
ABSTRACT
Opportunities to track environmental changes over more than a century are rare in tropical mountains. Edward Whymper’s survey of flora and fauna on the summit of Mt. Corazón (Ecuador, 4788 m a.s.l.) in 1880 provides a unique opportunity to compare historical observations with the current composition of plant and insect communities on a tropical alpine mountain top. We studied Whymper’s archives and historic specimens in London and Paris, and performed a resurvey of vascular plants and ground beetles (Coleoptera Carabidae) in January 2020.
Currently, a large part of the summit area of Corazón is heavily damaged by trampling and stone removal due to mountain tourism, and no vascular plants are present in the deteriorated area on the top of the ridge. However, more species were collected in 2020 than in 1880: 22 of vascular plants vs. 7, and 4 of ground beetles vs. 1. Upslope shifts over 140 years may partly explain this increase in species richness, although the low numbers of Whymper’s sampling may also be due to less skilled collectors and to the presence of permanent snow beds on the summit.
The current faunistic and floristic data presented in this contribution can be used as a baseline for future resurveys of Corazón, in order to monitor changes in the species distribution and community composition of its summit area. Owing to the very small area of its superpáramo and to the soil deterioration by trampling along the summit ridge, Corazón is especially exposed to the effects of climate change and to the risk of extirpation of endemic cold-adapted specialists.
Introduction
Climate change affects species distribution and community composition in montane habitats worldwide [Citation1–4]. Páramo sky islands, a highly diverse alpine habitat unique to the tropical high Andes, are particularly vulnerable to human-induced changes that are expected to have significant consequences on the species richness and ecosystem function [Citation5]. A common response of cold-adapted alpine species to climate change is migration towards higher elevations. Such upward shift has been reported for páramo ground beetles [Citation6] and vascular plants [Citation7], although the rate of the elevational shift has been debated [Citation8,Citation9]. Changes in distribution of alpine organisms are often documented by resurveys of mountain summits for which species composition was reported in the past [Citation3,Citation10–16]. On the one hand, a tendency towards increase of alpha diversity and homogenization of community composition has been observed, especially for plants on the summits of European Alps [Citation10]. On the other hand, the small-range and cold-adapted components of these mountaintop communities are particularly threatened by climate change, as warming increases with elevation [Citation17] and they have nowhere else to go to find a suitable habitat.
In this context, Edward Whymper’s expedition to Ecuador in 1879/80 offers a unique opportunity to track changes over more than a century. The expedition of this famous British mountaineer and naturalist [Citation18] took place at the end of the Little Ice Age cold period [Citation19], when summits currently devoid of permanent snow were still ice or snow-capped. On 2 February 1880, Whymper ascended Mt. Corazón, a volcano situated in the Andes of northern Ecuador [Citation20, p.108–112]. Corazón (4788 m a.s.l., S0° 31.916ʹ W78° 39.632ʹ) is a highly eroded remnant of a Pleistocene stratovolcano [Citation21], located in the Western Cordillera (). Whymper made a careful survey of the very summit of the mountain, trying to collect any living organism, including mosses, lichens, vascular plants, and soil arthropods. He reported the occurrence of six vascular plant species: Arenaria dicranoides, Werneria sp., Draba obovata, Draba imbricata, Draba indet., and Phlegmariurus crassus, the latter three species claimed to be abundant [Citation20, p.111 and 352–3]. Whymper also mentioned the presence of a large Culcitium “with thick, woolly leaves, and a nearly white, pendent, downy flower”, almost certainly Senecio canescens, “within a few feet of the extreme top of Corazón”, but could not preserve the collected specimens [Citation20, p.213]. He annotated that judging from their abundance and vigorous condition, these plants (all of which belong to typical species of the upper superpáramo in Ecuador [Citation22]) might have attained a greater elevation if there had been higher ground in the vicinity. Whymper also collected five lichens and five mosses (not studied), an earthworm, a Hemiptera and some spiders (not studied) and one ground beetle, Dyscolus diopsis.
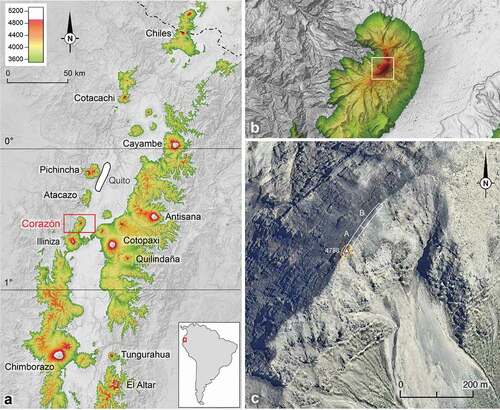
Figure 1. Localization of the study site. a: Superpáramo sky islands (orange and red areas) in Northern Ecuador. b: Detail of Mt. Corazón in the former map. c: Detail of the summit area (satellite image: ©2020 Google Earth)
Besides climate change, mountain tourism is a potential threat to the biodiversity of Mt. Corazón. Being easily accessible from Quito and the central Andean valley, it is a popular day-trip destination for hikers. Its summit can be reached from two sides, northeast and southeast, following well-marked trails. An internet search with the words “hiking”, “Corazón” and “Ecuador” yields >40 websites describing the Corazón hike or offering guide service for climbing it (consulted on 29 December 2020).
The first objective of this study was to retrace Whymper’s steps on Corazón in order to compare his observations with the current state of the superpáramo on the summit of this mountain. We limited the study to vascular plants and ground beetles (Coleoptera, Carabidae) because they are the only taxa collected by Whymper at high elevations that have been the subject of recent taxonomic revisions. Our second objective was to provide a baseline for future resurveys, in order to monitor changes in species distribution and community composition on a small superpáramo summit area, threatened both by ongoing climate warming and by mountain tourism.
Material and methods
The applicability of historical distribution data for comparative studies in climate change studies has been subject to discussion, due to uncertainties about their completeness, reliability and reproducibility [Citation23]. Errors caused by wrong plot relocation are frequent [Citation24]. In this regard, the accuracy of Whymper’s data is exceptional, as he registered with extreme care the date, place and elevation of every botanical or zoological sample he made [Citation6].
Prior to the fieldwork, we studied Whymper’s archives and historic collections to complete the information already available in his book published in 1892. His travel journal was consulted in May 2019 at the Scott Polar Research Institute of Cambridge University (manuscript collection No. MS 822/7/1). The circumstances of the survey are described in this journal with more details than in the published account. Whymper reached the summit at 12 am and left at 3.10 pm. During these three hours he took barometric measurements, drew a sketch of the summit, and collected plants and arthropods with the help of his Italian guide J.A. Carrel. Whymper described the surveyed area as a narrow and relatively flat summit ridge, 250 feet long (≈ 80 m). Based on this information, we considered that a sampling session performed by two persons over an 80 m long summit area for one hour and a half, subtracting the time spent by Whymper on weather measurements, topographic work and sampling of volcanic rock, would be a fairly accurate reproduction of the 1880 survey conditions.
Whymper’s entomological material is kept at the Natural History Museum (London), and at the Muséum National d’Histoire Naturelle (Paris) [Citation6]. The unique ground beetle specimen from the summit of Corazón was revised in Paris. The vascular plants collected by Whymper were revised at the British Museum (BM) and Kew (K) herbaria in May 2019. Of the seven species claimed to have been found on Corazón, only one voucher could be localized at K, that of Phlegmariurus crassus (named Lycopodium saururus in Whymper’s list [Citation20]). For the other species, we can only rely on the primary identifications made by W. Carruthers in 1891 and published the following year [Citation20]. Whereas Arenaria dicranoides (collected) and Senecio canescens (described) are species that cannot be mistaken and the identification provided by Whymper [Citation20] is thus reliable, identity of Draba sp. and Werneria sp. cannot be confirmed without examination of a herbarium voucher.
On 17 January 2020, we resurveyed the summit of Corazón in conditions as close as possible to those of 1880. The search was made in ca 10-meter diameter around the summit and in a 5-meter broad band stretching to both sides of the ridge to a distance of 80 m from the summit (, A). In this 500 m2 area, two botanists spent 45 minutes recording all vascular plant species present. Two entomologists spent 45 minutes searching and collecting ground beetles by hand, beneath stones and dead leaves. This simple and effective sampling method [Citation25] was likely also used by Whymper in 1880, as in another occasion he reported the discovery of carabid beetle specimens on the summit of the Pichincha volcano by turning over stones and rocks [Citation20]. Identifications of ground beetles were based on the keys and taxonomic descriptions in [Citation26,Citation27].
Additionally, to get a more complete baseline inventory of the flora and fauna of the Corazón upper superpáramo, collections were made in a broader area around the summit. Along the summit ridge, plants were further recorded for 45 minutes by the same two botanists from 80 to 160 m north of the summit (, B and ). Along the southern trail, from the lower elevation limit of the superpáramo to the summit area, the uppermost elevation occurrence of every observed plant species was recorded by two botanists (Supplemental material 1), and ground beetles were collected by two entomologists at two elevations, 4500 m and 4600 m a.s.l., following the same protocol as on the summit (Supplemental material 2).
Table 1. List of the vascular plant species encountered in January 2020 within a 0–80 m distance from the summit of Mt. Corazón (, zone A) and within an 80–160 m distance (zone B). Species collected by Whymper in 1880 are in bold. Voucher numbers refer to P. Sklenář’s collections deposited at QCA
Results
Degradation of the summit area
The very summit of Corazón and part of the summit ridge were heavily altered by visitors as a consequence of trampling and stone removal, and no vascular plants were present there. The summit itself was a 200 m2 large area of sandy soil bare of plants, where most of the stones had been moved to build a cairn at the uppermost point (). Further north, the summit ridge was also damaged by trampling. It was devoid of any plants, although three species were encountered just beside the ridge, i.e. Werneria nubigena, Draba obovata, and Luzula racemosa. As a whole, the deteriorated part amounted to 70% of the sampled area. This situation appears to be recent. Whymper’s description does not mention any alteration of the summit area, except for the presence of two dressed rocks [Citation20, p.110]. A photograph taken close to the summit in January 1986 shows several Senecio canescens plants that have disappeared since then (), and according to a professional mountain guide with a 20-year experience in Ecuadorian Andes (J. Larrea Yánez, personal communication, 29 December 2020), the width of the trail to the summit has widened considerably in recent years.
Figure 2. The summit of Mt Corazón in January 2020, viewed from the south

Figure 3. Clump of Senecio canescens near the summit of Mt. Corazón in January 1986. Photograph taken by P. Moret

Vascular plants
Around the very summit and within the 0–80 m distance from it (, A), i.e. in the area surveyed by Whymper in 1880, 22 vascular plant species were recorded. Twelve more species were observed further north (80–160 m north of the summit), in the same environment ().
We encountered three species of Brassicaceae, i.e. Draba obovata, D. aretioides, and Eudema nubigena, all of them quite common. Whymper reported three species of Draba from the summit of the mountain: Draba imbricata C.A. Mey (reported to be common), D. obovata Benth., and another common but not identified species. Whymper’s collection of Draba obovata from the summit of Corazón was confirmed by a specialist (Al-Shehbaz 2018). Draba imbricata is a species from Caucasus and some of its forms resemble D. aretioides (Al-Shehbaz, pers. comm. 2020). We may therefore assume with a reasonable confidence that Whymper’s plant was not D. imbricata but rather D. aretioides. A species listed as Draba sp.? was collected by Whymper on the Chimborazo volcano and later identified as Eudema nubigena by Al-Shehbaz (vouchers were also revised and identification confirmed by PS), which is a common superpáramo species with a Draba-like habit. Since there is no report of other species of Draba from Corazón than D. aretioides and D. obovata [Citation28], we assume that the unidentified Draba listed by Whymper was in fact E. nubigena. Four species of Werneria (including Xenophyllum) were encountered in 2020 on the summit of Corazón. It is a reasonable assumption that one of these species may correspond to the unidentified Werneria sp. reported by Whymper. If that assumption and our conclusions on Whymper’s collections of Draba are correct, then the seven species reported by Whymper to occur on the summit of Corazón in 1880 were all present there in 2020. This case highlights the crucial importance of vouchers in comparative studies on species diversity, not only in still relatively poorly taxonomically known areas, such as the equatorial high Andes.
Arenaria dicranoides is a typical upper superpáramo species and apparently was not reported from Corazón since Whymper (P. Sklenář, unpubl. data). We found only one specimen of this species growing 160 m away from the summit. This suggests that abundance of this species has declined in the summit area; it was not encountered along the southern summit ridge although it might be present on other slopes of the mountain, which were not explored by us.
Additionally, Corazón represents a new locality for two species which are endemic to the superpáramo of Ecuador. Pentacalia microdon is reported for the first time from another locality than Cotopaxi and Illiniza, and Xenophyllum rigidum is reported for the first time from another locality than Chimborazo, Antisana, and Illiniza (J. Calvo, pers. comm. 2020). As both species produce achenes with pappus, seed dispersal by wind to Corazón is possible. However, the nearby Cotopaxi and Illiniza volcanoes, where one or both species occur, respectively, are frequent destinations of hikers. Dispersal of seeds by tourists thus cannot be ruled out.
Ground beetles
Four ground beetle species were collected in the summit area of Corazón, all members of the genus Dyscolus: D. diopsis (Bates), D. megacephalus (Bates), Dyscolus oopteroides (Chaudoir) and D. rotundiceps (Bates). Seven more species were collected at lower elevations (4500 and 4600 m a.s.l.) in the upper superpáramo along the southern ridge (Supplemental material 2). Only two species, Dyscolus oopteroides and D. rotundiceps, were present at all surveyed elevations. Abundance of two species, Dyscolus oopteroides and D. diopsis, increases with the elevation. Dyscolus diopsis, the only species collected by Whymper, is currently by far the dominant carabid species in the summit area, amounting to 68% of the recorded individuals, whilst it was not recorded below 4600 m a.s.l. (Supplemental material 2).
Although ground beetles were generally scarce in the summit area, small local clusters have been observed beneath stones and dead leaves at the foot of Senecio canescens plants. This kind of microhabitat maintains a higher humidity and/or buffers its oscillation in contrast to the surrounding open environment. Survival of the ground beetle community in the summit area is therefore dependent on the presence of healthy vegetation and of the associated leaf litter.
Discussion
Whymper recorded much less species than we did: 7 vascular plant species vs 22, and 1 ground beetle species vs 4. Several reasons can explain this difference.
i) Whymper’s and Carrel’s skills as plant and beetle collectors were likely to be lesser than ours. Whymper was an experienced naturalist: as soon as 1862, at the beginning of his career as an alpinist, he was able to identify the plants he collected close to the snow-line on the Matterhorn [Citation29]. In 1867, when leading a British expedition to Greenland, he was accompanied by a botanist, Robert Brown, with whom he collected plants [Citation18]. Still he was not a fully trained botanist, and in particular, it is fairly possible that he was not able to differentiate between species that look alike at first sight (e.g. cushion-forming Xenophyllum humile vs X. rigidum).
ii) Whymper did not pay attention to grasses and other graminoid monocotyledonous species. Three grass species were encountered in 2020, and among them Luzula racemosa and Calamagrostis aurea were abundant.
iii) Whymper reported in his journal that “considerable snow beds” covered the western side of the summit ridge. Other historical reports by James Orton, Wilhelm Reiss, Alfons Stübel and Theodor Wolf, ranging from 1867 to 1892, confirm that at that time a significant part of the Corazón summit was permanently covered by snow [Citation30, p.103]. The perennial snow seems to have disappeared gradually over the following decades [Citation30, p.28]. The space available for plants to grow was therefore considerably lower in 1880 than in 2020, and the vicinity of the permanent snow line may then have limited the strength of the vegetation.
iv) Species richness was actually lower in 1880. This last hypothesis would be in accordance with the increase in species richness, due to upslope range shifts of species from lower elevations, which has been observed over the last decades on many mountain summits of higher latitudes [Citation10,Citation31]. Yet the part played by each of these factors cannot be disentangled with our methods, so the question whether any of the 15 species of vascular plants and 3 species of ground beetles not recorded by Whymper was already present there in 1880 will remain unanswered.
From a retrospective point of view, these biases in the interpretation of Whymper’s data do not allow us to evaluate in a more exact way changes in plant and insect communities between 1880 and 2020. The questionable reliability of Whymper’s data is, furthermore, enhanced by the uncertainty resulting from the impact of tourism, exemplified by the possible introduction of two plant species, Pentacalia microdon and Xenophyllum rigidum. However, from a prospective point of view, the current faunistic and floristic data presented in this contribution can be used as a baseline for future resurveys of Corazón, to monitor changes in the species distribution and community composition of its summit area, in the same line as recent studies on Andean [Citation32] or Guiana Shield [Citation33] summits.
All the ground beetle species observed on Corazón are strictly limited to the Pichincha–Chimborazo area of endemism [Citation34]. They are currently represented by isolated populations on several superpáramo sky islands, ranging from Cayambe to Llanganatis in the Eastern Cordillera and from Cotacachi to Chimborazo in the Western Cordillera. Regarding the composition of these ground beetle communities, a comparison can be made with the summit of Mt. Guagua Pichincha, near Quito, which peaks at 4784 m a.s.l. [Citation6,Citation25 and unpublished data]. From 4500 m a.s.l. upwards, Corazón has 11 species, Pichincha 12. Seven species are present in both sky islands (Dyscolus diopsis, D. maleodoratus, D. oopteroides, D. rotundiceps, D. megacephalus, Blennidus strictibasis and Bembidion fulvocinctum); seven are microendemic species exclusive to one or the other (Dyscolus verecundior, Blennidus sp. 1 and Oxytrechus sp. 1 on Corazón; Dyscolus verecundus, Blennidus mucronatus, Oxytrechus sp. 2 and O. sp. 3 on Pichincha); and the last two are widespread species only present on one of them (D. denigratus on Corazón, D. orthomus on Pichincha). On the very summit, diversity appears to be higher on Pichincha (6 vs 4 species), possibly due to a higher sampling effort, but relative abundance is quite similar for Dyscolus diopsis, D. oopteroides and D. megacephalus. The mountaintop communities thus appear to be similar at Pichincha and at Corazón. No such comparison could be made for the vascular plants of the Guagua Pichincha summit zone, since available datasets come from a secondary summit, Padre Encantado, situated ca. 250 m below the top elevation of Corazón and of Guagua Pichincha. The vegetation of the Padre Encantado summit, classified in two upper superpáramo communities (i.e. Festuca glumosa–Luzula racemosa and Arenaria dicranoides–Senecio canescens [Citation35]), shares the majority of its species with the summit area of Corazón, although the vegetation at Padre Encantado may be denser and more diverse on the account of its lower elevation [Citation32].
Corazón is especially exposed to the effects of climate change, owing to the very small area of its superpáramo. In northern Ecuador, a similar situation may occur in other eroded stratovolcanoes that peak between 4600 and 4900 m a.s.l.: from north to south, Chiles, Cotacachi, Sara Urcu, Imbabura, Pichincha, Sincholagua, Rumiñahui and Quilindaña. Each of these small isolated superpáramos hosts a unique species assemblage, and some of them possess exclusive microendemic species [Citation26]. Due to the pyramid-shape hypsography of these mountains [Citation36], the habitable area greatly reduces with increasing elevation. With the ongoing climate changes, the area occupied by the superpáramo will probably shrink due to the upslope shift of its lower limit, thus accelerating the risk of local extirpation of high-elevation stenotherm specialists [Citation5,Citation37], and of total extinction of precinctive species. In the case of Corazón, this would certainly be the case of the carabid Dyscolus diopsis which only lives there above 4600 m a.s.l. Moreover, some of these peaks are frequently climbed by mountaineers and hikers, especially Pichincha and Corazón, which results in severe damages by trampling. Destruction of natural habitats in the summit area adds another threat on these endangered species, as the summits would be their ultimate refuge before extinction.
Disclosure of potential conflicts of interest
No potential conflict of interest was reported by the author(s).
Author contributions
PMo designed the research; PS and PMo studied the historical plant and insect collections; all the authors performed the fieldwork; PS, RJ and PMu identified the plant specimens; PMo and WP identified the insect specimens; all the authors analysed the data; PS and PMo wrote the manuscript with contributions from all authors.
Supplemental Material
Download MS Word (27.5 KB)Acknowledgments
We thank Joel Calvo for confirmation of determinations and information about species distribution, Mauricio Diazgranados and Nicholas Hind for their assistance at the Royal Botanic Gardens, Kew, as well as Thierry Deuve at the Muséum national d’histoire naturelle, Paris. We also thank Jacobo Larrea Yánez for his advice and Josué Franco for his help in the fieldwork. The Ministry of the Environment of Ecuador provided research and collection permits (MAE-DNB-CM-2018-0082, 014-2019-IC-FAU-DPAP-NA and 009_invertebrados-2019-IC-FAU-DNB/MA).
Supplementary material
Supplemental data for this article can be accessed here
Additional information
Funding
References
- Chen IC, Hill JK, Ohlemüller R, et al. Rapid range shifts of species associated with high levels of climate warming. Science. 2011;333(6045):1024–1026.
- Buytaert W, Cuesta-Camacho F, Tobón C. Potential impacts of climate change on the environmental services of humid tropical alpine regions. Glob Ecol Biogeogr. 2011;20(1):19–33.
- Pauli H, Gottfried M, Dullinger S, et al. Recent plant diversity changes on Europe’s mountain summits. Science. 2012;336(6079):353–355.
- Báez S, Jaramillo L, Cuesta F, et al. Effects of climate change on Andean biodiversity: a synthesis of studies published until 2015. Neotrop Biodivers. 2016;2:181–194.
- Peyre G, Lenoir J, Karger DN, et al. The fate of páramo plant assemblages in the sky islands of the northern Andes. J Veg Sci. 2020 April;18. DOI:https://doi.org/10.1111/jvs.12898
- Moret P, Aráuz MA, Gobbi M, et al. Climate warming effects in the tropical Andes: first evidence for upslope shifts of Carabidae (Coleoptera) in Ecuador. Insect Conserv Divers. 2016;9(4):1–9. https://doi:
- Morueta-Holme N, Engemann K, Sandoval-Acuña P, et al. Strong upslope shifts in Chimborazo’s vegetation over two centuries since Humboldt. Proc Natl Acad Sci USA. 2015;112(41):12741–12745.
- Sklenář P. Advance of plant species on slopes of the Chimborazo volcano (Ecuador) calculated based on unreliable data. Proc Natl Acad Sci USA. 2016;113(4):E407–E408.
- Moret P, Muriel P, Jaramillo R, et al. Humboldt’s tableau physique revisited. Proc Natl Acad Sci USA. 2019;116(26):12889–12894.
- Jurasinski G, Kreyling J. Upward shift of alpine plants increases floristic similarity of mountain summits. J Veg Sci. 2007;18(5):711–718.
- Gottfried M, Pauli H, Futschik A, et al. Continent-wide response of mountain vegetation to climate change. Nat Clim Change. 2012;2(2):111–115.
- Stanisci A, Frate L, Morra Di Cella U, et al. Short-term signals of climate change in Italian summit vegetation: observations at two GLORIA sites. Plant Biosyst. 2016;150(2):227–235.
- Czortek P, Kapfer J, Delimat A, et al. Plant species composition shifts in the Tatra Mts as a response to environmental change: a resurvey study after 90 years. Folia Geobot. 2018;53(3):333–348.
- Lamprecht A, Semenchuk PR, Steinbauer K, et al. Climate change leads to accelerated transformation of high-elevation vegetation in the central Alps. New Phytol. 2018;220(2):447–459.
- Rumpf SB, Hülber K, Klonner G, et al. Range dynamics of mountain plants decrease with elevation. Proc Natl Acad Sci USA. 2018;115(8):1848–1853.
- Salick J, Fang Z, Hart R. Rapid changes in eastern Himalayan alpine flora with climate change. Am J Bot. 2019;106(4):520–530.
- Pepin N, Bradley R, Diaz H, et al. Elevation-dependent warming in mountain regions of the world. Nat Clim Change. 2015;5(5):424–430.
- Smith I. Shadow of the Matterhorn. The life of Edward Whymper. Hildersley (UK): Carreg Limited; 2011.
- Rabatel A, Francou B, Soruco A, et al. Current state of glaciers in the tropical Andes: a multi-century perspective on glacier evolution and climate change. Cryosphere. 2013;7(1):81–102.
- Whymper E. Travels amongst the great Andes of the Equator. London: John Murray; 1892.
- Hall ML, Samaniego P, Le Pennec JL, et al. Ecuadorian Andes volcanism: a review of Late Pliocene to present activity. J Volcanol Geotherm Res. 2008;176(1):1–6.
- Sklenář P, Balslev H. Superpáramo plant species diversity and phytogeography in Ecuador. Flora. 2005;200(5):416–433.
- Stöckli V, Wipf S, Nilsson C, et al. Using historical plant surveys to track biodiversity on mountain summits. Plant Ecolog Divers. 2011;4(4):415–425.
- Kapfer J, Hédl R, Jurasinski G, et al. Resurveying historical vegetation data – opportunities and challenges. Appl Veg Sci. 2017;20(2):164–171.
- Gobbi M, Barragán A, Brambilla M, et al. Hand searching versus pitfall trapping: how to assess biodiversity of ground beetles (Coleoptera: Carabidae) in high altitude equatorial Andes? J Insect Conserv. 2018;22(3–4):533–543.
- Moret P. Los coleópteros Carabidae del páramo en los Andes del Ecuador. Sistemática, ecología y biogeografía. Quito: Pontificia Universidad Católica del Ecuador; 2005.
- Moret P, Murienne J. Integrative taxonomy of the genus Dyscolus (Coleoptera, Carabidae, Platynini) in Ecuadorian Andes. Eur J Taxon. 2020;646:1–55.
- Al-Shehbaz I. A monograph of the south American species of draba (Brassicaceae). Ann Mo Bot Gard. 2018;103(4):463–590.
- Whymper E. Scrambles amongst the Alps: in the years 1860–’69. Philadelphia (PA): J.B. Lippincott & Co; 1872.
- Hastenrath S. The glaciation of the Ecuadorian Andes. Rotterdam: A.A. Balkema; 1981.
- Steinbauer MJ, Grytnes JA, Jurasinski G, et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature. 2018;556(7700):231–234.
- Cuesta F, Muriel P, Llambí LD, et al. Latitudinal and altitudinal patterns of plant community diversity on mountain summits across the tropical Andes. Ecography. 2017;40(12):1381–1394.
- Safont E, Rull V, Vegas-Vilarrubia T, et al. Establishing a baseline of plant diversity and endemism on a neotropical mountain summit for future comparative studies assessing upward migration: an approach from biogeography and nature conservation. Syst Biodivers. 2014;12(3):292–314.
- Moret P. Altitudinal distribution, diversity and endemicity of Carabidae (Coleoptera) in the páramos of Ecuadorian andes. Ann Soc entomol Fr. (ns). 2009;45(4):500–510.
- Sklenář P Vegetation ecology and phytogeography of Ecuadorian superpáramos. PhD thesis, Charles University, Prague; 2000.
- Elsen PR, Tingley MW. Global mountain topography and the fate of montane species under climate change. Nat Clim Change. 2015;5(8):772–776.
- Urban MC. Escalator to extinction. Proc Natl Acad Sci USA. 2018;115(47):11871–11873.