
Abstract
The complete mitochondrial genome of the Hexagrammos agrammus is presented in this study. The mitochondrial genome is 16,512 bp long and consists of 13 protein-coding genes, 2 rRNA genes, 22 tRNA genes, and a control region. The gene order and composition were similar to those of most other vertebrates. The nucleotide compositions of the heavy strand are 17.27% of G, 26.10% of T, 26.85% of A, and 29.78% of C. With the exception of the NADH dehydrogenase subunit 6 (ND6) and 8 tRNA genes, all other mitochondrial genes are encoded on the heavy strand. The phylogenetic analysis by neighbour-joining (NJ) method showed that H. agrammus has the closer relationship with Hexagrammos otakii and Hexagrammos lagocephalus in the phylogenetic relationship.
Spotty belly greenling, Hexagrammos agrammus, is an endemic species in the northwestern Pacific, being locally distributed along the coasts of the China. This species is commercially important in the northwestern Pacific and also caught by game fishing. Several studies have been carried out regarding the morphology, systematic and ecology of the species (Kanamoto Citation1979; Crow et al. Citation2004; Kwak et al. Citation2005; Kimura et al. Citation2007). However, studies on the genetic diversity of H. agrammus have little been conducted yet. Assessments of genetic information are essential to develop strategies for the conservation and management of fisheries resources. The next-generation sequencing (NGS) technologies, such as Illumina, allow considerable numbers of sequence data to be rapidly and efficiently characterized, which make it particularly feasible for mitogenomes (Gilbert et al. Citation2007). Moreover, Illumina sequencing has been successfully used to assemble the mitogenomes of fish species (Cui et al. Citation2009). Therefore, we determined to sequence the complete mitochondrial genome of H. agrammus using the next-generation sequencing (NGS) techniques strategy in order to find new DNA markers for the studies on genetics of H. agrammus.
The specimen of H. agrammus was collected from the coastal water of Qingdao (36.41°N, 120.77°E), China, during May 2019. The examined specimen was preserved at Fisheries Ecology and Biodiversity Laboratory in Zhejiang Ocean University under specimen accession NO. ZJOU-04057 and the muscle tissue was preserved in 95% ethanol. The genomic DNA was extracted from dorsal-lateral muscles (30 mg) using Rapid Animal Genomic DNA Isolation Kit (Sangon Biotech Co., Ltd., Shanghai, CN). A genomic library was established and followed by next-generation sequencing. Whole genome resequencing (sequencing depth 50X) was conducted by using Illumina Hiseq4000 platform with the sequencing insertion of 350-bp. Quality check for sequencing data was done by FastQC (Andrews Citation2010) and the filtered clean data were assembled and mapped to complete mitogenome sequence using NOVOPlasty v3.7.2 (Dierckxsens et al. Citation2017). The assembled sequence was subsequently annotated using the online Mitochondrial Genome Database of Fish server (Iwasaki et al. Citation2013) and the MITOS Web Server (Bernt et al. Citation2013).
The final sequence has been deposited in GenBank with accession number MT363637. The complete mitochondrial genome of H. agrammus (16,512 bp in length) consists of 13 protein-coding genes, 22 transfer RNA genes (tRNA), 2 ribosomal RNA genes (12S rRNA and 16S rRNA), and 2 non-coding control regions (control region and origin of light-strand replication). The arrangement of all genes is identical to that of most vertebrates (Wang et al. Citation2008; Chen Citation2013; Chiang et al. Citation2013). Most of the genes are encoded on the heavy strand (H-strand), except for the eight tRNA genes (-Gln, -Ala, -Asn, -Cys,-Tyr, -Ser, -Glu and -Pro) and one protein-coding gene (NADH dehydrogenase subunit 6, ND6). The overall nucleotide compositions of the heavy strand in descending order are 17.27% of G, 26.10% of T, 26.85% of A, and 29.78% of C, with a slight A + T-rich feature (54.95%). All the protein-coding genes begin with an ATG start codon except for COI started with GTG. Three types of stop codons revealed are TAA (COI, ATP8, ATP6, COIII, ND4L, ND5), TAG (ND1, ND2, ND3, ND6), and T (COII, ND4, Cytb). These features are common among vertebrate mitochondrial genome, and TAA is supposed to be appeared via posttranscriptional polyadenylation (Ojala et al. Citation1981). The longest one is ND5 gene (1839 bp) among protein-coding genes, whereas the shortest is ATPase 8 gene (168 bp). The two ribosomal RNA genes, 12S rRNA (947 bp), and 16S rRNA (1,692 bp), are located between tRNA-Phe (GAA) and tRNA-Leu (TAA), and are separated by the tRNA-Val gene with the same situation found in other vertebrates. Most genes are either abutted or overlapped. The 22 tRNA genes vary from 66 to 74 bp in length. All these could be folded into the typical cloverleaf secondary structure although numerous non-complementary and T–G base pairs exist in the stem regions. The control region was 839 bp in length, located between tRNA-Pro (TGG) and tRNA-Phe (GAA) gene. The nucleotide composition of control region was 32.65% of A, 20.26% of C, 16.45% of G, 30.63% of T.
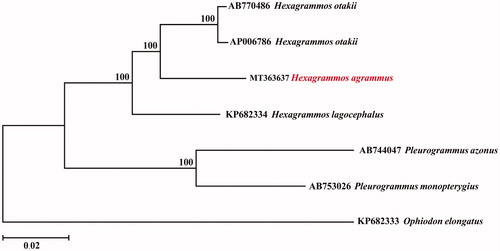
To confirm the phylogenetic position of H. agrammus among family Hexagrammidae, a neighbour-joining (NJ) tree was reconstructed using MEGA6 (Tamura et al. Citation2013), with the complete mtDNA sequences from 7 species of family Hexagrammidae. As shown in , the H. agrammus has a relatively closer relationship with Hexagrammos otakii and Hexagrammos lagocephalus. Furthermore, the species among Hexagrammos and Pleurogrammus clustered into a monophyletic clade suggested the closer relationship between them than between Ophiodon elongatus. The information of the mitogenome will be beneficial for future phylogenetic studies and specimen identification of Hexagrammidae species.
Figure 1. Phylogenetic relationships (neighbor-joining tree) for 7 species of family Hexagrammidae. Numbers on each node are bootstrap values of 1000 replicates.
Acknowledgements
We thank Dr. Linlin Zhao for sample collection and we are grateful to Dr. Zhi Chen for the help of sequence analysis.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The data that support the findings of this study is openly available in GenBank of NCBI at https://www.ncbi.nlm.nih.gov under accession number MT363637.
Additional information
Funding
References
- Andrews S. 2010. FastQC: A quality control tool for high throughput sequence data. [accessed 2018 Oct 4]. http://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- Bernt M, Donath A, Jühling F, Externbrink F, Florentz C, Fritzsch G, Pütz J, Middendorf M, Stadler PF. 2013. MITOS: improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 69(2):313–319.
- Chen IS. 2013. The complete mitochondrial genome of Chinese sucker Myxocyprinus asiaticus (Cypriniformes, Catostomidae). Mitochondrial DNA. 24(6):680–682.
- Chiang TY, Chen IS, Lin HD, Chang WB, Ju YM. 2013. Complete mitochondrial genome of Sicyopterus japonicus (Perciformes, Gobiidae). Mitochondrial DNA. 24(3):191–193.
- Crow KD, Kanamoto Z, Bernardi G. 2004. Molecular phylogeny of the hexagrammid fishes using a multi-locus approach. Mol Phylogenet Evol. 32(3):986–997.
- Cui Z, Liu Y, Li CP, You F, Chu HK. 2009. The complete mitochondrial genome of the large yellow croaker, Larimichthys crocea (Perciformes, Sciaenidae): unusual features of its control region and the phylogenetic position of the Sciaenidae. Gene. 432(1–2):33–43.
- Dierckxsens N, Mardulyn P, Smits G. 2017. NOVOPlasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 45(4):e18–e18.
- Gilbert MTP, Tomsho LP, Rendulic S, Packard M, Drautz DI, Sher A, Tikhonov A, Dalen L, Kuznetsova T, Kosintsev P, et al. 2007. Whole genome shot gun sequencing of mitochondria from ancient hair shafts. Science. 317(5846):1927–1930.
- Iwasaki W, Fukunaga T, Isagozawa R, Yamada K, Maeda Y, Satoh TP, Sado T, Mabuchi K, Takeshima H, Miya M, et al. 2013. MitoFish and MitoAnnotator: a mitochondrial genome database of fish with an accurate and automatic annotation pipeline. Mol Biol Evol. 30(11):2531–2540.
- Kanamoto Z. 1979. On the ecology of hexagrammid fish IV: mode of the distribution of Agrammus agrammus (Temminck et Schlegel) and Hexagrammos otakii Jordan et Starks and composition, abundance and food items of reef fish around the several reefs. Jpn J Ecol. 29:171–183.
- Kimura MR, Yanagimoto T, Munehara H. 2007. Maternal identification of hybrid eggs in Hexageammos spp. by means of multiplex amplified product length polymorphism of mitochondrial DNA. Aquat Biol. 1:187–194.
- Kwak SN, Baeck GW, Klumpp DW. 2005. Comparative feeding ecology of two sympatric greenling species, Hexagrammos otakii and Hexagrammos agrammus in eelgrass Zostera marina beds. Environ Biol Fish. 74(2):129–140.
- Ojala D, Montoya J, Attardi G. 1981. tRNA punctuation model of RNA processing in human mitochondria. Nature. 290(5806):470–474.
- Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 30(12):2725–2729.
- Wang C, Chen Q, Lu G, Xu J, Yang Q, Li S. 2008. Complete mitochondrial genome of the grass carp (Teleostei, Cyprinidae, Gobioninae). Gene. 424(1-2):96–101.