosques producirá un incremento en la densidad de renovales, debido a que este incremento no se relaciona con la densidad del bosque, sino con el tamaño de los individuos que lo componen.
Existen asociaciones positivas entre la mayor densidad de renovales de P. tarapacana y los micrositios bajo la influencia de rocas, las cuales podrían explicarse por la mayor concentración humedad en el suelo adyacente o debajo de grandes rocas y en depresiones superficiales, lo que mejora la germinación, crecimiento y supervivencia de semillas, particularmente en entornos donde la desecación de la superficie puede ser un factor importante [Citation11]. El mayor contenido de humedad del suelo, en este estudio, fue encontrado en el entorno inmediato de grandes rocas (micrositios Borde de roca – BR y Suelo bajo protección de roca – SBR) debido, entre otros factores, a que en esos micrositios se concentra el agua proveniente de la precipitación [Citation11]. Estos resultados se explican por el hecho de que las grandes rocas también proporcionan sombra durante el día, reduciendo la evaporación del suelo y fluctuaciones de temperatura más pequeñas [Citation11,Citation19].
Un hallazgo interesante es el alto porcentaje de humedad relativa en el micrositio Hojarasca (H), similar a lo observado en Borde de roca y Suelo bajo protección de roca. Sin embargo, en contraste a lo ocurrido en estos micrositios bajo la influencia de rocas, en H no se encontraron renovales de P. tarapacana. Algunos autores afirman que una capa gruesa de hojarasca perjudica la incorporación de renovales, debido a la mayor distancia entre la semilla y el suelo mineral y a la radiación solar reducida si las semillas están cubiertas por la hojarasca [Citation13,Citation65,Citation66]. En el presente estudio, la hojarasca observada podría impedir la incorporación de renovales, pero ser beneficiosa para el ejemplar adulto ya que funcionaría como acumulador de la humedad y aislante de las temperaturas extremas de la superficie [Citation13].
Para evaluar el crecimiento en altura de los renovales, se utilizó la técnica de estimación de la edad mediante nudos descrita Hoch y Körner [Citation56], que demostró tener potencialidad de uso para P. tarapacana [Citation67]. La distancia comprendida entre el ápice y el tercer nudo del renoval no mostró tener relación con los diferentes micrositios, implicando que los diferentes micrositios presentan diferencias para los primeros años de instalación de la plántula (emergencia de radícula, desarrollo de raíz principal y de primeras hojas verdaderas) siendo ésta la etapa más vulnerable, y no el posterior crecimiento en altura del renoval [Citation11,Citation13,Citation56].
Para la expansión de la cubierta forestal, es importante comprender la capacidad de las plántulas para colonizar, sobrevivir y crecer en estas áreas e identificar importantes barreras bióticas y abióticas [Citation68]. Hoch y Körner [Citation56] señalan que existe, para estas poblaciones, una fase crítica para el establecimiento de árboles de P. tarapacana, siendo más importante la protección dada por rocas o plantas en la etapa de germinación que en la supervivencia de las plántulas. Sin embargo, en el presente trabajo los renovales fueron encontrados casi exclusivamente en Borde de roca (BR) y Suelo bajo protección de roca (SBR), coincidiendo con los resultados obtenidos por Saavedra [Citation59] en donde el 75% de la regeneración se encuentra protegida por piedras, lo que indica que es probable que la germinación no sea un factor limitante y que las condiciones para la supervivencia y el crecimiento de las plantas sean restrictivas [Citation11,Citation69]. En poblaciones de P. tomentella, el reclutamiento de plántulas constituye un cuello de botella en la regeneración de poblaciones fuertemente perturbadas [Citation70]. En estas poblaciones, las plantas reproductivas pueden producir grandes cantidades de frutos, pero un mayor rendimiento reproductivo no se traduce en mayores tasas de reclutamiento de plántulas. De esta forma, se acepta, en el caso de BR y SBR, la hipótesis de que micrositios que presentan una mayor retención de humedad son aquellos que presentarán una mayor densidad de renovales, mientras que Hojarasca (H) no es un micrositio óptimo para la instalación de renovales. Esto indicaría la importancia de la oferta de micrositios que proporcionen protección durante el crecimiento y desarrollo de las plántulas en estos ambientes extremos como la Puna [Citation71].
Observamos una variada oferta de micrositios en la zona de estudio, siendo la Glera (G), Suelo desnudo (SD) y Borde de roca (BR) los micrositios que presentaron mayor superficie. A su vez, la densidad de los renovales varió con esta oferta de micrositios, ya que el establecimiento y la supervivencia exitosa de plántulas ocurren en sitios seguros y que brindan las condiciones necesarias para el desarrollo de los renovales [Citation5,Citation11]. Como describe Jumpponen [Citation11], la presencia de plántulas depende menos de la topografía en relación con el entorno, que de los parámetros que describen a los micrositios en sí. Por lo tanto, se espera que la distribución de plántulas, en los bosques alpinos, esté determinada por la oferta de micrositio, que gobiernen la captura de semillas, la germinación, protección y el establecimiento exitoso. La hipótesis planteada -la mayor abundancia de renovales ocurrirá en sitios que presenten la mayor superficie de micrositios propicios para la retención de humedad- se acepta parcialmente, debido a que la oferta de micrositio H no influye sobre la densidad de renovales, y sí BR y SBR que influyen positivamente. Debido a la importancia de la facilitación por micrositios como Borde de roca y Suelo bajo protección de roca en áreas desérticas, sería una consideración importante para la restauración de bosques de P. tarapacana [Citation19]. Estos antecedentes podrían generar nuevos conocimientos para el desarrollo de métodos de restauración a partir de una comprensión más profunda de la dinámica de regeneración natural [Citation70].
Estos resultados permiten definir los micrositios propicios con fines de restauración y conservación. La identificación de los micrositios aptos para la regeneración de P. tarapacana puede servir como antecedente para identificar las áreas que podrían recuperarse de forma natural o los lugares donde mejor se podría incorporar una planta por medio de la restauración activa. Es necesario continuar con estudios sobre la estructura de estos bosques y su relación con características ambientales, sobre todo del suelo donde se desarrollan estas formaciones, dado que la bibliografía es escasa al respecto. Asimismo, sugerimos realizar estudios que abarquen distintos gradientes de influencias abióticas o topográficas que contribuyan al entendimiento de la ecología de estos bosques y generen aportes sólidos a su conservación y restauración.
Agradecimientos
A la comunidad de Coranzulí, que nos permitió ingresar en estos bosques para poder investigar la forma de conservarlos. A José Hector Siares y Nirma Martina Lamas directora de la escuela ESC N°362 Héroes de Malvinas por la ayuda y el apoyo en este trabajo, a la Ing. Agr. Griselda Beatriz Maizarez por su trabajo en el campo y a los/las revisores/as anónimos/as por sus acertados comentarios que han mejorado sustancialmente el manuscrito.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Christie DA, Armesto J. Regeneration microsites and tree species coexistence in temperate rain forest of Chiloé Island, Chile. J Ecol. 2003;91(5):776–784.
- Toro Manríquez M, Promis A, Huertas Herrera A, et al. Influencia del micrositio y la exposición en la regeneración de bosques de Nothofagus pumilio afectados por Castor canadensis en Tierra del Fuego: un análisis exploratorio [Influence of the microsite and exposure on the regeneration of Nothofagus pumilio forests affected by Castor canadensis in Tierra del Fuego: an exploratory analysis]. Bosque. 2018;39(3):431–440. Spanish.
- Dalling JW, Hubbell SP. Seed size, growth rate and gap microsite conditions as determinants of recruitment success for pioneer species. J Ecol. 2002;90(3):557–569.
- Titus J, Nowak R, Smith S. Soil resource heterogeneity in the Mojave Desert. J Arid Environ. 2002;52(3):269–292.
- Dobrowski SZ, Swanson AK, Abatzoglou JT, et al. Forest structure and species traits mediate projected recruitment declines in western US tree species. Glob Ecol Biogeogr. 2015;24(8):917–927. .
- Körner C. Alpine treelines: functional ecology of the global high elevation tree limits. Basel: Springer; 2012.
- Cabrera AL. Ecología vegetal de la Puna [Plant ecology of the Puna]. Colloq Geogr. 1968;9: 91–116. Spanish.
- Brack Egg A, Mendiola C. Ecologı́a del Perú [Ecology of Peru]. Lima: Quebecor world Peru; 2004. ( Spanish).
- Monteiro JAF, Hiltbrunner E, Körner C. Functional morphology and microclimate of Festuca orthophylla, the dominant tall tussock grass in the Andean Altiplano. Flora. 2011;206(4):387–396.
- Asefa M, Cao M, Zhang G, et al. Environmental filtering structures tree functional traits combination and lineages across space in tropical tree assemblages. Sci Rep. 2017;7(1):132. .
- Jumpponen A, Vare H, Mattson KG, et al. Characterization of ‘safe sites’ for pioneers in primary succession on recently deglaciated terrain. J EcolN. 1999;87(1):98–105. .
- Bader MY, Van Geloof I, Rietkerk R. High solar radiation hinders tree regeneration above the alpine treeline in northern Ecuador. Plant Ecol. 2007;191(1):33–45.
- Cierjacks A, Iglesias JE, Wesche K, et al. Impact of sowing, canopy cover and litter on seedling dynamics of two Polylepis species at upper tree lines in central Ecuador. J Trop Ecol. 2007;23(3):309–318. .
- Rehm EMK, Feeley J. Forest patches and the upward migration of timberline in the southern Peruvian Andes. For Ecol Manage. 2013;305:204–211.
- Acuña-Rodriguez IS, Cavieres LA, Gianoli E. Nurse effect in seedling establishment: facilitation and tolerance to damage in the Andes of central Chile. Rev Chil Hist Natural. 2006;79:329–336.
- Parker KC. Site-related demographic patterns of organ-pipe cactus populatlons in southern Arizona. Bull Torrey Bot Club. 1987;114(2):149–155.
- Perez FL. Particle sorting due to off-road vehicle traffic in a high Andean paramo. Catena. 1991;18(3–4):239–254.
- Kleier C, Rundel PW. Microsite requirements, population structure and growth of the cushion plant Azorella compacta in the tropical Chilean Andes. Aust Ecol. 2004;29(4):461–470.
- Kleier C, Lambrinos JG. The importance of nurse associations for three tropical alpine life forms. Arctic, Antarctic, and Alpine Research. 2005;37(3):331–336.
- Callaway RM, Brooker RW, Choler P, et al. Positive interactions among alpine plants increase with stress. Nature. 2002;417(6891):844–848. .
- Valiente-Banuet A, Vite F, Zavala-Hurtado JA. Interaction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana. J Veg Sci. 1991;2(1):11–14.
- Tewksbury JJ, Lloyd JD. Positive interactions under nurse plants: spatial scale, stress gradients and benefactor size. Oecologia. 2001;127(3):425–434.
- Catorci A, Cesaretti S, Velasquez JL, et al. Plant–plant spatial interactions in the dry Puna (southern Peruvian Andes). Alp Bot. 2011;121(2):113–121. .
- Johnson D, McCulloh K, Reinhardt K. The earliest stages of tree growth: development, physiology and impacts on microclimate. In: Meinzer FC, Lachenbruch B, Dawson TE, editors. Size- and Age-Related Changes in Tree Structure and Function. New York (NY): Springer; 2011. p. 65–87.
- Yirdaw E, Starr M, Negash M, et al. Influence of topographic aspect on floristic diversity, structure and treeline of afromontane cloud forests in the Bale Mountains, Ethiopia. J For Res. 2015;26(4):919–931. .
- Kraft NJB, Adler PB, Godoy O, et al. Community assembly, coexistence, and the environmental filtering metaphor. Funct Ecol. 2015;29(5):592–599. .
- Gallardo-Cruz JA, Pérez-García EA, Meave JA. β-Diversity and vegetation structure as influenced by slope aspect and altitude in a seasonally dry tropical landscape. Landsc Ecol. 2009;24(4):473–482.
- Werner FA, Homeier J. Is tropical montane forest heterogeneity promoted by a resource-driven feedback cycle? Evidence from nutrient relations, herbivory and litter decomposition along a topographical gradient. Funct Ecol. 2015;29(3):430–440.
- Nambiar EKS, Sands R. Competition for water and nutrients in forests. Can J For Res. 1999;23(10):1955–1968.
- Balandier P, Collet C, Miller JH, et al. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry. 2006;79(1):3–27. .
- Gajardo R. La Vegetación Natural de Chile: clasificación y Distribución Geografica [The Natural Vegetation of Chile: geographical Classification and Distribution]. Santiago: Editorial Universitaria; 1994. ( Spanish).
- Morales MS, Villalba R, Grau HR, et al. Rainfall controlled tree growth in high elevation subtropical treelines. Ecology. 2004;85(11):3080–3089. .
- Kessler M, Herzog SK. Conservation status in Bolivia of timberline habitats, elfin forest and their birds. Cotinga. 1998;10:50–54.
- Jácome J, Kessler M, Smith AR. A human-induced downward-skewed elevational abundance distribution of pteridophytes in the Bolivian Andes. Glob Ecol Biogeogr. 2007;16(3):313–318.
- Simpson BB. A revision of the Genus Polylepis (Rosaceae: sanguisorbeae). Smithson Contrib Bot. 1979;43(43):1–62.
- Kessler M, Schmidt-Lebuhn AN. Taxonomical and distributional notes on Polylepis (Rosaceae). Org Diver Evol. 2006;5:1–10.
- Renison D, Cuyckens GAE, Pacheco S, et al. Distribución y estado de conservación de las poblaciones de árboles y arbustos del género Polylepis (Rosaceae) en las montañas de Argentina [Distribution and conservation status of populations of trees and shrubs of the genus Polylepis (Rosaceae) in the mountains of Argentina]. Ecol Austral. 2013;23(1):27–36. Spanish.
- Zutta BR, Phillip WR, Saatchi S, et al. Prediciendo la distribución de Polylepis: bosques Andinos vulnerables y cada vez más importantes [Predicting the distribution of Polylepis: vulnerable and increasingly important Andean forests]. Rev Peru Biol. 2012;19(2):205–212. Spanish. .
- Kessler M. The genus Polylepis (Rosaceae) in Bolivia. Candollea. 1995;50:131–171.
- Kessler M. Bosques de Polylepis [Polylepis forests]. In: Moraes M, Øllgaard B, Kvist LP, et al. editors. Botánica Económica de los Andes Centrales [Economic Botany of the Central Andes]. La Paz: Universidad Mayor de San Andrés, 2006:110–120. Spanish.
- Morales MS, Christie D, Neukom R, et al. Variabilidad hidroclimática en el sur del altiplano: pasado, presente y futuro [Hydroclimatic variability in the southern highlands: past, present and future]. Tucumán, Argentina: Serie Conservación de la Naturaleza, Fundación Miguel Lillo; 2018. (Grau HR, Babot J, Izquierdo A, et al. editors. La Puna Argentina: naturaleza y cultura; vol. 24). Spanish.
- Christie DA, Lara A, Barichivich J, et al. El Niño-Southern Oscillation signal in the world’s highest-elevation tree-ring chronologies from the Altiplano, Central Andes. Palaeogeogr Palaeoclimatol Palaeoecol. 2009;281(3–4):309–319. .
- Solíz C, Villalba R, Argollo J, et al. Spatio-temporal variations in Polylepis tarapacana radial growth across the Bolivian Altiplano during the 20th century. Palaeogeogr Palaeoclimatol Palaeoecol. 2009;281(3–4):296–308. .
- Kuuluvainen T, Kalmari R. Regeneration microsites of Picea abies seedlings in a windthrow area of a boreal oldgrowth forest in southern Finland. Ann Bot Fenn. 2003;40:401–413.
- Bailey T, Davidson N, Close D. Understanding the re-generation niche: microsite attributes and recruitment of eucalypts in dry forests. For Ecol Manage. 2012;269:229–238.
- Tecco PA, Pais-Bosch AI, Funes G, et al. Mountain invasions on the way: are there climatic constraints for the expansion of alien woody species along an elevation gradient in Argentina? J Plant Ecol. 2016;9(4):380–392. .
- Cabrera ÁL. Fitogeografía de la república argentina [Phytogeography of the Argentine Republic]. Bol Soc Argent Bot. 1971;14: 1–42. Spanish.
- Aceituno P. Elementos del clima en el altiplano sudamericano [Elements of the climate in the South American highlands]. Rev Geofis. 1993;44: 37–55. Spanish.
- Cuyckens GAE, Christie DA, Domic AI, et al. Climate change and the distribution and conservation of the world’s highest elevation woodlands in the South American Altiplano. Glob Planet Chang. 2016;137:79–87.
- Renison D, Hensen I, Suárez R, et al. Soil conservation in Polylepis mountain forests of Central Argentina: is livestock reducing our Natural capital? Austral Ecol. 2010;35(4):435–443. .
- Mendoza EA, González JA. Las ecorregiones del noroeste argentino basadas en la clasificación climática de köppen [The ecoregions of the Argentine Northwest based on the Köppen climate classification]. Ser Conserv Nat. 2011;19: 3–41. Spanish.
- Morales MS, Christie DA, Villalba R, et al. Precipitation changes in the South American Altiplano since 1300 AD reconstructed by tree-rings. Clim Past. 2012;8(2):653–666. .
- Wawrzyk AC, Vilá BL. Dinámica de pastoreo en dos comunidades de la puna de Jujuy, Argentina. Lagunillas del Farallón y Suripujio [Grazing dynamics in two communities of the Jujuy puna, Argentina. Lagunillas del Farallón and Suripujio]. Rev Antropol Chil. 2013;45(2): 349–362. Spanish.
- Prodan M, Peters R, Cox F, et al. Mensura forestal [Forest mensuration]. San José, Costa Rica: IICA; 1997. Spanish.
- Assmann E. The principles of forest yield study: studies in the organic production, structure, increment, and yield of forest stands. Oxford: Pergamon; 1970.
- Hoch G, Körner C. Growth, demography and carbon relations of Polylepis trees at the world’s highest treeline. Funct Ecol. 2005;19(6):941–951.
- Martínez Pastur G, Jordán C, Lencinas MV, et al. Landscape and microenvironmental conditions influence over regeneration dynamics in old-growth Nothofagus betuloides Southern Patagonian forests. Plant Biosyst. 2012;146(1):201–213. .
- Braun G. The use of digital methods in assessing forest patterns in an Andean environment: the Polylepis example. Mt Res Dev. 1997;17(3):253–262.
- Saavedra J. Caracterización de la estructura poblacional de Polylepis tarapacana en sector cerro Chiguana, cuenca del salar de Surire, para establecer los lineamientos de un programa de restauración ecológica [Characterization of the population structure of Polylepis tarapacana in the Cerro Chiguana sector, Salar de Surire basin, to establish the guidelines for an ecological restoration program] [master’s thesis]. Arica: Universidad de Tarapacá; 2013. ( Spanish).
- Choque A Evaluación del estado de conservación y propuesta de manejo silvicultural en formaciones naturales de queñoa de altura (Polylepis tarapacana Phil.) en la Reserva Nacional Alto Loa, Región de Antofagasta [Assessment of the state of conservation and proposal of silvicultural management in natural formations of high altitude queñoa (Polylepis tarapacana Phil.) In the Alto Loa National Reserve, Antofagasta Region]. [degree’s thesis].Santiago: Universidad de Chile; 2010. Spanish.
- Piña PZ. Caracterización de hábitat y distribución espacial de formaciones boscosas de queñoa (Polylepis tarapacana) en los territorios del Parque Nacional Salar del Huasco, región de Tarapacá [Habitat characterization and spatial distribution of forest formations of queñoa (Polylepis tarapacana) in the territories of the Salar del Huasco National Park, Tarapacá region]. Biodiversidata. 2015;3: 12–20. Spanish.
- Ríos S Estudio de la distribución y caracterización ecológica de las poblaciones locales de queñoa de altura (Polylepis tarapacana Phil.) en el sector de la provincia de Iquique. Tarapacá. Chile [Study of the distribution and ecological characterization of local populations of highland queñoa (Polylepis tarapacana Phil.) In the sector of the province of Iquique. Tarapac;. Chile] [degree’s thesis].Santiago: Universidad de Chile; 1998. Spanish.
- Domic AI, Mamani E, Camilo G. Fenología reproductiva de la kewiña (Polylepis tomentella, Rosaceae) en la puna semihúmeda de Chuquisaca (Bolivia) [Reproductive phenology of the kewiña (Polylepis tomentella, Rosaceae) in the semi-humid puna of Chuquisaca (Bolivia)]. Ecol Bol. 2013;48(1): 31–45. Spanish.
- Hertel D, Wesche K. Tropical moist Polylepis stands at the treeline in East Bolivia: the effect of elevation on stand microclimate, above- and below-ground structure, and regeneration. Trees. 2008;22(3):303–315.
- Facelli JM, Pickett STA. Plant litter: its dynamics and effects on plant community structure. The Bot Rev. 1991;57(1):1–33.
- Peterson CJ, Facelli JM. Contrasting germination and seedling growth of Betula alleghaniensis and Rhus typhina subjected to various amounts and types of plant litter. Am J Bot. 1992;79(11):1209–1216.
- López VL, Crisci G, Cuyckens GAE, et al. Conteo de marcas de yemas para estimar la edad en Polylepis tarapacana: Una técnica potencial [Bud mark count to estimate age in Polylepis tarapacana: A potential technique]. Ecol austral. 2021;31:182-189. Spanish.
- Domic AI, Capriles JM. Allometry and effects of extreme elevation on growth velocity of the Andean tree Polylepis tarapacana Philippi (Rosaceae). Plant Ecol. 2009;205(2):223–234.
- Hamrick JL, Lee JM. Effect of soil surface topography and litter cover on the germination, survival, and growth of musk thistle (Carduus nutans). Am J Bot. 1987;74(3):451–457.
- Domic AI, Camilo GR, Capriles JM. Small-scale farming and grazing reduce regeneration of polylepis tomentella (rosaceae) in the semiarid andes of bolivia. Biotropica. 2014;46(1):106–113.
- Uriarte M, Bruna EM, Rubim P, et al. Effects of forest fragmentation on the seedling recruitment of a tropical herb: assessing seed vs safe-site limitation. Ecology. 2010;91:1317–1328.