
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
In Ethiopia, most of the studies on mammals have focused on single species and protected areas. This survey investigated the impacts of habitats and seasons on the medium and large-sized mammals’ diversity and distribution in a human-dominated landscape. This survey was conducted using 36 systematically distributed transect lines within stratified habitat types. A total of 685 records belonging to 21 species, six orders, and 13 families were identified, including globally vulnerable Panthera leo, Panthera pardus, and Hippopotamus amphibious. Overall, Papio anubis and Chlorocebus aethiops were the dominant species recorded in the area. Except for three species, all species encountered in the three habitats are subsets of the species recorded in the grassland. The species record frequency was highest in the forest. The dry season characterized by a higher frequency of records but by lower species richness than the wet season. In general, the study area is potential for mammalian species conservation in Ethiopia.
Introduction
Mammalian species act as umbrella species of terrestrial ecosystems since they contribute to the conservation endeavors of other species (Bene et al., Citation2013; Udy et al., Citation2021) and they keep up ecosystem balance. Medium and large-sized mammalian species play key roles throughout many of the world’s ecosystems, including grazing, predation and seed dispersal (Gebresenbet et al., Citation2018; IUCN, Citation2021; Reeder et al., Citation2019). Moreover, they provide important human benefits such as food, recreation, and income (Penjor et al., Citation2021; Wolf & Ripple, Citation2018). Conversely, mammalian species have been in an extinction crisis globally and locally due to anthropogenic activities (Ripple et al., Citation2014; Worku & Girma, Citation2020). Habitat loss and degradation and harvesting (hunting/gathering for food, medicine, fuel, and materials) are by far the main threats to mammalian species (Bakala & Mekonen, Citation2020; Kasso & Bekele, Citation2017; Mekonen, Citation2020; Qufa & Bekele, Citation2019; Wale, Citation2017).
In addition, habitat and seasonal heterogeneity can decide the population patterns of wildlife, either a population decrease or increase. Hence, quantifying their diversity and distribution across habitats and seasons is pivotal for developing conservation strategies to avoid extermination and to secure the richness of mammalian biodiversity (Girma & Worku, Citation2020; Kasso & Bekele, Citation2017; Udy et al., Citation2021), like the area for the present study.
Worldwide Class Mammalia is composed of 5487 species. Of these, more than 1150, 360, 320 different mammalian species are found in Africa (Newbold et al., Citation2015), eastern Africa (Diriba et al., Citation2020; Girma et al., Citation2012) and Ethiopia (Amare, Citation2015; Lavrenchenko & Bekele, Citation2017; Tefera, Citation2011; Yalden et al., Citation1986), respectively. Among the recognized 320 mammalian species of Ethiopia, 55 are endemic and distributed in 14 orders and 39 families (Lavrenchenko & Bekele, Citation2017; Rabira et al., Citation2015). Furthermore, Ethiopia is one of the top 25 biodiversity-rich countries in the world and hosts two of the world’s 34 biodiversity hotspots. More than 60% of the mammalian species are medium and large-sized (Negeri et al., Citation2015). Topographic diversity and climate are the most significant predictors of mammalian species diversity in the country (Amare, Citation2015; Bakala & Mekonen, Citation2020; Belete & Melese, Citation2016; Tefera, Citation2011). However, the wildlife population has diminished both in abundance and distribution through the loss of habitat, hunting, and land clearance for farming; land degradation due to overgrazing (Gebresenbet et al., Citation2018; Girma & Worku, Citation2020; Lemma & Tekalign, Citation2020; Worku & Girma, Citation2020). To reverse the situation, research-based action is in need.
In Ethiopia, most of the studies on mammalian species were confined to protected areas (Fetene et al., Citation2019; Wale, Citation2017) but the diversity and distribution status of mammalian species outside protected areas such as human-dominated landscape are poorly known (Gebresenbet et al., Citation2018). Be that as it may, the study of mammalian species in communal areas is equally important (Girma & Worku, Citation2020; Lemma & Tekalign, Citation2020; Tamrat et al., Citation2020; Udy et al., Citation2021) even more because of the huge anthropogenic pressures (Burgin et al., Citation2018; Girma et al., Citation2012; Legese et al., Citation2019; Worku & Girma, Citation2020). There are some documented information on mammalian species of human-dominated landscapes in the northern, southwestern, southeastern, and central parts of Ethiopia (Bakala & Mekonen, Citation2020; Gebresenbet et al., Citation2018; Getachew & Mesele, Citation2018; Lavrenchenko & Bekele, Citation2017; Legese et al., Citation2019; Qufa & Bekele, Citation2019) and a few in Southern Ethiopia (Diriba et al., Citation2020; Girma et al., Citation2012; Lemma & Tekalign, Citation2020). There are several intact forests in the southern parts of Ethiopia. However, their fauna is still not well documented.
The present study was carried out in the Mirab Abaya district, Gamo Zone, Southern Ethiopia. The landscape is largely a forest habitat that harbors distinctive mammal species. The study area is associated with Lake Abaya, the largest lake in the Ethiopian Rift Valley system, which is the main water source for mammalian species and the lake created wetland habitat. In spite of this, the landscape has been under human obstructions (e.g., poaching, settlement, expansion of banana and vegetable plantations, firewood collection, and logging for charcoal production) and livestock pressure. Such human-induced actions can adversely affect the wildlife of the landscape. Understanding the distribution of prominent biological components such as mammalian species in the area is important to urgent management actions. Moreover, there is no ecological study on biodiversity undertaken in the area until now.
Therefore, to contribute towards closing these gaps and to supply the primary essential quantitative bits of knowledge, the present study examined the impacts of habitats and seasons on mammalian species composition, diversity and distribution using direct and indirect mammalian species evidence along line transect. The research questions surveyed were as follows: i) Are mammalian species composition and distribution vary between habitat types and seasons? It hypothesized that larger size forested habitat and the wet season would have more mammal species composition compared to agricultural areas and the dry season as more resources are available in the larger sized forest area and wet season; ii) does mammalian species diversity vary among habitats and seasons? It hypothesized that mammal species richness (number of species) and abundance (frequency of records) vary between wet and dry seasons and among habitats due to resource differences. The findings of the present study are crucial to justify the conservation status of mammalian species in different habitat types and seasons in the landscape.
Materials and methods
The study area
FFL is found in Mirab Abaya district in Gamo Zone, Southern Ethiopia, and lies between 06°10ʹ12” to 06°15ʹ00” N latitude and 37°42ʹ36” to 37°47ʹ24” E longitude (see ) and at about 475 km away from Addis Ababa, the capital city of Ethiopia. The study area is located 30 km north of Arba Minch town, the capital city of Gamo Zone. The total area of the FFL is around 100 km2. The FFL bounded by Fura Kebele (the lowest administrative unit in Ethiopia) to the south, Faragosa Kebele to the north, and Lake Abaya to the west and southwest. Fura and Faragosa Kebeles are settlement areas in the study landscape. There is one main asphalted road from Addis Ababa to Arba Minch that crosses the FFL makes it easily accessible.
Figure 1. Location of the study area and sampled area where transect line distributed among different habitats in the study area.
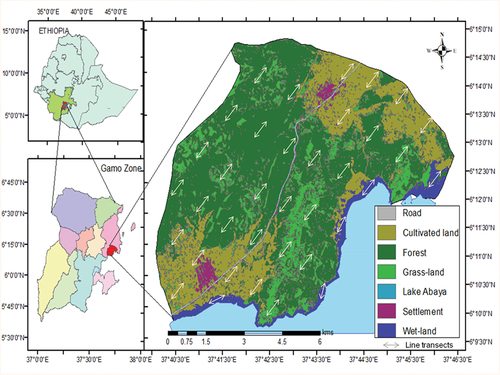
The altitudinal range of the FFL is 1,182–1,505 m a.s.l. The study area has a bimodal rainfall: June to September (heavy rains) and March to April (light rains), the remaining months of the year are fairly dry. The mean monthly rainfall and temperature in the area are 41.8–161.4 mm and 14.75–26.75°C, respectively (ENMSA, Citation2019).
FFL characterized by heterogeneous habitats. Thus, based on land-use types and water availability, the landscape divided into four habitat types: forest (area = 36.04 km2), wetland (area = 10.12 km2), grassland (area = 17.74 km2), and cultivated land (area = 24.19 km2) (see ). The habitats were determined using degrees on Google Earth map, ArcGIS, reconnaissance, and with the help of a GPS. Each habitat types were further divided into spatially isolated sites (wetland = 5, forest = 14, grassland = 9, cultivated land = 10) where the line transects lay. Based on transect reconnaissance, the common flora of the area consists of Terminalia brownie, Acacia spp, Dodonaea angustifolia, Acalypha fruticosa, Maytenus arbutifolia, Olea europaea, Ximenia americana, Syzygium guineense, Prunus africana, Bridelia scleroneura, Maytenus undata, Vangueria apiculata, Rhus vulgaris and Ozoroa insigns.
Data collection
Diurnal line‐transect is a well‐recognized and cost‐effective methodology for surveying medium and large vertebrates in tropical forests and savannas (Sutherland, Citation2006). It is one of the best methods for estimating abundance of relatively large and conspicuous mammals (Sutherland, Citation2006). So, combining diurnal line‐transect with indirect surveys (including fresh tracks, feces, hair, horns, burrows and digging) can enhance the detectability for many mammal species, contributing to maximize the species lists (Getachew & Mesele, Citation2018).
To collect mammalian data, a fixed-width line transect sampling method was used. Based on satellite images, ArcGIS and preliminary survey, the study area was first stratified into the four habitat types described above: forest, wetland, grassland, and cultivated land following Worku and Girma (Citation2020). Each habitat type further divided into spatially isolated sites as described above. This was delineated on a top map of the area, and transects were then established systematically in representative (homogenous vegetation) areas of each habitat type in spatially isolated sites. The distance between adjacent transects and from habitat edge to a transect was limited to a minimum of 0.5 km, to avoid double counting and to avoid edge impacts, respectively (Sutherland, Citation2006). A total of 36 line transects were established across the four major habitat types. The number of transects varied among habitats depending on the isolated sites: 14 in the forest, 5 in wetland, and 9 in grassland and 10 in cultivated land habitats. The length of each transect line was 2 km and a fixed sighting distance of 100 m on both sides of transects was used in the habitats. During data collection, the starting and ending points of each transect fed into a Garmin GPS unit and used for navigation.
Data collected in August and September 2019 during the wet season and January and February 2020 during the dry season. Mammal surveys carried out for two days per season and two times per day (early in the morning between 6:00, 10:00 hr and late in the afternoon between 15:00, 18:00 hr; when most animals are thought to be more active) (Belete & Melese, Citation2016; Woldegeorgis & Wube, Citation2012). Therefore, each transect line was surveyed eight times during the study period.
During transect visits, a researcher and five trained data collectors traversed the line transects. The data collectors were walking quietly and gently and at a constant speed along each transect against the direction of the wind to minimize disturbances of mammalian species. During data collection, a researcher and five trained field assistants traversed the transect lines. The observers were walking quietly and gently along each transect and at a constant speed along each transect against the direction of the wind to minimize disturbances of mammalian species.
Data were collected by recording animal observations and signs (fecal droppings, feed marks, tracks, burrows, territorial markings, spine sound, and other evidence) (Kingdong, Citation2015; Rabira et al., Citation2015). To avoid recounting of the same sign during subsequent monthly sampling periods, only the counted signs by data collectors and the researcher were marked at a place. Data collectors shifted in transects to minimize bias. Data recorded whenever an individual animal or group or signs of animals sighted were as follows: date, time, habitat type, species name, individual number of each species, and GPS location (Diriba et al., Citation2020; Girma et al., Citation2012; Rabira et al., Citation2015). Animal counting was made by the naked eye and using Bushnell laser rangefinder binoculars. Whenever deemed necessary, the field guide book was used for the identification of mammalian species (Kingdong, Citation2015). Only unambiguous signs recorded. Data from the four replicate surveys in seasons were pooled together for each transect and used for analysis (Diriba et al., Citation2020; Girma & Worku, Citation2020). Data collectors were used COVID 19 preventive measures such as social distancing and mouth mask throughout the data collection and compiling data.
Data analysis
Based on records of direct and indirect sign surveys along transects, a presence/absence data matrix was generated and processed for the study habitats and seasons. The recorded evidence of mammalian species classified to their respective orders, families, species level, and IUCN Red List categories. The conservation status of each species was also identified based on the IUCN Red List (IUCN, Citation2021). An individual-based rarefaction method was used to estimate species richness and abundance in habitat types and seasons (Legese et al., Citation2019; Qufa & Bekele, Citation2019). Similarities in species composition among habitats were presented using Venn diagram. The summed abundance of the number of encounters of each species recorded along each transect for each habitat type in each season was used as the input for the individual-based richness computation. Species richness, diversity, and evenness of mammalian species in the study area were analyzed by Shannon–Weaver diversity Index, Simpson’s diversity index, and evenness index, respectively, using the following formula (Sutherland, Citation2006):
Table
where H′ = Shannon–Weaver diversity Index, 1-D = Simpson diversity index, D = dominance, S = the number of species, Pi = the proportion of sampled species expressed as a proportion of the total sample, ln = logbasen.
Then, variations in the number of species belonging to each relative abundance category among habitats were tested using chi-square test. To examine the similarity of species composition among habitat types, Morisita–Horn index (sensitive to the abundance of the most abundant species) analysis was conducted between each pair of habitats (Sutherland, Citation2006). Habitat analysis performed using ArcGIS and all data analysis performed using Minitab software.
Results
Species composition and richness
A total of 21 mammalian species belonging to six orders and 13 families were identified in the FFL (see ). Order Carnivora was the first and the second most abundant order in terms of the number of families (five families) and species (six species), respectively. Order Artiodactyla was the second and the first most abundant in terms of the number of families (three families) and species (seven species), respectively. Four mammalian orders were represented each by a single species. At the family level, Bovidae (four species) and Canidae (three species) were the dominant families. The families Suidae, Felidae, and Sciuridae were represented each by two species, but eight families were represented each by a single species. Based on IUCN Red List categories, three species such as Hippopotamus amphibius, Panthera leo, and Panthera pardus were vulnerable species found in the study area. Tragelaphus imberbis, Redunca redunca, and Ourebia ourebi were categorized as lower risk/conservation dependent IUCN categories (see ). Out of a total of 21 species recorded, 20 species recorded during the wet and dry seasons, while Panthera pardus recorded only in the wet season (see ).
Table 1. List of mammalian species recorded in the FFL, their scientific names, common names, local names, and IUCN red list categories.
Species distribution
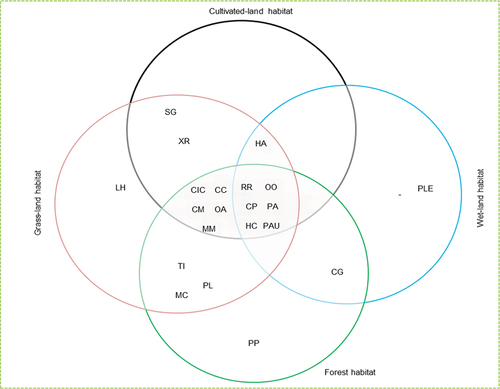
At the habitat level, mammal species richness and assemblage were varied among the four habitat types, in increasing order of wetland < cultivated land < forest < grassland (see ). Redunca redunca, Ourebia ourebi, Phacochoerus aethiopicus, Hystrix cristata, Papio anubis, and Chlorocebus aethiops were the six (23.81%) species that shared all habitat types in common (habitat generalists), while Lepus habessinicus, Panthera pardus and Panthera leo were habitat specialists recorded only in grassland, forest and wetland, respectively (see ). Except for Panthera leo, Panthera pardus, and Colobus guereza, All species encountered in the three habitats are subsets of the species recorded in the grassland.
Figure 2. The Venn diagram showing assemblage of mammalian species in habitat types.
Species relative abundance
A total of 685 pieces of evidence of mammalian species were recorded in the FFL. The number of records varied among orders and families. The abundant order by the number of records from the study area was order Primates which include 290, followed by order Artiodactyla including 194. The least abundant order was Tubulidentata, which composes only eight records. The most abundant family by the number of records was Cercopithecidae (290), whereas the least was Viverridae, comprising only three records. Based on the frequency of records, Papio anubis (20.15%) was the most abundant in the study area followed by Chlorocebus aethiops (19.27%). Based on IUCN Red List categories, the vulnerable species such as Panthera pardus and Panthera leo each contributed less than 0.29%, whereas Hippopotamus amphibius contributed 4.09% of the total records.
The results of the present study showed that of the 685 total records, 30.80% (N = 211) was recorded in the natural forest, 20% (N = 137) in the wetland, 29.64% (N = 203) in grassland and 19.56% (N = 134) in the cultivated land habitats. The number of records of mammalian species was varied significantly among habitats (KWχ2 = 6.03; P > 0.05).
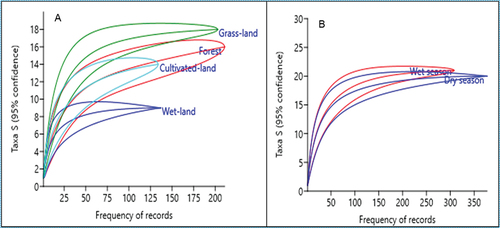
The mean number of species richness and relative abundance (frequency of records) computed by the rarefaction curve among the four stratified habitat types and seasons is represented by below.
Figure 3. Mean number of species richness and frequency of records computed by rarefaction curve among the four stratified habitat types (A) and seasons (B).
At species-specific level, Chlorocebus aethiops was most abundant species in forest habitat (33.18%, n = 70) and wetland (32.11%, n = 44) followed by Papio anubis (26.54%, n = 56) and Hippopotamus amphibius (20.43%, n = 28), respectively. Papio anubis (23.49%, n = 47) was also the most abundant in grassland followed by Phacochoerus aethiopicus (13.30%, n = 27) while, in cultivated land the most abundant was Xerus rutilus (29.14%, n = 39) followed by Crocuta crocuta (22.39%, n = 30). Panthera pardus and Panthera leo were only recorded in forest and wetland habitats, respectively. Mammalian species frequency of records among the four habitat types is described in below.
Figure 4. Mammalian species frequency of records among four habitat types.
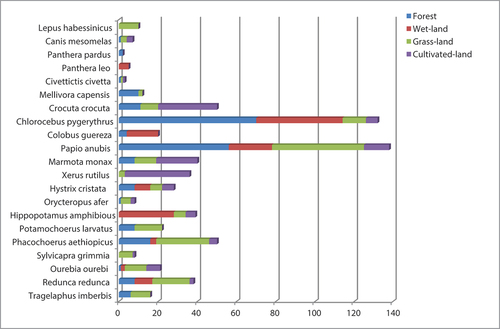
Figure 5. Mammalian species frequency of records in wet and dry seasons.
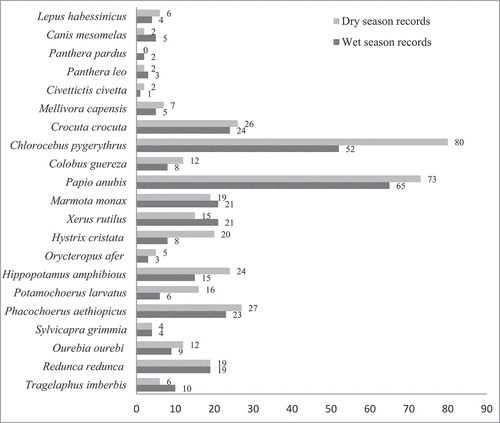
The number of records of mammalian species was higher in the dry season (n = 377, 55.04%) than in the wet season (n = 308, 44.96%). The abundance of mammalian species varied significantly between seasons (χ2 = 40.783; df = 20; P < 0.05). Two species (Papio anubis and Chlorocebus aethiops) were relatively the most abundant in both seasons (see ). These two species contributed 37.99% and 40.58% of the total records of the wet and dry season survey, respectively. The remaining mammalian records contributed between 0.32 and 7.79% in the wet season and 0.53 and 6.90% during the dry season survey. Frequency of records across habitat types was significantly different (χ2 = 43.147; df = 20; P < 0.05) between seasons.
Species diversity and similarity indices
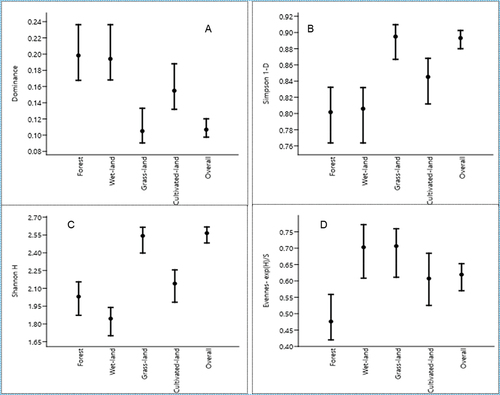
The Shannon diversity of mammal species was higher in the grassland (H = 2.543) than in other habitats. However, there was no significant difference in Shannon – Wiener Index values between the four habitat types. The higher and lower evenness of the mammalian species was recorded in grassland (E = 0.7064) and natural forest (E = 0.4761). The dominance of mammalian species was recorded from the highest to the lowest in the natural forest (D = 0.1983) and grassland (D = 0.1051), respectively. Shannon diversity index and dominance of mammalian species were similar during the dry and wet seasons. The overall species richness of FFL was 21 and Shannon–Wiener Index value (H) was 2.56, and Simpson’s index of diversity showed the highest species diversity (0.8968) in the study area. Among the four habitats, more similarity of mammalian species was observed between forest and grassland (0.609) and the least similarity observed wetland vs. cultivated land (0.273). The diversity indices and similarity of mammalian species across habitats are described in .
Figure 6. Error bars of mammalian species diversity indices across habitat types.
Discussion
Species taxonomic composition
The orders and families of mammalian species recorded in the present study were higher than in the study conducted on medium and large-sized mammals in different localities. For instance, Legese et al. (Citation2019) identified five orders and seven families in the Wabe forest, Ethiopia. Also, Qufa and Bekele (Citation2019) identified seven orders and 11 families from the Lebu Natural Protected Forest, Southwest Showa, Ethiopia; Lemma and Tekalign (Citation2020) recorded four orders and five families in the Humbo Community-Based Forest Area, Southern Ethiopia; herein FFL 6 orders and 13 families were recorded. On the contrary, Girma and Worku (Citation2020) identified nine families and five orders in the Nensebo Forest, Southern Ethiopia, which is higher than the present study.
The Primates were the most abundant order recorded and all belong to the family Cercopithecidae. Similarly, several studies have also reported a higher relative abundance of Primates than other orders from different parts of Ethiopia (Bakala & Mekonen, Citation2020; Belete & Melese, Citation2016; Rabira et al., Citation2015; Worku & Girma, Citation2020). This is could be due to the high reproductive successes, their more adaptive nature to different habitats, diversified foraging behavior, and high tolerance level of Primates to human disturbances (Lemma & Tekalign, Citation2020; Negeri et al., Citation2015). Order Carnivora contained the highest number of families (4) among other orders. The result is consistent with the different studies elsewhere in Ethiopia (Bakala & Mekonen, Citation2020; Girma & Worku, Citation2020; Lemma & Tekalign, Citation2020; Rabira et al., Citation2015). Order Artiodactyla has the highest species richness and the second abundant order recorded. This is in agreement with the study in the Nensebo forest in Southern Ethiopia where Artiodactyla was the most abundant order containing more species (Girma & Worku, Citation2020; Lemma & Tekalign, Citation2020). Orders such as Rodentia, Tubulidentata, and Lagomorpha were recorded as less in the number of individuals. This is in line with other studies in different localities in Ethiopia (Fetene et al., Citation2019; Getachew & Mesele, Citation2018; Girma & Worku, Citation2020; Rabira et al., Citation2015; Worku & Girma, Citation2020).
Species richness
The present survey revealed 21 different large and medium-sized mammalian species from the FFL. Some studies that have used similar transect line techniques and in areas of different protection levels across the country and elsewhere revealed that the mammalian species recorded were lower than the results obtained from the present study. For example, Lemma and Tekalign (Citation2020) recorded a total of eight large and medium mammalian species in Humbo Community-Based Forest Area, Southern Ethiopia; Woldegeorgis and Wube (Citation2012) recorded 14 mammal species from Yayu forest in southwest Ethiopia; Getachew and Mesele (Citation2018) recorded even lower (12) mammal species in the Mengaza communal forest, East Gojjam, Ethiopia. This variation might account for variation in mammalian species group composition, variation in vegetation structure and human influence, and livestock grazing. The findings of the present study, therefore, highlight that FFL has valuable importance for the conservation of Ethiopia’s mammal species. In addition, the number of mammalian species recorded during the present study was also comparable to several other studies conducted in Ethiopia and elsewhere. For instance, Njoroge et al. (Citation2009) recorded 23 species in the Arawale National Reserve, Kenya; Bene et al. (Citation2013) recorded 23 species in the Sime Darby, Liberia; Girma et al. (Citation2012) recorded 19 species in the Wendo Genet, Ethiopia. The relative abundance of food sources, dense green vegetation cover, and availability of water (Lake Abaya) might be the major factors governing their abundance and species richness in the present study area.
The present study showed that globally vulnerable species such as Panthera leo, Panthera pardus, and Hippopotamus amphibius were recorded in the area, indicating the area is potential for mammalian biodiversity conservation.
Species relative abundance
Papio anubis and Chlorocebus aethiops were the most recorded and Civettictis civetta, Panthera leo, and Panthera pardus were the least recorded mammal species in the study area. The low abundance (frequency of records) of carnivores might be associated with their nocturnal behavior. As described by Wolf and Ripple (Citation2018), Gebresenbet et al. (Citation2018), Lemma and Tekalign (Citation2020), and Worku and Girma (Citation2020), most carnivore species are solitary, nocturnal and crepuscular so that their presence could not be easily documented.
The present study contradicts the hypothesized trend of a higher frequency of records during the wet season than the dry season because of resource availability. For example, the number of records of mammalian species recorded during the dry season (377) surpassed the number recorded during the wet season (308). Moreover, most of the species of the FFL such as Lepus habessinicus, Mellivora capensis, Crocuta crocuta, Chlorocebus aethiops, Colobus guereza, Papio anubis, Orycteropus afer, Hippopotamus amphibius, Phacochoerus aethiopicus and Ourebia ourebi were recorded relatively in a lower frequency of records during the wet season compared with the dry season. This in line with the work of Kasso and Bekele (Citation2017) in the Assela fragmented forest, Ethiopia, but disagrees with the work of Worku and Girma (Citation2020) in the Geremba forest of Southern Ethiopia where more mammalian species were observed during the wet season than the dry season. The possible explanation for this could be the growth of herbaceous and ground vegetation provided thick cover for the mammalian species, which makes the sighting of them difficult (Diriba et al., Citation2020; Girma et al., Citation2012; Girma & Worku, Citation2020; Qufa & Bekele, Citation2019).
Species distribution
The results of the present study showed that of the 685 total records, the frequency of records was higher in the forest (30.80%) followed by wetland. The result agrees with other studies (Bakala & Mekonen, Citation2020; Rabira et al., Citation2015).
All species recorded in the forest, wetland, and cultivated land (except Panthera leo, Colobus guereza, and Panthera pardus) habitats are subsets of the species recorded in the grassland habitat. Given the small size of the grassland habitat compared with the forest, these results are surprising and disagree with the well-established area-species relationship; which states that habitats with greater area tend to contain a higher number of species compared with habitats with a smaller area (Bakala & Mekonen, Citation2020; Diriba et al., Citation2020; Udy et al., Citation2021; Worku & Girma, Citation2020). Specifically, the presence of a large number of order Artiodactyla (herbivore species) guilds found in the grassland, as a result of higher habitat quality, might have also attracted a high number of order Carnivora species and resulting in increased diversity (Diriba et al., Citation2020; Fetene et al., Citation2019; Girma & Worku, Citation2020). Therefore, the different habitats should be given equivalent conservation attention. Further focused studies are needed on prey-predator relationships for impactive management planning in the FFL.
Species assemblage of the wetland habitat was nine and dominated by Hippopotamus amphibius and Chlorocebus aethiops . This indicates that, despite hosting the lowest number of species, the wetland habitat supports species that are unique to that habitat type, specifically the vulnerable Hippopotamus amphibius. Thus, the wetland habitat plays a complementary role in increasing mammal diversity and water sources for mammalian species of the FFL. Similar results have been demonstrated by a number of studies (Bakala & Mekonen, Citation2020; Fetene et al., Citation2019; Rabira et al., Citation2015; Udy et al., Citation2021; Worku & Girma, Citation2020), suggesting that a combination of wetland and other habitats is crucial to the long-term maintenance of viable populations of some species.
Panthera pardus and Panthera leo are the most widely distributed cats in the world, where food and cover are available (Burgin et al., Citation2018; Wolf & Ripple, Citation2018); however, they are vulnerable and at risk of local extinction (IUCN, Citation2021; Lavrenchenko & Bekele, Citation2017; Ripple et al., Citation2014; Tefera, Citation2011). Furthermore, in the present study area, they were restricted to wetland and forest habitats, respectively. This might be due to the conflict with the local people due to predation for domestic animals (Tefera, Citation2011). Also, this might contribute to the rareness of these species. In the present study, Hippopotamus amphibius is distributed in three habitat types and the most abundant in wetland habitat. This might be because of their ecological preferences and adaptation of mammalian species playing a role in their distribution in different habitat types (Penjor et al., Citation2021; Tamrat et al., Citation2020; Udy et al., Citation2021; Wolf & Ripple, Citation2018). The presence of these conservation concern species demonstrates that the study landscape is a potential area for wildlife conservation.
Diversity index of the landscape
The species index of the diversity of the study area showed higher species richness (H = 2.56; 1-D = 0.8968) than to study conducted by Qufa and Bekele (Citation2019) in the Lebu natural protected forest, Ethiopia (H = 2.119; 1-D = 0.8167). The grassland habitat is characterized by greater species richness and Shannon diversity index, and vice versa for wetland habitats. The present study was also comparable to the species diversity index (D = 7.142) recorded in the Geremba forest by Worku and Girma (Citation2020). Different possible factors like availability of food sources, dense forest cover, and water might be contributed to higher species richness.
Conclusion and recommendations
The findings of the study reveal that FFL supports a considerable number of medium and large mammalian species, including three globally threatened species: the vulnerable Panthera pardus, Panthera leo, and Hippopotamus amphibius (IUCN, Citation2021). This is the first ecological information on the diversity of mammalian species of the FFL, which would serve as valuable baseline information for stakeholders to make impactive conservation decisions and for researchers wishing to conduct related ecological studies. Additionally, the least conservation concern species such as Papio anubis, Chlorocebus aethiops, and Phacochoerus aethiopicus are highly recorded in the study area across habitat types and seasons. The Simpson index showed that the area harbors diverse mammalian species. The number of medium and large-sized mammalian species recorded in the study area is higher and comparable to other localities in Ethiopia and elsewhere using similar transect line technique, sampling, and direct and indirect field methods.
Despite the importance of FFL as the home for 21 mammalian species belonging to six orders and 13 families, it is not legalized as a wildlife refuge area. Therefore, to ensure the long-term conservation of the mammalian species of the forest, the following recommendations are suggested: The federal and regional governments should legalize it as a wildlife refuge area to conserve mammalian species of the area. Clear demarcation of the area is also essential. Local community and knowledge-based conservation and management initiatives must be implemented in the area.
Study limitations and future research directions
Limitations of this study that might affect the strength of inference are carefully considered. Due to logistical constraints, we were unable to access many portions of the study area or to implement a random sampling design, so our transect lines survey may represent a biased sample yielding incomplete information on mammalian species occurrence patterns in the landscape. Nevertheless, we were able to sample across strata in our hypothesized factors of influence, and we attempted to control for the impacts of spatial and temporal sampling, such as habitats and seasons, on occurrence.
We infrequently recorded several of the mammalian species in the FFL, a common challenge in surveys of rare and elusive species (e.g., carnivores), and despite advantages of both direct and indirect sign survey approach, stronger inference is ultimately achieved only by greater sampling effort (including more targeted, species-specific sampling) (Penjor et al., Citation2021).
Our indices representing spatial and temporal features of hypothesized importance were generated from the best available information, but their reliability may be diminished by associated uncertainty. Transect data underestimated the occurrence and abundance of smaller species (Gebresenbet et al., Citation2018), so corresponding diversity indices are dominated by the larger and better-detected species. Future work focusing more specifically on these smaller species and illusive carnivore species is therefore in need.
Important variation in mammalian habitat quality may not have been adequately described by habitat types, which might be more tightly linked to the ecological characteristics of certain species (Udy et al., Citation2021). For example, specie richness is higher in cultivated lands than wetland, while such survey show great promise for improving ecological understanding across spatial and temporal scales, they are not a substitute for detailed, field-based assessments of habitat that are largely lacking for FFL.
Nevertheless, such limitations are common to many human-dominated landscapes, particularly in developing nations like Ethiopia, and our study highlights a conservation relevant approach to characterizing a landscape. Future work should seek to test and improve upon these measures of landscape heterogeneity and address other important ecological and anthropogenic factors. Impact of vegetation, impact of human disturbance such as roads, human-wildlife relation (Newbold et al., Citation2015; Penjor et al., Citation2021), competition among mammals and home range (Tamrat et al., Citation2020) and advanced survey approach (e.g., camera trapping (Penjor et al., Citation2021) should be surveyed.
Availability of data
All data used are included in the article and supplementary material.
Authors’ contributions
Berhanu Gebo, Serekebirhan Takele and Simon Shibru conceived, designed the study data collection. Berhanu Gebo conducted fieldwork, analysis, write the manuscript and revised the whole document. Serekebirhan Takele and Simon Shibru designed the survey method, edited the manuscript and revised the final version of the main document for submission for potential review. All authors contributed to the writing of the manuscript and approved the submitted version.
Acknowledgments
Our special gratitude goes to the Mirab Abaya Wereda Administrative office for allowing us to research in the Faragosa-Fura landscape. We also duly acknowledge Faragosa and Fura Kebeles administrative office and agricultural extension workers for their assistance during data collection in the habitat types. We also thank the Department of Biology, College of Natural Science, Arba Minch University, for their invaluable logistics and financial support. We are thankful for the Arba Minch College of Teachers education (Ayalew Abera, Yaricho Yaya, Daniel Befikadu) for the logistic support as well as covering the living and other costs of the Ph.D. candidate and we are grateful for the support provided.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Amare, A. (2015). Wildlife resources of Ethiopia: opportunities, challenges and future directions: From ecotourism perspective: A review paper. Natural Resources, 6(6), 405–422. https://doi.org/10.4236/nr.2015.66039
- Bakala, F., & Mekonen, G. (2020). Species diversity and relative abundance of medium and large-sized wild mammalian species: A study from adaba community forest, West Arsi Zone, Southeast Ethiopia. African Journal of Ecology, 59(2), 38–43. https://doi.org/10.1111/aje.12827
- Belete, T., & Melese, M. (2016). Assessment of large mammalian species potential in Tululujia Wildlife Reserve, Southwestern Ethiopia. International Journal of Agricultural and Life Sciences. https://doi.org/10.22573/spg.ijals.016.s12200070
- Bene, J. K., Bitty, E. A., Bohoussou, K. H., Abedi-, M., Gamys, J., & Soribah, P. A. J. (2013). current conservation status of large mammalian species in Sime darby oil palm concession in Liberia. Global Journal of Biology, Agriculture & Health Sciences, 2(3), 93–102
- Burgin, C. J., Colella, J. P., Kahn, P. L., & Upham, N. S. (2018). How many species of mammalian species are there? Journal of Mammalogy, 99(1), 1–14. https://doi.org/10.1093/jmammal/gyx147
- Diriba, G., Tamene, S., Mengesha, G., & Asefa, A. (2020). Diversity of medium and large mammalian species in the Loka Abaya National Park, southern Ethiopia. Ecology and Evolution, 10(18), 9896–9905. https://doi.org/10.1002/ece3.6649
- ENMSA. (2019). Ethiopia national meteorological service agency, data base for metrological data source (2009‐2019) of Mirab Abaya District, Ethiopia,Mega publisher
- Fetene, A., Yeshitela, K., & Gebremariam, E. (2019). The impacts of anthropogenic landscape change on the abundance and habitat use of terrestrial large mammalian species of Nech Sar National Park. Environmental Systems Research, 8(1), 1–16. https://doi.org/10.1186/s40068-019-0147-z
- Gebresenbet, F., Baraki, B., Yirga, G., Sillero-Zubiri, C., & Bauer, H. (2018). A culture of tolerance: Coexisting with large carnivores in the Kafa Highlands, Ethiopia. ORYX, 52(4), 751–760. https://doi.org/10.1017/S0030605316001356
- Getachew, A., & Mesele, Y. (2018). Species composition and relative abundance of medium and large mammalian species in Mengaza communal forest, East Gojjam, Ethiopia. Journal of Ecology and the Natural Environment, 10(2), 34–40. https://doi.org/10.5897/jene2017.0667
- Girma, Z., Mamo, Y., & Ersado, M. (2012). Species composition, distribution and relative abundance of large mammalian species in and around wondo genet forest patch, Southern Ethiopia. Asian Journal of Applied Sciences, 5(8), 538–551. https://doi.org/10.3923/ajaps.2012.538.551
- Girma, Z., & Worku, Z. (2020). Large mammal diversity in Nensebo Forest, Southern Ethiopia. International Journal of Zoology, 2020, 1–11. https://doi.org/10.1155/2020/8819019
- IUCN. (2021). The IUCN red list of threatened species. Version 2020-3.12 January 2021. https://www.iucnredlist.org
- Kasso, M., & Bekele, A. (2017). Diversity, abundance and distribution of mammalian species in Fragmented Remnant Forests around Asella Town, Ethiopia. MAYFEB Journal of Biology, 1(January).
- Kingdong, J. (2015). Kingdon field guide to African mammalian species-bloomsbury natural history (2nd ed.).
- Lavrenchenko, L. A., & Bekele, A. (2017). Diversity and conservation of Ethiopian mammalian species: What have we learned in 30 years? Ethiopian Journal of Biological Sciences, 16, 1–20.
- Legese, K., Bekele, A., & Kiros, S. (2019). A survey of large and medium-sized mammalian species in Wabe forest fragments, Gurage zone, Ethiopia. International Journal of Avian & Wildlife Biology, 4(2), 32–38. https://doi.org/10.15406/ijawb.2019.04.00149
- Lemma, A., & Tekalign, W. (2020). Abundance, species diversity, and distribution of diurnal mammalian species in humbo community-based forest area, Southern Ethiopia. International Journal of Zoology, 2020, 1–5. https://doi.org/10.1155/2020/5761697
- Mekonen, S. (2020). Coexistence between human and wildlife: The nature, causes and mitigations of human wildlife conflict around Bale Mountains National Park, Southeast Ethiopia. BMC Ecology, 20(1), 1–9. https://doi.org/10.1186/s12898-020-00319–1
- Negeri, D., Gadisa, T., & Habtamu, T. (2015). The diversity, distribution and relative abundance of medium and large-sized mammalian species in Baroye Controlled Hunting Area, Illubabor Zone, Southwest Ethiopia. International Journal of Molecular Evolution and Biodiversity, 5(4), 1–9. https://doi.org/10.5376/ijmeb.2015.05.0004
- Newbold, T., Hudson, L. N., Hill, S. L. L., Contu, S., Lysenko, I., Senior, R. A., Börger, L., Bennett, D. J., Choimes, A., Collen, B., Day, J., De Palma, A., Díaz, S., Echeverria-Londoño, S., Edgar, M. J., Feldman, A., Garon, M., Harrison, M. L. K., Alhusseini, T., & Purvis, A. (2015). Global impacts of land use on local terrestrial biodiversity. Nature, 520(7545), 45–50. https://doi.org/10.1038/nature14324
- Njoroge, P., Yego, R., Muchane, M., Githiru, M., Njeri, T., & Giani, A. (2009). A survey of the large and medium sized mammalian species of Arawale National Reserve, Kenya. Journal of East African Natural History, 98(1), 119–128. https://doi.org/10.2982/028.098.0108
- Penjor, U., Wangdi, S., Tandin, T., & Macdonald, D. W. (2021). Vulnerability of mammal communities to the combined of anthropic land-use and climate change in the Himalayan conservation landscape of Bhutan. Ecological Indicators, 121, 107085. https://doi.org/10.1016/j.ecolind.2020.107085
- Qufa, C. A., & Bekele, A. (2019). A preliminary survey of medium and large-sized mammalian species from lebu natural protected Forest, Southwest Showa, Ethiopia. Ecology and Evolution, 9(21), 12322–12331. https://doi.org/10.1002/ece3.5733
- Rabira, G., Tsegaye, G., & Tadesse, H. (2015). The diversity, abundance and habitat association of medium and large-sized mammalian species of Dati Wolel National Park, Western Ethiopia. International Journal of Biodiversity and Conservation, 7(2), 112–118. https://doi.org/10.5897/ijbc2014.0808
- Reeder, D. M., Helgen, K. M., & Wilson, D. E. (2019). Global trends and biases in new mammal species discoveries. Global Trends and Biases in New Mammal Species Discoveries. https://doi.org/10.5962/bhl.title.156951
- Ripple, W. J., Estes, J. A., Beschta, R. L., Wilmers, C. C., Ritchie, E. G., Hebblewhite, M., Berger, J., Elmhagen, B., Letnic, M., Nelson, M. P., Schmitz, O. J., Smith, D. W., Wallach, A. D., & Wirsing, A. J. (2014). Status and ecological impacts of the world’s largest carnivores. Science, 343, 6167. https://doi.org/10.1126/science.1241484
- Sutherland, W. J. (2006). Ecological census techniques: A handbook (Second ed.). Cambridge University Press.
- Udy, K., Fritsch, M., Meyer, K. M., Grass, I., Hanß, S., Hartig, F., Kneib, T., Kreft, H., Kukunda, C. B., Pe’er, G., Reininghaus, H., Tietjen, B., Tscharntke, T., Van Waveren, C. S., & Wiegand, K. (2021). Environmental heterogeneity predicts global species richness patterns better than area. Global Ecology and Biogeography, 30(4), 842–851. https://doi.org/10.1111/geb.13261
- Wale, M. (2017). Wildlife threats and their relative severity of Eastern Ethiopia Protected Areas. Ecology and Evolutionary Biology, 2(4), 59. https://doi.org/10.11648/j.eeb.20170204.12
- Wolf, C., & Ripple, W. J. (2018). Rewilding the world’s large carnivores. Royal Society Open Science, 5(3). https://doi.org/10.1098/rsos.172235
- Worku, Z., & Girma, Z. (2020). Large mammal diversity and endemism at Geremba Mountain Fragment, Southern Ethiopia. International Journal of Ecology, 2020, 1–11. https://doi.org/10.1155/2020/3840594
- Yalden, D. W., Largen, M. J., Kock, D., & Yalden, D. W. (1986). Catalogue of the mammalian species of ethiopia: 6. perissodactyla, proboscidea, hyracoidea, lagomorpha, tubulidentata, sirenia and cetacea. Monitore Zoologico Italiano, Supplemento, 21(1), 31–103. https://doi.org/10.1080/03749444.1986.10736707
- Tamrat, M., Atickem, A., Tsegaye, D., Nguyen, N., Bekele, A., Evangelista, P., Fashing, P. J., & Stenseth, N. C. (2020). Human-wildlife conflict and coexistence: A case study from Senkele Swayne’s Hartebeest Sanctuary in Ethiopia. Wildlife Biology, 2020(3),1-9. https://doi.org/10.2981/wlb.00712
- Tefera, M. (2011). Wildlife in Ethiopia: Endemic Large Mammalian species. World Journal of Zoology, 6(2), 108–116
- Woldegeorgis, G., & Wube, T. (2012). A Survey on Mammalian species of The Yayu Forest in Southwest Ethiopia. Journal of Science, 35(2), 135–138