
ABSTRACT
We describe a new articulate Selaginella species (i.e. S. germinans) from the Chapada dos Veadeiros region in the State of Goiás, Brazil. This species is characterized by articulate stems, dorsal rhizophores, flagelliform stems and branch apices that form flattened buds or gemmae at their tips, as well as a single basal megasporangium on each strobilus, which is subtended by a megasporophyll and few other infertile sporophylls. We also discuss the taxonomic affinity, habitat, distribution, and conservation status of this new species, which is illustrated with digitized micrograph images of stem sections, leaves, and megaspores taken with a Scanning Electron Microscope (SEM). We propose that the formation of buds or gemmae in this new species is probably an adaptive vegetative reproductive mechanism to adverse natural or man-induced environmental conditions (e.g. fires and drought periods) in its habitat.
SUBJECTS:
Introduction
Molecular and morphological phylogenetic analyses have confirmed the monophyly of Selaginella (Zhou et al. Citation2015; Weststrand and Korall Citation2016a). Nevertheless, the infrageneric classification of the genus is unsettled and various classification systems have been proposed. The most recent and amply followed system based on morphological characters advanced by Jermy (Citation1986, Citation1990) recognized five subgenera (i.e. Selaginella, Tetragonostachys, Ericetorum, Heterostachys, and Stachygynandrum). The molecular and morphology-based infrageneric classification recently proposed by Zhou and Zhang (Citation2015) and Weststrand and Korall (Citation2016b) share some similarities but diverge in other aspects (Smith and Kessler Citation2018; Valdespino et al. Citation2018). Zhou and Zhang (Citation2015) recognized six subgenera (i.e. Selaginella, Boreoselaginella, Pulviniella, Ericetorum, Heterostachys, and Stachygynandrum), whereas Weststrand and Korall (Citation2016b) distinguished seven subgenera (i.e. Selaginella, Rupestrae, Lepidophyllae, Gymnogynum, Exaltatae, Ericetorum, and Stachygynandrum). All of these and previous classifications concur in recognizing as a natural alliance, at different taxonomic levels (see Zhou and Zhang Citation2015; Weststrand and Korall Citation2016b; for review), a group of species characterized by articulate stems (i.e. typically swollen or collapsed nodes, which are green, greenish-red or reddish shortly before each branching point; upon drying these nodes become somewhat darkened or even black). These articulate species have stems normally with one, two, or three vascular bundles (some taxa with a more complex stelar pattern), dorsal rhizophores, and a single megasporangium at the base of the strobili that usually only bears four megaspores and that is protected by several modified leaves (i.e. sporophylls). Somers (Citation1978, Citation1982) described a unique type of microsporangium in this group, one that is actively involved in a microspore ejection and dispersal mechanism that further morphologically defines this species alliance. This species group is commonly referred to as the “articulate” or “Articulatae” and includes about 50 species mostly from the New World tropics, with only two taxa found in the Old World; typically, species are found in exposed habitats. In the Americas, Venezuela with ca. 100 spp is the most species-rich country for Selaginella (Valdespino Citation2015, Citation2017a, Citation2017b), followed by Brazil with 85 previously recognized species. Based on very recent studies, the number of Brazilian Selaginella species is thought to be about 89–96 spp. (Valdespino et al. Citation2018) and among these, at least 20 correspond to the so-called “articulate” group. Herein, a most distinct articulate Selaginella species (i.e. S. germinans Valdespino & C. López) from the Chapada dos Veadeiros region in the State of Goiás, Brazil, is described as new. It was first recognized as a distinct taxon by the first author some years ago but description was dependant on revision of additional material, which became available during research at the herbarium of The New York Botanical Garden (NY).
Material & methods
This study is based on examination of morphological characters using an Olympus SZ 60-STS and a Olympus SZX16 stereomicroscopes at 0.7–11.5X of four Selaginella collections made in or around the Chapada dos Veadeiros region in the State of Goiás, Brazil, over the last 62 years, with the most recent gatherings made in 2011. In total, seven duplicates of these collections from K, NY, PMA, and US (herbarium acronyms according to Thiers Citation2018) were examined. Lateral, median, and axillary leaves and megaspore samples were removed for scanning electron microscopy (SEM) according to standard techniques as described by Valdespino (Citation1995) from designated type specimens. Samples were prepared, viewed, and imaged using a Zeiss Model Evo 40vp SEM at 15–20 kV at the Smithsonian Tropical Research Institute (STRI) in Panama. Raw SEM images were later post-processed and assembled in a multipart figure using Adobe Photoshop as explained in Valdespino (Citation2016). Terminology, measurements, and conservation status provided in the taxon description follows Valdespino (Citation2017a; Valdespino et al. Citation2018; and references therein).
Taxonomic treatment
Selaginella germinans Valdespino & C. López, sp. nov. ()
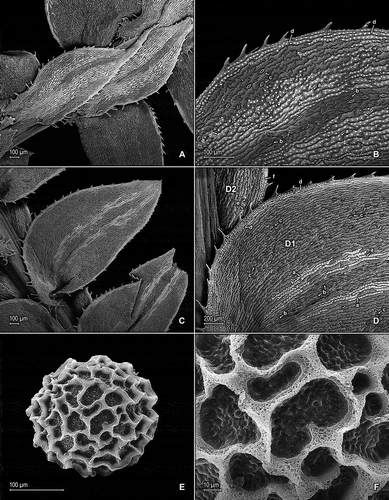
Figure 1. Selaginella germinans Valdespino & C. López. A. Section of upper surface of stem branch showing median and lateral leaves. B. Close-up of median leaf from main stem, upper surface; note, elongate and papillate idioblasts on both sides of the midrib (a), stomata along midrib (b) and on submarginal portion of lamina (c), and marginal, elongate and papillate idioblasts (d). C. Section of lower surface of main stem showing lateral leaves and portion of outer halves of median leaves. D. Close-up of proximal-acroscopic portion of lateral leaf (D1) and outer halve of median leaves (D2), lower surfaces; note, elongate and papillate idioblasts on both sides of the midrib (a), stomata along midrib (b) and on submarginal to submedial portions of lamina (c), and marginal, elongate and papillate idioblasts (d) on lateral leaf and submarginal stomata (e) and marginal, elongate and papillate idioblasts (f) on outer halve of median leaf. E. Megaspore, distal face; note close reticulate sculpturing pattern with high muri. F. Close-up of megaspore, distal face (same megaspore as in E); note high muri with perforate, sponge-like microstructure and rugulate and perforate microstructure on each reticulum lumen. A–F taken from holotype, Anderson et al. 7435 (holotype: NY).
Diagnosis
Selaginella germinans differs from S. sulcata (Desv.) Spring ex Mart. by its ovate (vs. oblong) lateral leaves; stomata of lateral leaves along midribs submarginal and irregularly arranged (vs. arranged in defined rows) along both sides of the midribs; lateral leaves with a single acroscopic auricle (vs. with both an acroscopic and a basiscopic auricle); acroscopic auricles 1/10 or 1/11 (vs. ⅕–1/8) the length of the lamina, curving inward (vs. straight or slightly curving inward), short-ciliate (vs. dentate); acroscopic margins along proximal ⅓ short-ciliate (vs. dentate); lower surfaces of lateral and axillary leaves with (vs. without) two distinct bands of idioblast-like cells on both sides and parallel to midribs, each band comprising three to four rows of idioblast-like cells (vs. idioblast-like cells when present few and scattered); and median leaves with a single auricle (vs. biauricled).
Type
BRAZIL. Goiás: Chapada dos Veadeiros, 20 km by road S of Terezina, 1200 m, 18 March 1973, W.R. Anderson et al. 7435 (holotype: NY!; isotypes: K!, PMA!).
Description
Plants terrestrial. Stems creeping to decumbent, stramineous, 10−17 cm long, 0.6–0.8 mm diam., inconspicuously articulate, flagelliform, stoloniferous, 2-branched. Rhizophores dorsal, restricted to bases of the stems, filiform, 0.2−0.4 mm diam. Leaves dimorphic throughout, chartaceous, both surfaces glabrous, the upper surfaces green to stramineous (when old), comprising elongate or puzzle-like cells with sinuate walls and hyaline, elongate, and papillate cells along the margins and along both sides of the midribs, the lower surfaces pale green to stramineous (when old) and comprising elongate cells with sinuate walls. Lateral leaves distant or imbricate distally, patent, ovate, 1.8−2.1 × 1.0−1.3 mm; bases asymmetric, the acroscopic bases overlapping the stems, with short-ciliate auricles, the basiscopic bases free from the stems, without auricles; the acroscopic margins hyaline and comprising a band one to five cells wide, each cell elongate and papillate, the papillae in one row, short-ciliate along proximal ¼ and serrulate along distal ¾, basiscopic margins greenish to hyaline and comprising a band one to three cells wide with each cell as in acroscopic margins, short-ciliate along proximal ⅕ and serrulate distally; apices acute, entire or tipped by one to three tooth-like projections; upper surfaces without conspicuous idioblasts, without stomata, lower surfaces with conspicuous, papillate idioblast-like cells and with stomata arranged in three or four rows along midribs and submedial to submarginal on both halves of each leaf lamina, the idioblast-like cells in two distinct bands on both sides and parallel to midribs, each band comprising three to four rows of idioblast-like cells, the papillae in one or two rows on each cell lumen, the stomata along the lamina length. Median leaves imbricate, peltate, ascending, ovate-elliptic to ovate-lanceolate, 1.5−2 × 0.8−1 mm; bases asymmetric and auricled, the auricles ciliate with an inner, incipient, small and cuneate lobe and an outer, well-developed, long, rectangular lobe; margins hyaline and comprising a band one to three cells wide, each cell elongate and papillate, the papillae in one row, short-ciliate along proximal ⅓−½, otherwise denticulate to serrate distally; apices aristate, the aristae with margins denticulate and with upper surfaces puberulent with tooth-like projections and stomata along proximal ¼, each arista ½ or less the length of the leaf lamina; upper surfaces with idioblast-like cells and stomata in one or two rows each along midribs and submargins, the idioblast-like cells papillate, arranged in 8 to 10 rows parallel to both sides of the midribs, the papillae in one or occasionally two rows, the stomata along distal ⅔ of the midrib of the lamina and submarginal to submedial along proximal ½ of the lamina, the lower surfaces without idioblasts and with stomata along proximal ½. Axillary leaves with shape, margins, and apices similar to lateral leaves; the bases biauriculate with auricles short-ciliate and incurved. Strobili terminal on branch tips or lateral, compact, quadrangular, 3.0−5.0 mm long. Sporophylls mostly monomorphic except for those protecting the megasporangia which are enlarged, without a laminar flap, ovate to ovate-elliptic, 0.9−1.1 × 0.5−0.7 mm, with a slightly developed keel along distal ½ of the midribs; bases rounded; margins narrowly hyaline, serrate; apices attenuate to acuminate and tipped by one or two tooth-like projections; dorsal sporophylls with both surfaces lacking idioblasts; upper surfaces pale-green, lower surfaces silvery-green; ventral sporophylls with both surfaces hyaline to silvery-green. Megasporangia one per strobili, on basal portion of ventral sporophylls; megaspores cream to pale yellow (some malformed and seemingly abortive), strongly reticulate with high muri on both faces, each reticulum lumen with rugulate and perforate (sponge-like) microstructure, ca. 250−260 μm. Microsporangia in ventral and dorsal rows throughout strobili; microspores pale yellow, not measured.
Habitat and distribution
Selaginella germinans grows on sandy soil, sandstone outcrops along river banks in Cerrado (tropical savanna) vegetation and mesophytic forest at 1040–1400 m; it is known only from the Chapada dos Veadeiros region in the State of Goiás, Brazil.
Conservation status
The Chapada dos Veadeiros region is part of the central Brazilian Plateau in the State of Goiás with a landscape ranging in elevations from 800 to 1650 m that typically harbors savanna vegetation (Sebastiani et al. Citation2015; da Silva, Pscheidt, and Cordeiro Citation2016; and references cited therein for a more complete phytophysionomic description of this area). This region is one of the most important remnants of the Cerrado vegetation in Brazil and has a rich and highly endemic flora (Mendoza, Simon, and Cavalcanti Citation2015; Sebastiani et al. Citation2015). Most of the plant diversity of the region, including the few known specimens of S. germinans, is confined to Chapada dos Veadeiros National Park (Sebastiani et al. Citation2015). Unfortunately, this region is currently threatened by agricultural expansion, natural and human-induced fires, commercial mining of minerals, and extraction of plants for craft works (Sebastiani et al. Citation2015), as well as tourism. Therefore, we consider S. germinans Critically Endangered (CR), according to the IUCN (Citation2012) categories and criteria.
Additional specimens examined (paratypes)
BRAZIL. Goiás: Chapada dos Veadeiros, 6−7 km E of Alto do Paraíso on road to Nova Roma, 1400 m, 7 March 1973, Anderson et al. 6548 (K, NY), 14°30ʹS, 47°30ʹW, 30 April 1956, Dawson 14,767 (US), Alto Paraíso de Goiás, road from Alto Paraíso de Goiás to Nova Roma (GO 239), ca. 1.5 km E of Alto Paraíso de Goiás, −14,116S, −47,499W [14°06ʹ57.6ʹʹS, 47°29ʹ56.4ʹʹW], 1040 m, 17 January 2011, Labiak & Mickel 5282 (NY, PMA, UPCB-n.v.).
Etymology
The specific epithet derives from the Latin “germen” meaning “sprout” and refers to the apical buds or gemmae on flagelliform stem and branch apices.
Notes
Selaginella germinans is a creeping species characterized by articulate stems with flagelliform projections on main stems and branch apices, which frequently are tipped by somewhat flattened buds or gemmae, peltate and auriculate median leaf bases with ciliate margins, ciliate and biauriculate axillary leaf bases with the auricles straight to slightly incurved, and acroscopic lateral leaf bases auricled with the auricles projecting downward and incurved with their lower surfaces comprising elongate and papillate cells, as well as a single basal megasporangium on each strobilus and subtended by a megasporophyll and few other infertile sporophylls. The articulate stems, dorsal rhizophores, a single basal megasporangium protected by modified sporophylls, and the unique type of microsporangia, as described by Somers (Citation1978, Citation1982), allow placement of S. germinans within subg. Gymnogynum following Weststrand and Korall (Citation2016b), subg. Ericetorum sect. Articulatae according to Zhou and Zhang (Citation2015), or subg. Stachygynandrum ser. Articulatae sensu Jermy (Citation1986, Citation1990) and Hieronymus (Citation[1901] 1902). Furthermore, we propose that the somewhat flattened, apical buds or gemmae on stems and branch apices found in this new species most probably are an adaptive vegetative reproductive mechanism to withstand adverse natural or human-induced environmental conditions such as fire and periods of drought in its habitat. Other plants growing in the same kind of vegetation as S. germinans develop a similar survival mechanism. For example, Genlisea tuberosa Rivadavia (Lentibulariaceae) forms subterraneous tubers during the dry, winter season to endure until the wet season, when it forms fully developed rosettes above ground and is able to set flowers (Rivadavia, Gonella, and Fleischmann Citation2013). In Selaginella, production of similar buds or gemmae as a vegetative reproductive strategy is not uncommon and can be found in some non-articulate species, e.g. S. moellendorffii Hieron., from the Old World (China, Japan, Philippines, Taiwan, and Vietnam) and S. porphyrospora A.Braun in the New World (Mexico, Guatemala, Honduras, Salvador, Nicaragua, Costa Rica, Panama, Colombia, Venezuela, and Ecuador), where these buds are swollen. However, the most similar comparable flattened buds or gemmae as reported here for S. germinans are found in other neotropical articulate species such as S. horizontalis (C.Presl.) Spring, S. schizobasis Baker, S. sertata Spring, and S. sulcata sensu lato, which together form the S. horizontalis complex. The last four taxa are typically well adapted to dry and sun-exposed conditions, such as is most probably the case for S. germinans.
Selaginella germinans is morphologically similar to and may be confused with S. sulcata because of its lateral leaves with stomata arranged in three or fourrows along midribs and hyaline, elongate, and papillate idioblast-like cells along margins, and median leaves peltate and auriculate with the upper surfaces comprising elongate, idioblast-like and papillate cells parallel to the midribs, as well as stomata submarginal and along the midribs extending into the aristate apices, each apex tipped by short, tooth-like projections, and biauricled axillary leaves. Nevertheless, the characters contrasted in the diagnosis separate these two species. In addition, S. germinans can be distinguished from S. sulcata by stem branches always with (vs. without or occasionally with) flagelliform apices, rhizophores restricted to proximal ½ (vs. throughout or along proximal ½) of the stems, stramineous (vs. frequently brownish) stems that are 2- (vs. 3- or 4-) branched, lateral leaves 1.8−2.1 (vs. 3.0−8.0) mm long, the median leaf idioblast-like cells with one or occasionally two (vs. with two) rows of papillae on each cell lumen, ovate (vs. oblong to oblong-lanceolate) axillary leaves, short (vs. large) strobili, each 3.0−5.0 (vs. 6.0−20) mm long, and reticulate megaspores 250−260 (vs. 630−650) μm diam with rugulate and perforate (i.e. sponge-like) microstructure on each lumen and muri (vs. microstructure on each cell lumen granular and mesh-like on muri). Selaginella germinans is further distinguished from S. sulcata by its median leaf with 8–10 (vs. 14–30) rows of elongate and papillate idioblast-like cells, parallel to midribs on upper surfaces, peltate bases each with a single auricle (vs. biauricled), the auricle straight and flap-like with a large outer rectangular lobe and an incipient inner, small, and cuneate lobe (vs. auricles divergent, the outer auricle large, rectangular, and skirt-like and the inner auricle smaller and usually cuneate), the apices each with an arista ½ or less (vs. ½–¾ or almost equal) the length of the leaf lamina and with stomata along proximal ¼ (vs. along proximal ⅛) on upper surfaces; and axillary leaves with two incurved (vs. straight and parallel to, or divergent from stems or occasionally slightly incurved) auricles.
Among other Brazilian Selaginella species, S. germinans may be confused with S. flagellata Spring because of their somewhat similar habit, rhizophores restricted to the base of the stems, and flagelliform stem and branch apices. Nevertheless, S. germinans is easily recognized from S. flagellata by its creeping to decumbent (vs. suberect to erect) and articulate (vs. non-articulate) stems, dorsal (vs. axillary) rhizophores, auricled (vs. not auriculate) leaf bases, and each strobilus with a single basal megasporangium (vs. with several megasporangia in two ventral rows).
As earlier discussed, S. germinans is so far only known from the Chapada dos Veadeiros region in the State of State of Goiás, Brazil. This region is one of the most important remnants of the Cerrado vegetation in that country and has a rich and highly endemic flora (Mendoza, Simon, and Cavalcanti Citation2015; Sebastiani et al. Citation2015). Nevertheless, Morton (Citation1960) stated that the pteridophyte (i.e. ferns and lycophytes) flora of the State of Goiás was not rich and he further determined one of the paratypes (i.e. Dawson 14767, US!) as Selaginella sp. Morton’s species-richness characterization may have been premature and due, in part, to the lack of critical collecting of plants and particularly of ferns and lycophytes at that time. We now know, for example, that in the case of Selaginella, at least 12 species, including S. germinans, are found in the State of Goiás (Valdespino Citation2015; Valdespino et al. Citation2015; Flora do Brasil 2020 under construction Citation2018), and this total represents ca. 13% of all selaginellas so far known in Brazil. Moreover, five of those species (i.e. S. cabrerensis Hieron., S. germinans, S. minima Spring, S. simplex Baker, and S. vestiens Baker) are typically found only in Goiás and/or in nearby states, whereas additional seven other taxa [i.e. S. convoluta (Arn.) Spring, S. erectifolia Spring, S. erythropus (Mart.) Spring, S. flexuosa Spring, S. marginata (Humb. & Bonpl. ex Willd.) Spring s.l., S. saltuicola Valdespino, and S. sellowii Hieron.] are parts of the floristic element of the Central Brazilian Savanna phytogeographic domain (Valdespino Citation2015; Valdespino et al. Citation2015; Flora do Brasil 2020 under construction Citation2018). Furthermore, Mendoza, Simon, and Cavalcanti (Citation2015) considered the overall number of plant species reported for the Chapada dos Veadeiros region to be underestimated based on the number of new taxa recently described for this site and because it still has large, botanically unexplored areas due to difficult access. Therefore, it is reasonable to expect that as a more concerted collecting effort is undertaken in the Chapada dos Veadeiros region, along with a critical revision of Selaginella specimens already collected there, additional known and unknown taxa will eventually be uncovered in this area, as was the case for S. germinans.
Acknowledgments
We are grateful to curators and staff of herbaria cited for allowing study of their Selaginella specimens. We are thankful to the New York Botanical Garden, the offices of the dean of the Faculty of Natural and Exact Sciences and Technology and the Vice-president for Research and Graduate Programs of the University of Panama, the Smithsonian Tropical Research Institute in Panama, and the National Research System of Panama (SNI) for support provided for this research. Our gratitude also extends to Christopher Gioia for revision and checking the English draft of this paper and to Pedro Argudo of NovoArt for help preparing illustration. IAV also is grateful to Chris for his continuous unwavering encouragement. Last, we are indebted to subject editor of Botany Letters and Alan R. Smith for their suggestions to improve the manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Notes on contributors
Iván A. Valdespino
Iván A. Valdespino holds a Ph.D. from The New York Botanical Garden and The City University of New York, New York joint doctoral program and was a postdoctoral fellow at the California Academy of Sciences, San Francisco, California. His research interests centers on ferns and lycophytes and he is recognized as a World authority on the taxonomy and systematics of Selaginella, particularly from the Neotropics. Currently, he is a professor at the Botany Department of the University of Panama and a fellow of the National Research System (SNI) of Panama.Contribution: conceptualized and carried out the study, including SEM work, wrote and revised the manuscript.
Christian A. López
Christian A. López is an undergraduate student at the Botany Department of the University of Panama where he is conducting studies on the taxonomy and systematics of the articulate Selaginella species under the direction of IAV and Alejandra Vasco (now at BRIT).Contribution: carried out SEM work and co-wrote and revised the manuscript.
Jorge Ceballos
Jorge Ceballos obtained his bachelor degree from the Botany Department of the University of Panama and works at the Microscopy Laboratory of the Smithsonian Tropical Research Institute (Panama).Contribution: carried out SEM and assisted in writing the manuscript.
References
- da Silva, M. J., A. C. Pscheidt, and I. Cordeiro. 2016. “Microstachys crassifolia sp. nov. (Euphorbiaceae) from Chapada dos Veadeiros, Goiás, Brazil.” Nordic Journal of Botany 34: 364–369. doi:10.1111/njb.00896.
- Flora do Brasil 2020 under construction. 2018. “Jardim Botânico Do Rio De Janeiro.” Accessed March 20, 2018. http://www.floradobrasil.jbrj.gov.br/.
- Hieronymus, G. [1901] 1902. “Selaginellaceae.” In Die Natürlichen Pflanzenfamilien, 1(4), edited by A. Engler and K. Prantl, 621–716. Leipzig: W. Engelmann.
- IUCN (International Union for Conservation of Nature). 2012. IUCN Red List Categories and Criteria, Version 3.1. Second Edition. Prepared by the IUCN Species Survival Commission. Gland, Switzerland and Cambridge, United Kingdom: IUCN.
- Jermy, A. C. 1986. “Subgeneric Names in Selaginella.” Fern Gazette 13 (2): 117–118.
- Jermy, A. C. 1990. “Selaginellaceae.” In Pteridophytes and Gymnosperms, the Families and Genera of Vascular Plants I, edited by K. U. Kramer and P. S. Green, 39–45. New York: Springer-Verlag.
- Mendoza, J. M., M. F. Simon, and T. B. Cavalcanti. 2015. “Three New Endemic Species of Manihot (Euphorbiaceae) from the Chapada dos Veadeiros, Brazil.” Arnaldoa 22 (3): 297–312. doi:10.11646/phytotaxa.273.3.1.
- Morton, C. V. 1960. “The Machris Brazilian Expedition. Botany: Pteridophyta.” Los Angeles County Museum Contributions in Science 35: 1–8.
- Rivadavia, F., P. M. Gonella, and A. Fleischmann. 2013. “A New and Tuberous Species of Gensiela (Lentibulariaceae) from the Campos Rupestres of Brazil.” Systematic Botany 38 (2): 464–470. doi:10.1600/036364413X666679.
- Sebastiani, R., A. Francener, C. Da Vieira, and R. F. Da Almeida. 2015. “An Illustrated Checklist of Malpighiaceae from the Chapada dos Veadeiros Region, Goiás, Brazil.” Check List 11 (6): 1801. doi:10.15560/11.6.1801.
- Smith, A. R., and M. Kessler. 2018. “Prodromus of a Fern Flora of Bolivia. III. Selaginellaceae.” Phytotaxa 344 (3): 248–258. doi:10.1164/phytotaxa.344.3.5.
- Somers, P. 1978. “A systematic survey of the Articulatae series of the genus Selaginella and monographic treatment of the S. sulcata group (sensu str.).” Ph.D diss., The University of Tennessee, Knoxville.
- Somers, P. 1982. “A Unique Type of Microsporangium in Selaginella, Series Articulatae.” American Fern Journal 72 (3): 88–92. doi:10.2307/1546603.
- Thiers, B. 2018. “Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium.” Accessed March 20, 2018. http://www.sweetgum.nybg.org/ih/.
- Valdespino, I. A. 1995. “A monographic revision of Selaginella P. Beauv. subgenus Heterostachys Baker in Central and South America.” Ph.D diss., The City University of New York, New York.
- Valdespino, I. A. 2015. “Novelties in Selaginella (Selaginellaceae – Lycopodiophyta), with Emphasis on Brazilian Species.” PhytoKeys 57: 93–133. doi:10.3897/phytokeys.57.6489.
- Valdespino, I. A. 2016. “Selaginella aculeatifolia sp. nov. (Selaginellaceae– Lycopodiophyta) from the Foothills of Cerro Neblina, Venezuela.” Nordic Journal of Botany 34: 370–375. doi:10.1111/njb.00954.
- Valdespino, I. A. 2017a. “Novel Fern- and Centipede-Like Selaginella (Selaginellaceae) Species and a New Combination from South America.” PhytoKeys 91: 13–38. doi:10.3897/phytokeys.91.21417.
- Valdespino, I. A. 2017b. “Selaginella hyalogramma (Selaguinellaceae – Lycopodiophyta): A New Species from Venezuela, South America.” American Fern Journal 107 (2): 72–83. doi:10.1640/0002-8444-107.2.72.
- Valdespino, I. A., G. Heringer, A. Salino, L. A. A. Góes-Neto, and J. Ceballos. 2015. “Seven New Species of Selaginella Subg. Stachygynandrum (Selaginellaceae) from Brazil and New Synonyms for the Genus.” PhytoKeys 50: 61–99. doi:10.3897/phytokeys.50.4873.
- Valdespino, I. A., C. A. López, A. M. Sierra, and J. Ceballos. 2018. “From the Guiana Highlands to the Brazilian Atlantic Rain Forest: Four New Species of Selaginella (Selaginellaceae – Lycopodiophyta: S. agioneuma, S. magnafornensis, S. ventricosa, and S. zartmanii).” Peer Journal 6: e4708. doi:10.7717/peerj.4708.
- Weststrand, S., and P. Korall. 2016a. “Phylogeny of Selaginellaceae: There Is Value in Morphology after All.” American Journal of Botany 103 (12): 2136–2159. doi:10.3732/ajb.1600156.
- Weststrand, S., and P. Korall. 2016b. “A Subgeneric Classification of Selaginella (Selaginellaceae).” American Journal of Botany 103 (12): 2160–2169. doi:10.3732/ajb.1600288.
- Zhou, X. M., C. J. Rothfels, L. B. Zhang, Z. R. He, T. Le Péchon, H. He, N. T. Lu, et al. 2015. “A Large-Scale Phylogeny of the Lycophyte Genus Selaginella (Selaginellaceae: Lycopodiopsida) Based on Plastid and Nuclear Loci.” Cladistics 32 (4): 360–389. doi:10.1111/cla.12136.
- Zhou, X. M., and L. B. Zhang. 2015. “A Classification of Selaginella (Selaginellaceae) Based on Molecular (Chloroplast and Nuclear), Macromorphological, and Spore Features.” Taxon 64 (6): 1117–1140. doi:10.12705/646.2.