
Abstract
We determined the complete chloroplast genome of Pedicularis hallaisanensis (Orobanchaceae), a hemi-parasitic perennial herb. This genome is 143,469 bp long and features a large single-copy region (81,664 bp) and a small single-copy region (12,203 bp), separated by two inverted-repeat regions (24,801 bp each). It contains 115 genes – 70 for coding, eight for rRNA, and 37 for tRNA. However, 11 ndh genes have been pseudogenized, truncated, or deleted. Our phylogenetic tree showed that these hemi-parasitic plants are sister to holo-parasitic genera within Orobanchaceae.
Pedicularis hallaisanensis Hurus. (Orobanchaceae) is a perennial herb that is endemic but endangered in Korea (Cho and Choi Citation2011). This species is root-hemi-parasitic, similar to other Pedicularis members. Although the chloroplast (cp) genomes of land plants are highly conserved (Jansen and Ruhlman Citation2012), the genomes of parasitic plants, such as those within Orobanchaceae, have experienced remarkable changes in size, structure, and gene contents (Xie et al. Citation2012; Wicke et al. Citation2013). Here, we report the complete cp genome of the hemi-parasitic P. hallaisanensis and examine its phylogenetic position within Orobanchaceae.
Plant material of P. hallaisanensis was collected from Mt. Halla on Jeju Island, Korea (N 33°21′49″, E 126°31′44″), and a voucher specimen was deposited in the herbarium of Inha University (Cho. 98454). Genomic DNA was extracted from young, silica gel-dried leaves through a protocol that used a DNeasy Plant Mini Kit (Qiagen, Seoul, Korea). The extracted DNA was sequenced via the Mi-Seq Illumina platform (LAS, Seoul, Korea), generating 11,213,054 raw reads. The cp genome was assembled using Geneious 10.2.3 software (Kearse et al. Citation2012) and reference sequences of P. ishidoyana (NC029700) and Lathraea squamaria (NC027838). Gene annotations were made with the DOGMA program (Wyman et al. Citation2004), and were manually corrected for start and stop codons and for intro/exon boundaries. Using default parameters and 1000 bootstrap replicates, we constructed a phylogenetic tree based on maximum likelihood (ML) analysis that included RAxML-HPC v.8 (Stamatakis Citation2014), which is available from the CIPRES gateway (Miller et al. Citation2010).
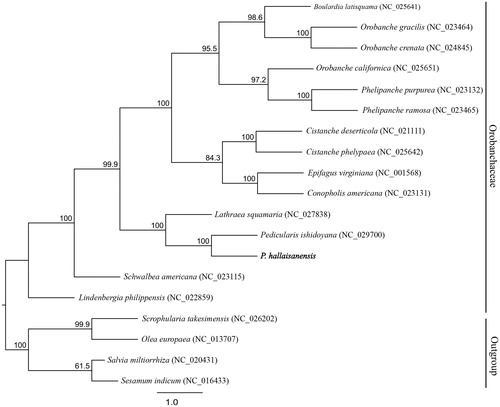
The complete cp genome of P. hallaisanensis (GenBank: MG770330) is 143,469 bp long, and shows a quadripartite structure with two inverted-repeat regions (24,801 bp each) that separate a large single-copy region (81,664 bp) and a small single-copy region (12,203 bp). This cp genome contains 115 genes, i.e. 70 for coding, eight for ribosomal RNA, and 37 for transfer RNA. All 11 of its ndh genes have been pseudogenized (ndhB, ndhE, ndhH, ndhG, ndhJ, and ndhK), truncated (ndhA and ndhD), or deleted (ndhC, ndhF, and ndhI), a phenomenon also seen with other parasitic species (Krause Citation2011). Our phylogenetic tree includes 61 coding genes extracted from the cp genome of P. hallaisanensis, plus 18 related species downloaded from the NCBI database in Lamiales. The ML tree demonstrates that Pedicularis is sister to Lathraea squamaria, based on their high bootstrap values (). These new phylogenetic data provide insight into the evolutionary progress of Orobanchaceae.
Figure 1. Phylogenetic tree constructed with RAxML, using dataset of 61 concatenated protein-coding regions from 19 chloroplast genomes. Numbers above nodes indicate bootstrap values (1000 replicates) > 50%.
Disclosure statement
The authors report no conflicts of interest. They, alone, are responsible for the content and writing of this paper.
Additional information
Funding
Related Research Data
References
- Cho WB, Choi BH. 2011. Taxonomic position of Pedicularis hallaisanensis Hurusawa, an endemic plant of Mt. Halla. Korean J Plant Taxon. 41:130–137 (in Korean).
- Jansen RK, Ruhlman TA. 2012. Plastid genomes of seed plants. In: Genomics of Chloroplasts and Mitochondria. Amsterdam (The Netherlands): Springer. pp. 103–126.
- Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, et al. 2012. Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 28:1647–1649.
- Krause K. 2011. Piecing together the puzzle of parasitic plant plastome evolution. Planta. 234:647–656.
- Miller MA, Pfeiffer W, Schwartz T. 2010. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans (LA). 1–8.
- Stamatakis A. 2014. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 30:1312–1313.
- Wicke S, Muller KF, de Pamphilis CW, Quandt D, Wickett NJ, Zhang Y, Schneeweiss GM. 2013. Mechanisms of functional and physical genome reduction in photosynthetic and nonphotosynthetic parasitic plants of the broomrape family. Plant Cell. 25:3711–3725.
- Wyman SK, Jansen RK, Boore JL. 2004. Automatic annotation of organellar genomes with DOGMA. Bioinformatics. 20:3252–3255.
- Xie XF, Yan HF, Wang FY, Ge XJ, Hu CM, Hao G. 2012. Chloroplast DNA phylogeography of Primula ovalifolia in central and adjacent southwestern China: past gradual expansion and geographical isolation. J System Evol. 50:284–294.