
Abstract
The chloroplast (cp) genome sequence of Bruguiera gymnorrhiza was characterized. The cp genome length was 163,795 bp in length, with a GC content of 35.3%, containing a large single copy (LSC) of 90,830 bp, a small single copy (SSC) of 20,207 bp, and a pair of inverted repeats (IRs) of 26,379 bp. The genome contained 121 genes, including 84 protein-coding genes, 37 tRNA genes, and 8 rRNA genes. A phylogenetic analysis using cp genomes of mangroves and ecologically associated species resolved B. gymnorrhiza in Bruguiera with B. sexangula var. rhynchopetala. This complete chloroplast sequence offers a promising tool for further species identification and evolutionary studies of Bruguiera, as well as for mangroves.
Keywords:
Introduction
Mangroves are a diverse group of about 70 woody trees and shrubs that inhabit the coasts of tropical and subtropical regions (Ball Citation1988; Duke Citation1992). Despite the seemingly harsh environment, mangrove ecosystems are highly productive ecosystems with rates of primary production equal to those of tropical humid evergreen forests and coral reefs (Barr et al. Citation2010; Alongi Citation2014; Lu et al. Citation2017). Nowadays mangroves are threatened by climate changed-induced drought, as well as, relative sea level rise (Lovelock Citation2020; Saintilan et al. Citation2020). Mangrove forests have been severely degraded over the past half century (Blasco et al. Citation2001; Donato et al. Citation2011), injecting new urgency into understanding the genetic resources of mangroves.
Large-leafed mangrove Bruguiera gymnorrhiza is one of the most important and widespread mangrove species, and widely distributed from the eastern coast of Africa through Asia to subtropical Australia (Allen and Duke Citation2006). The wood of B. gymnorrhiza is widely used for structural components of traditional homes, while people also using B. gymnorrhiza for food, as well as for dyes and traditional medicines (Allen and Duke Citation2006). Therefore, providing the genome sequences of species B. gymnorrhiza will help to spur research on these most interesting adaptations, and also could offer some needed information for its usage and conservation. Chloroplast DNA (cpDNA) have been proved could provide useful and abundant information on genetic diversity and evolution based on our previous studies (Jiang et al. Citation2016; Lei et al. Citation2018), as well as in mangroves (Chen et al. Citation2019; Yang et al. Citation2019; Shi et al. Citation2020). In this study, we assembled and characterized the chloroplast genome of B. gymnorrhiza based on Illumina pair-end data, and built a phylogenetic tree using plastomes available in mangroves and ecologically associated species.
B. gymnorrhiza was collected in Sanya Tielu Port mangrove reserve (Sanya, PR China, 18° 15′ N/109° 42′ E). The voucher (B. gymnorrhiza_Jiang_B7) is stored in Guangxi University, plant ecophysiology and evolution research group herbarium. Total genomic DNA was extracted from 0.1 g of frozen fresh leaves as previously described (Jiang et al. Citation2016; Lei et al. Citation2018; Xu et al. Citation2018). A 350-bp paired-end library was constructed and sequenced by Novogene (Beijing, PR China) using an Illumina HiSeqX-ten system (Illumina, San Diego, CA), about 1 Gb raw data filtered for read quality was obtained. We performed a de novo assembly using NOVOPlasty3.6 (Dierckxsens et al. Citation2016), seed sequence from mangrove Avicennia marina was retrieved from NCBI under the accession number (AB114520.1). The assembled sequence was then imported into Geneious R9 (Biomatters Ltd, Auckland, New Zealand), to check manually as described previously (Jiang et al. Citation2016; Hinsinger and Strijk Citation2017; Xu et al. Citation2017). The cp genome annotation was transferred from Rhizophora x lamarkii (NC_046517), a species from Rhizophoraceae. The final annotations were confirmed and integrated from results of CPGAVAS2 (Shi et al. Citation2019) and Chloroplot (Zheng et al. Citation2020).
The assembled cp genome of B. gymnorrhiza had a length of 163,795 bp (GenBank accession number MW402841). The cp genome exhibited the typical composition of LSC, SSC regions and two inverted repeats (IRa and IRb) of 90,830, 20,207, and 26,379 bp, respectively. The overall GC content of the plastome of B. gymnorrhiza was 35.3%, while the GC content in LSC, SSC, IRa, and IRb regions were 32.9%, 28.0%, 42.2%, respectively. We identified 121 genes, including 84 protein-coding genes, 37 tRNA genes, and 8 ribosomal RNA genes. Thirteen genes (atpF, petB, petD, rpoC1, trnK-UUU, trnL-UAA, trnT-CGU, trnV-UAC in LSC; ndhA locates in SSC; ndhB, rpl2, trnA-UGC, trnI-GAU in the IRs regions) contain 1 introns; while clpP, ycf3 in LSC, and rpS12 in IRb contain 2 introns, respectively. In total, 18 genes were duplicated in the IR regions, including 8 protein-coding genes (rps19, rpl2, rpl23, ycf1, ycf2, ndhB, rps7, rps12), 7 tRNA genes (trnA-UGC, trnI-GAU, trnI-CAU, trnL-CAA, trnN-GUU, trnR-ACG, trnV-GAC), and 4 rRNA genes (4.5S, 5S, 16S, 23S). Interestingly, B. gymnorrhiza had its own codon usage bias in comparison to Barringtonia racemosa from the analysis by Chloroplot (Zheng et al. Citation2020), especially large differences were found in psbK, ndhG, and rps7.
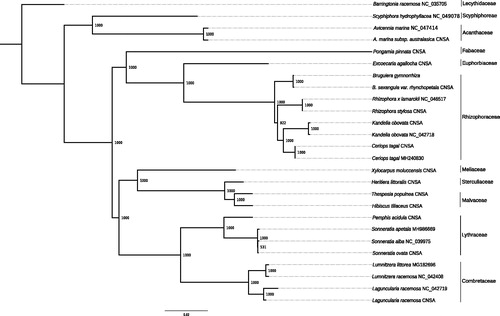
Twenty-four plastomes of mangroves and ecologically associated species were retrieved from GenBank (accessed 17 November 2020), and CNSA (https://db.cngb.org/cnsa/) according to the reference (Shi et al. Citation2020), plus Barringtonia racemosa as an out-group (). All the cps were aligned with MAFFT v7.307 (Katoh and Standley Citation2013), and a maximum likelihood (ML) tree was built using PhyML v3.3 (Guindon et al. Citation2009) with a GTR + I + G model and support estimated with 1000 bootstrap replicates. All but one node were highly supported (bootstrap support ≥92), with the two species of Bruguiera clustered together. The plastome of B. gymnorrhiza provides a useful bioresource that will help to assess population diversity for conservation purposes, and will also benefit to further genetic studies in Rhizophoraceae.
Figure 1. ML phylogeny of mangroves and ecologically associated species based on 25 cp genomes retrieved from the present study, GenBank and CNSA. The tree is rooted with Barringtonia racemosa. Bootstraps values (1000 replicates) are shown at the nodes. Scale in substitution per site.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability
The genome sequence data that support the findings of this study are openly available in GenBank of NCBI at (https://www.ncbi.nlm.nih.gov/) under the accession no. MW402841. The associated BioProject, SRA, and Bio-Sample numbers are PRJNA713533, SRR13933381, and SAMN18253765, respectively. Data were also available in the database: CNGB Sequence Archive (CNSA) of China National GeneBank DataBase (CNGBdb) with accession number CNP0001525 (https://db.cngb.org/search/project/CNP0001525/).
Additional information
Funding
References
- Allen J, Duke N. 2006. Bruguiera gymnorrhiza, large-leafed mangrove. Chapter in traditional trees of pacific islands: their culture, environment, and use: 139–152.
- Alongi DM. 2014. Carbon cycling and storage in mangrove forests. Ann Rev Mar Sci. 6:195–219.
- Ball MC. 1988. Ecophysiology of mangroves. Trees. 2(3):129–142.
- Barr JG, Engel V, Fuentes JD, Zieman JC, O'Halloran TL, Smith TJ, III, Anderson GH. 2010. Controls on mangrove forest-atmosphere carbon dioxide exchanges in western Everglades National Park. J Geophys Res-Biogeo. 115(G02020):1–14.
- Blasco F, Aizpuru M, Gers C. 2001. Depletion of the mangroves of Continental Asia. Wetl Ecol Manag. 9(3):255–266.
- Chen Y, Yang Y, Li J, Jin Y, Liu Q, Zhang Y. 2019. The complete chloroplast genome sequence of a medicinal mangrove tree Ceriops tagal and its phylogenetic analysis. Mitochondrial DNA Part B. 4(1):267–268.
- Dierckxsens N, Mardulyn P, Smits G. 2016. NOVOPlasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 45(4):e18–e18.
- Donato DC, Kauffman JB, Murdiyarso D, Kurnianto S, Stidham M, Kanninen M. 2011. Mangroves among the most carbon-rich forests in the tropics. Nature Geosci. 4(5):293–297.
- Duke NC. 1992. Mangrove floristics and biogeography. In: AIR, DMA, editors. Tropical mangrove ecosystems. Washington D.C.: American Geophysiocal Union; p. 63–100.
- Guindon S, Delsuc F, Dufayard J-F, Gascuel O. 2009. Estimating maximum likelihood phylogenies with PhyML. In: Posada D, editor. Bioinformatics for DNA sequence analysis. Totowa (NJ): Humana Press; p. 113–137.
- Hinsinger DD, Strijk JS. 2017. Toward phylogenomics of Lauraceae: the complete chloroplast genome sequence of Litsea glutinosa (Lauraceae), an invasive tree species on Indian and Pacific Ocean islands. Plant Gene. 9:71–79.
- Jiang GF, Hinsinger DD, Strijk JS. 2016. Comparison of intraspecific, interspecific and intergeneric chloroplast diversity in Cycads. Sci Rep. 6:31473.
- Katoh K, Standley DM. 2013. MAFFT multiple sequence alignment Software Version 7: improvements in performance and usability. Mol Biol Evol. 30(4):772–780.
- Lei JY, Hinsinger DD, Jiang GF. 2018. Characterization of the complete chloroplast genome of endangered Cycads Zamia fischeri Miq. ex Lem. Mitochondrial DNA B Resour. 3(2):1059–1061.
- Lovelock CE. 2020. Blue carbon from the past forecasts the future. Science. 368(6495):1050–1052.
- Lu W, Xiao J, Liu F, Zhang Y, Liu C, Lin G. 2017. Contrasting ecosystem CO2 fluxes of inland and coastal wetlands: a meta-analysis of eddy covariance data . Glob Chang Biol. 23(3):1180–1198.
- Saintilan N, Khan NS, Ashe E, Kelleway JJ, Rogers K, Woodroffe CD, Horton BP. 2020. Thresholds of mangrove survival under rapid sea level rise. Science. 368(6495):1118–1121.
- Shi C, Han K, Li L, Seim I, Lee SM-Y, Xu X, Yang H, Fan G, Liu X. 2020. Complete chloroplast genomes of 14 mangroves: phylogenetic and comparative genomic analyses. Biomed Res Int. 2020:8731857.
- Shi L, Chen H, Jiang M, Wang L, Wu X, Huang L, Liu C. 2019. CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res. 47(W1):W65–W73.
- Xu LM, Hinsinger DD, Jiang G-F. 2017. The complete mitochondrial genome of the Agrocybe aegerita, an edible mushroom. Mitochondrial DNA B Resour. 2(2):791–792.
- Xu LM, Hinsinger DD, Jiang G-F. 2018. The complete mitochondrial genome of the Basidiomycete fungus Pleurotus cornucopiae (Paulet) Rolland. Mitochondrial DNA B Resour. 3(1):73–75.
- Yang Y, Zhang Y, Chen Y, Gul J, Zhang J, Liu Q, Chen Q. 2019. Complete chloroplast genome sequence of the mangrove species Kandelia obovata and comparative analyses with related species. PeerJ. 7:e7713.
- Zheng S, Poczai P, Hyvonen J, Tang J, Amiryousefi A. 2020. Chloroplot: an online program for the versatile plotting of organelle genomes. Front Genet. 11:576124.