
Abstract
Ilex cornuta Lindl. & Paxton is a species distributed in eastern China and Korea, utilized as traditional medical plants as well as horticultural species. Here, we completed chloroplast genome of I. cornuta. Its length is 157,224 bp long and has four subregions: 86,610 bp of large single copy (LSC) and 18,429 bp of small single copy (SSC) regions are separated by 26,092 bp of inverted repeat (IR) regions including 131 genes (86 protein-coding genes, eight rRNAs, and 37 tRNAs). The overall GC content of this chloroplast genome is 37.7% and those in the LSC, SSC, and IR regions are 35.7%, 31.9%, and 42.9%, respectively. Phylogenetic trees show that I. cornuta is clustered in Section Ilex clade clearly. In addition, two Ilex species from South America are not in a basal clade, which is different from previous phylogenetic study, suggesting more Ilex chloroplast genomes are required to understand phylogenetic relationships of Ilex species.
Ilex cornuta Lindl. & Paxton, distributed only in eastern China and Korea (Hu Citation1949), has been utilized as a horticultural species because its leaves are distinctive rectangular foliage (one or two spines per side; Yang and Peng Citation2019) and its fruits are red berries (Hu Citation1949). Many cultivars and hybrids have been developed based on this species (Hodges et al. Citation2001). Together with Ilex aquiforlium usually distributed in Europe, it is a typical species for Christmas tree inside home. In addition, it has been utilized as medical plant in China so that it contains several useful compounds (Wenjuan et al. Citation1986; Alikaridis Citation1987).
Till now, nine complete chloroplast genomes of Ilex were sequenced: Seven of nine species are from China (Yao et al. Citation2016) and two are from South America (Cascales et al. Citation2017). Chloroplast genome of I. cornuta was also analyzed one time; however, they could not complete its chloroplast genome due to lack of sample (Cascales et al. Citation2017).
Total DNA of I. cornuta isolated from Chollipo arboretum in Tae-an gun, South Chungcheong Province, Korea (Voucher in InfoBoss Cyber Herbarium (IN); S. Nam, IB-00590) was extracted from fresh leaves using a DNeasy Plant Mini Kit (QIAGEN, Hilden, Germany). Genome was sequenced using HiSeqX at Macrogen Inc., Korea, and de novo assembly and confirmation were performed by Velvet 1.2.10 (Zerbino and Birney Citation2008), SOAPGapCloser 1.12 (Zhao et al. Citation2011), BWA 0.7.17 (Li Citation2013), and SAMtools 1.9 (Li et al. Citation2009). Geneious R11 11.0.5 (Biomatters Ltd., Auckland, New Zealand) was used for annotation based on Ilex paraguariensis chloroplast, complete genome (KP016928; Cascales et al. Citation2017).
The chloroplast genome of I. cornuta (Genbank accession is MK335536) is 157,224 bp (GC ratio is 37.7%) and has four subregions: 86,610 bp of large single copy (LSC; 35.7%) and 18,429 bp of small single copy (SSC; 31.9%) regions are separated by 26,092 bp of inverted repeat (IR; 42.9%). It contains 131 genes (86 protein-coding genes, eight rRNAs, and 37 tRNAs); 18 genes (seven protein-coding gene, four rRNAs, and seven tRNAs) are duplicated in IR regions.
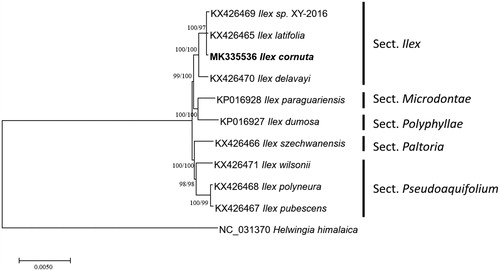
Ten Ilex and one Helwingia chloroplast genomes from Aquifoliaceae were used for constructing neighbor joining (bootstrap repeat is 10,000) and maximum likelihood (bootstrap repeat is 1,000) phylogenetic trees using MEGA X (Kumar et al. Citation2018) after aligning whole chloroplast genome sequences using MAFFT 7.388 (Katoh and Standley Citation2013). Phylogenetic trees show that I. cornuta is clustered with Ilex latifolia and new species of Ilex (Section Ilex in ; Yao et al. Citation2016), agreeing with previous studies (Cuénoud et al. Citation2000; Jiang et al. Citation2017) in Section Ilex (Wu et al. Citation2008). In addition, Ilex dumosa and Ilex paraguariensis originated from South America are not in basial clade of Ilex genus with high bootstrap supports (), disagreeing with the previous study (Gottlieb et al. Citation2005). Due to lack of phylogenetic study of Ilex genus covering both Eastern Asia and South America samples at the same time, additional Ilex chloroplast genomes are strongly required to uncover Ilex phylogenetic relationship.
Figure 1. Neighbor joining (bootstrap repeat is 10,000) and maximum likelihood (bootstrap repeat is 1,000) phylogenetic trees of ten Ilex and one Helwingia chloroplast genomes from Aquifoliaceae: Ilex cornuta (MK335536 in this study), Ilex latifolia (KX426465), Ilex sp. XY-2016 (KX426469), Ilex delavayi (KX426470), Ilex paraguaiensis (KP016928), Ilex dumorsa (KP016927), Ilex szechwanensis (KX426466), Ilex wilsonii (KX426471), Ilex polyneura (KX426468), Ilex pubescens (KX426467), and Helwingia himalaica (NC_031370). Section names were displayed in the right side of phylogenetic tree (Gottlieb et al. Citation2005; Wu et al. Citation2008; Jiang et al. Citation2017). Phylogenetic tree was drawn based on neighbor joining tree. The numbers above branches indicate bootstrap support values of maximum likelihood and neighbor joining phylogenetic trees, respectively.
Disclosure statement
The authors declare that they have no competing interests.
Additional information
Funding
Related Research Data
References
- Alikaridis F. 1987. Natural constituents of Ilex species. J Ethnopharmacol. 20:121–144.
- Cascales J, Bracco M, Garberoglio MJ, Poggio L, Gottlieb AM. 2017. Integral phylogenomic approach over Ilex L. species from southern South America. Life. 7:47.
- Cuénoud P, Del Pero Martinez MA, Loizeau P-A, Spichiger R, Andrews S, Manen J-F. 2000. Molecular phylogeny and biogeography of the genus Ilex L. (Aquifoliaceae). Ann Bot. 85:111–122.
- Gottlieb AM, Giberti GC, Poggio L. 2005. Molecular analyses of the genus Ilex (Aquifoliaceae) in southern South America, evidence from AFLP and ITS sequence data. Am J Bot. 92:352–369.
- Hodges G, Ruter JM, Kristine Braman S. 2001. Susceptibility of Ilex species, hybrids and cultivars to Florida wax scale (Ceroplastes floridensis Comstock). J Environ Horticult. 19:32–36.
- Hu S-y. 1949. The genus Ilex in China. J Arnold Arboretum. 30:233–344.
- Jiang L, Xu K, Fan Q, Peng H. 2017. A new species of Ilex (Aquifoliaceae) from Jiangxi Province, China, based on morphological and molecular data. Phytotaxa. 298:147–157.
- Katoh K, Standley DM. 2013. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 30:772–780.
- Kumar S, Stecher G, Li M, Knyaz C, Tamura K. 2018. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol. 35:1547–1549.
- Li H. 2013. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv preprint arXiv:13033997.
- Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R. 2009. The sequence alignment/map format and SAMtools. Bioinformatics. 25:2078–2079.
- Wenjuan Q, Xiue W, Junjie Z, Fukuyama Y, Yamada T, Nakagawa K. 1986. Triterpenoid glycosides from leaves of Ilex cornuta. Phytochemistry. 25:913–916.
- Wu Z, Raven P, Hong D. 2008. Flora of China. Vol. 11: Oxalidaceae through Aceraceae. Beijing: Science Press, and St. Louis: Missouri Botanical Garden Press.
- Yang Y, Peng H. 2019. Taxonomic notes on Ilex sect. Ilex (Aquifoliaceae) from China I: revision of four species. Nordic J Bot. https://onlinelibrary.wiley.com/toc/17561051/0/ja
- Yao X, Tan Y-H, Liu Y-Y, Song Y, Yang J-B, Corlett RT. 2016. Chloroplast genome structure in Ilex (Aquifoliaceae). Sci Rep. 6:28559
- Zerbino DR, Birney E. 2008. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 18:821–829.
- Zhao Q-Y, Wang Y, Kong Y-M, Luo D, Li X, Hao P. 2011. Optimizing de novo transcriptome assembly from short-read RNA-Seq data: a comparative study. BMC Bioinform. 12:S2.