
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
The present study aimed to analyse the population structure, forest composition and regeneration dynamics of Betula utilis D.Don in two core zone of Nanda Devi Biosphere Reserve. A total of 346 plants species belonging to 198 genera and 82 families were recorded dominated by Asteraceae (24) followed by Rosaceae (21 species) and Ranunculaceae (19 species), respectively. The total tree stand density was ranged between 1133 ind.ha−1 and 2137 ind.ha−1 in the south and north aspect of the each sites. Among tree species B. utilis was showed the dominant tree species with highest density ranged between 663 ind.ha−1 and 1525 ind.ha−1 at north and south aspects of both sites. Highest seedling density (1033 saplings ha−1 and 1783 seedlings ha−1) and sapling density (1033 saplings ha−1 and 1470 saplings ha−1) was recorded for B. utilis in north and south aspects. The density-diameter curve of B. utilis forest resembled a reverse J-shape which exhibits good regeneration with higher number of individuals in seedling and sapling stages, followed by a sharp decline in tree classes. This study suggests urgent need of long term ecological monitoring of timberline zone to assess the significant change in timberline vegetation and development of sustainable management plans.
1. Introduction
Among the different forest communities along the Himalayan region, a definite boundary is frequently developed by the upper limit of tree growth and krumholz vegetation of the alpine meadows, generally known as timberline ecotone (Korner, Citation1998a; Holtmeier & Broll, Citation2005) and are known to be sensitive to biotic and climatic stressors (Allen & Breshears, Citation1998; Kullman, Citation1998). The Himalayan region has the highest and most diverse timberline in the world (Korner, Citation2003). The geographical setup, topography, undulant landscape, microclimatic conditions, aspects, herbivory and anthropogenic pressure always play a significant role in determining the structure, composition and regeneration pattern of plant communities along the timberline ecotone (Cairns & Moen, Citation2004; Holtmeier, Citation2009; Holtmeier & Broll, Citation2007; Korner, Citation2012; Weiser & Tuasz, Citation2007). The timberline zone is largely dominated by Betula utilis (Himalayan birch) and covers the ecotone or buffer zone between the dense coniferous forest and the sub-alpine and alpine areas in Western Himalaya. The natural B. utilis forest with its plant communities in ecologically significant region is climax vegetation, but changing dynamically. However, the B. utilis holds intrinsic socio-cultural, religious, spiritual and economic values that are inherent elements of the hill people quality of life. It is among the few keystone and native species of timberline ecosystem locally known as “Bhojpatra” belonging to the family betulaceae, play a significant role to maintaining the structure and functioning of the timberline ecosystem and safeguard the micro-environment. There are about fifty species of this genus all over the world, and three species are reported from India, viz. Betula alnoides. Ham. EXD. Don Betula cylindrostachys Gamble and Betula utilis D. Don Syn bhojpatra Lindley. B. utilis grows in temperate to timber line zone mostly in association with Cedrus deodara, Taxus baccata, Pinus wallichiana, Abies pindrow, Rhododendron anthopogon, Juniperus communis and Salix species etc. between the altitudinal ranges of 3000–4200 masl, but some time descending sporadically as low as 1800 m asl (Phondani et al., Citation2010). In Western Himalaya it is distributed mostly in Timberline areas of Harikidun, Gangotri, Gaumukh, Yamunotri, Kedarnath, Tungnath, Rudranath, Valley of Flowers, Bhyundar and Niti valley etc. (Naithani, Citation1984). Tree is endemic to the Himalayan region and is also used during religious ceremonies and has a prominent role in socio-cultural values of the Himalayan people. During ancient times, “Birch” bark was used for literary purposes as writing material in place of paper (Gaur, Citation1999). The bark, stings and leaves of Betula utilis have various medicinal properties and is used against various health ailments (Naithani, Citation1984). Oflate timberline zones of Central Himalaya have been increasingly affected by loss of biodiversity due to large scale exploitation of bioresources due to anthropogenic interferences. During recent past it has been observed that B. utilis is facing severe threat due to the over exploitation for medicinal and fuel purposes, overgrazing, forest fire, pathogen attacks, defoliation, snow drift, lighting, climatic variability, erosion and landslide etc. are the reasons for declining the population although the species is already in the list of threatened plant as per by IUCN (Ved et al., Citation2003). Despite several studies on plant communities along altitudinal gradients in the Himalaya (Acharya et al., Citation2011; Bhattarai & Vetaas, Citation2006; Bhattarai et al., Citation2004; Kharkwal et al., Citation2005), timberline ecotone (Bharti et al., Citation2012; Gairola et al., Citation2014; Rai et al., Citation2013; Singh et al., Citation2012), impact of climate change on vegetation (Chawla et al., Citation2012; Grabherr et al., Citation1994; Grace et al., Citation2002; Kanka et al., Citation2005; Kumar, Citation2012), it is apparent that there still exist several gaps in our understanding of the functional and phylogenetic patterns of plant diversity along altitudinal gradient and also about the climatic controls of seedling recruitment dynamics and ecophysiology of some key timberline species. Therefore, the aim of present study is to investigate the population structure, species composition and regeneration pattern in the timberline zone with particular reference to Betula utilis to formulate appropriate strategies for conservation and management on one hand and add to the existing knowledge gap on the other.
2. Materials and methods
2.1. Study area
The study was carried out in two core zones of Nanda Devi Biosphere Reserve (NDBR), viz., (A) Valley of Flower National Park (VoFNP, 30° 41ʹ N to 79° 33ʹ E) and (B) Nanda Devi National Park (NDNP, 30°16ʹ N to 79° 44ʹ E) along the timberline area with elevation ranges between 2800 m to 4200 m asl in Western Himalaya, Uttarakhand. NDBR was recognized as a World Heritage Site in 1992 and was included in the UNESCO’s world network of Biosphere Reserves in 2004. The timberline forest of NDBR harbors rich diversity of unique plant species particularly of medicinal plants (Hajra & Balodi, Citation1995; Kala et al., Citation1998; Samant, Citation1993; Silori, Citation2001; Srivastava, Citation1999). The climate of the study area is characterized by short cool summers and long severe winters.
2.2. Plot layout and tree inventory
The study was conducted in two major forest types; sites having pure patches of B. utilis stand are considered as Pure B. utilis forest while sites having mixed tree species with dominant B. utilis species as mixed B. utilis forest with different aspect North and South (N-S). A total of eight forest stands having size of 1 hectare (4 each in North and South aspects) in pure and mix B. utilis community were selected for the detailed study. While the selected stands, a total of 160 plots of 10 m × 10 m were laid randomly for enumeration of tree species. Each of the 10 × 10 m quadrat was further subdivided into 5 m × 5 m sub-quadrats for enumeration of shrub and saplings, and further 1 m × 1 m plots were laid randomly within the 10 m × 10 m plots for studying the seedlings and herb species (Misra, Citation1968). Trees in each quadrate were surveyed for height and diameter at breast height (those exceeding height of 2 m). Due to the slow growth rate and low stature of trees along the timberline, a tree having below 2 m height were considered as a tree and girth classes was prepared with the difference of 10 cm. All the classes were divided into seedling, sapling and different girth classes viz., A: 20–30 cm, B: 31–40 cm, C: 41–50 cm, D: 51–60 cm, E: 61–70 cm, F: 71–80 cm, G: 81–90 cm and H: >90 cm. In each quadrate, all trees (≥20 cm cbh,) saplings (10–20 cm cbh) and seedling (≤10 cm collar height circumference) were individually measured. The sum of the relative values, viz., frequency, density and dominance were used for calculating importance value index (IVI) (Curtis, Citation1959). Tree diversity was determined by using Shannon-Wiener diversity index (Shannon & Weaver, Citation1949) as:
where, H is Shannon-Wiener diversity index, pi is the proportion of individuals belonging to species i and ln is natural log (i.e., 2.718). Regeneration status of species was totally based on population size of seedlings and saplings. The regeneration status of dominant trees was assessed based on proportional distribution of density of individuals in each seedling, sapling and adult tree species in forest stand. The regeneration of tree species was considered “good” when seedling density > sapling density > adult tree density; “fair” when seedling density > sapling density ≤ adult density; “poor” when the species survived in only the sapling stage but not in the seedling stage; “none” for species with no sapling or seedling stages but present as adult trees; and “new” when adults of a species were absent but sapling and/or seedling stage(s) were present (Khan et al., Citation1987; Shankar, Citation2001).
3. Results
3.1. Species life forms
A total of 346 species of vascular plants (i.e., Angiosperms, Gymnosperms and Pteridophytes) belonging to 198 genera and 82 families were recorded. Of these, 66 families and 167 genera were of angiosperm, 4 families and 8 genera were of gymnosperms and 12 families and 17 genera were of pteridophytes were recorded between 2600–4600 m asl from the both study sites of NDBR. Asteraceae (25 spp.) dominated the list of families followed by Rosaceae (21 spp.), Ranunculaceae (19 spp.), Poaceae (14 spp.), Polygonaceae (12 spp.), Lamiaceae (10 spp.) and Fabaceae (9 spp.), respectively. In Gymnosperms, species diversity within the family is poorly represented in the site and families Pinaceae (5 spp.) and Cupressaceae (4 spp.) were the dominant ones. The dominant families of Pteridophytes are Dryopteridaceae (7), Athyriaceae (5) and Davalliaceae (3 spp.). The dominant genera among the angiosperm are Saxifraga (8), Polygonum (6), Primula and Berberis (5 spp. In each), Potentilla, Anemone, Araceae and Asteraceae (4 spp. in each). The floristic composition and forests type changed from lower mixed B. utilis forest (2800–3500 m asl) to pure B. utilis and Rhododendron forest (3500–4200 m asl) at both south and north aspects ().
Table 1. General characteristic features of the selected sites of the timberline region of Nanda Devi Biosphere Reserve (NDBR).
3.2. Diversity-dominance parameters
The variation in Shannon-Wiener and Simpson diversity index as computed for different layer of plant species at each sites (). In pure B. utilis forests of VoFNP the highest Simpson diversity index was recorded for tree layer (0.60 and 0.82) and minimum for herb layer (0.02 and 0.04) at both north and south aspects, respectively. Whereas in mixed forest, maximum values was recorded for seedling layer (0.49 and 0.21) and minimum for herb layer (0.04 and 0.07) at south and north aspect, respectively. In pure B. utilis forest of NDNP the maximum Simpson diversity index was observed for tree layer (0.77) followed by seedling (0.62); and minimum value for shrub layer (0.12) at North aspect. However, in mixed B. utilis forest, maximum Simpson diversity index was estimated for seedling layer (0.45) followed by tree (0.35) and sapling layer (0.30); and minimum value was recorded for shrub (0.05) layer at North aspect. The Shannon-Wiener species diversity index (H) in pure B. utilis forests represented highest by herb (3.47) and lowest by tree layer (0.36) at North aspect whereas in mixed forests, herb layer exhibits highest (2.98) and seedling layer showed lowest (0.91) at South aspect of VoFNP. However, in NDNP, the highest Shannon-Wiener diversity was recorded for herb layer (2.26) and lowest (0.41) for tree layer at North aspect. Whereas in mixed B. utilis forest, highest (2.56) and lowest (0.96) Shannon-Wiener diversity was recorded for shrub layer at South and North aspects, respectively.
Table 2. Diversity-dominance parameters in Valley of Flower National Park (VoFNP) and Nanda Devi National Park (NDNP).
3.3. Population density, composition and structure
The total tree stand density was recorded 1820 ind.ha−1 in the south aspect and 1420 ind.ha−1 at north aspect in the VoFNP (). B. utilis was found dominant tree species with highest density (1254 ind.ha−1), IVI (266.56) in north aspect as compared to the south aspect. The minimum density (16 ind.ha−1) was recorded for Taxus baccata in the north aspect while, Acer caesium (12 ind.ha−1) in south aspect. The higher total basal cover was observed for B. utilis (28.61 m2ha−1) and minimum for A. caesium (2.6 m2ha−1) in the south aspect. Rhododendron campanulatum (IVI −13.44 and IVI-32.28) and Abies pindrow (IVI-12.48 and IVI-13.98) were found as co-dominant species in both north and south aspects, respectively. The mixed B. utilis forest showed total density of 1246 ind.ha−1 in the north aspect and 1462 ind.ha−1 in the south aspect. In the north aspect, highest density (663 ind.ha−1) and IVI (136.48) were recorded for B. utilis. Similarly, in south aspect B. utilis was found dominant species with maximum density (487 ind.ha−1) and IVI (85.68). The minimum density was recorded for the Vibrunum grandiflorum (6 ind.ha−1), followed by Sorbus foliolosa at both aspects. The total basal cover of all species was observed 126.29 m2ha−1 at north and 64.46 m2ha−1 at south aspect, respectively. In the north aspect A. pindrow (IVI-72.16), Cedrus deodara (IVI-32.4), A. caesium (IVI-18.89) and R. campanulatum (IVI-15.62), whereas at south aspect C. deodara (IVI-63.43), A. pindrow (IVI-48.69), T. baccata (IVI-38.93) and A. caesium (IVI-22.64) were found the co-dominant tree species ().
Table 3. North and south aspect of pure and mixed Betula utilis forest of valley of flower National Park.
In the tree layer B. utilis (IVI-217.31) was found to be the most dominant tree species followed by R. campanulatum and A. pindrow at both North and South aspect at NDNP (). The total tree stand density (2137 ind.ha−1) was recorded maximum at north aspect as compared to the south aspect (1133 ind.ha−1). Among the tree species B. utilis showed highest density (1525 ind.ha−1 and 793 ind.ha−1) and IVI (217.31 and 255.8) whereas A. caesium (12 ind.ha−1) and S. foliolosa (23 ind.ha−1) exhibits lowest at north and south aspects, respectively. The basal cover was also recorded maximum for B. utilis (22.2 m2ha−1 and 26.8 m2ha−1) and lowest for A. caesium and S. foliolosa. The total tree density was found higher at north aspect (2145 ind.ha−1) for mixed B. utilis forest as compared to south aspect (920 ind.ha−1). North aspect showed highest density (854 ind.ha−1) and IVI (114.32) of B. utilis, whereas minimum density was recorded for the Picea smithiana and A. caesium at both the aspects (). In the north aspect tree species such as P. wallichiana, A. pindrow, Populus ciliata and S. sikkimensis were found the co-dominant tree species whereas in the south aspect it was represented by A. pindrow, P. wallichiana, C. deodara and T. baccata ().
Table 4. North and south aspect of pure and mixed Betula utilis forest of Nanda Devi National Park.
3.4. Regeneration status of dominant tree species
The density-diameter curve of the tree population of Betula utilis both in pure and mixed forest resembled a reverse J-shape () which exhibit the both timberline forests regenerating well with higher number of individuals in the seedling and sapling stages, followed by a sharp decline in tree classes at different sites and aspects of the timberline areas (). However the population structure of B. utilis exhibited more or less similar trends among the pure and mixed forest across different aspects. High accumulation of seedlings and saplings and rapid decline of individuals in tree-size class were observed for Betula at North aspect of NDNP ()) and other associated species (i.e., A. pindrow, S. foliolosa, T. baccata) at all the pure Betula forest (–d)). R. campanulatum exhibited similarity with the overall forest structure () however, a accumulation of individuals at sapling stage and a sharp decline toward the seedling and tree size classes was observed at North aspect of NDNP ()). Among other species A. pindrow, S. sikkimensis and T. baccata showed a greater proportion of individuals in the seedling and sapling classes and a sharp decline toward the tree-size classes ().
Figure 1. Population structure of pure B. utilis dominated forest; (a) North aspect of VoFNP, (b) South aspect of VoFNP, (c) North aspect of NDNP, (d) South aspect of NDNP.
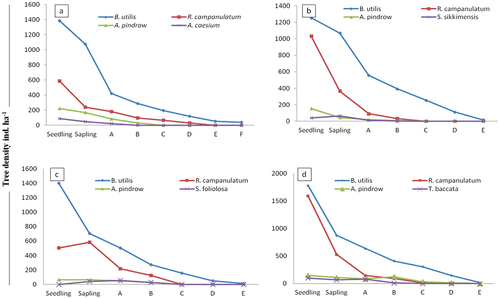
Figure 2. Population structure of mixed B. utilis dominated forest; (a) North aspect of VoFNP, (b)South aspect of VoFNP, (c) North aspect of NDNP, (d) South aspect of NDNP.
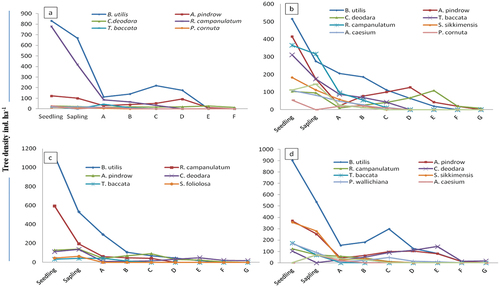
In NDNP, highest seedling density was recorded for B. utilis (1783 seedlings ha−1 and 1306 seedlings ha−1) in north and south aspects, respectively. However, total seedlings density was found maximum (3628 ha−1 and 2206 ha−1) in pure and mixed Betula forests at north aspect, respectively. Sapling stratum showed higher density and was ranged between 875 saplings ha−1 to 676 saplings ha−1 for B. utilis in North and South aspect of pure forest. The overall total sapling density was recorded 1529 saplings ha−1 and 1146 saplings ha−1 at north and south aspect.
In VoFNP, total seedling density for pure Betula utilis forest was recorded between 2850 seedling ha−1 and 2533 seedling ha−1 in north and south aspects. However, higher sapling density (1033 saplings ha−1 and 1470 saplings ha−1) was recorded for B. utilis and lowest was recorded for S. foliolosa and A. pindrow at north and south aspects in pure forest of VoFNP, respectively.
4. Discussion
Himalayan mountain ranges play a significant role in impacting the regional and global climate. Further, being governed by low temperature, high altitude regions of the world are more responsive to the changing climatic conditions and hence better indicators of the same (Grabherr et al., Citation1994; Price & Haslett, Citation1995). The growth and reproduction of plant communities here are mainly controlled by temperature (Grace et al., Citation2002) that gives rise to steep ecological gradients and narrow ecotones (Pauli et al., Citation2004); and any minor change in temperature may lead to change in tree line and nival zones. Studies on the population structure along altitudinal gradients of a mountain particularly of protected areas would be helpful in understanding the influences of environmental factors on the regeneration of natural forests that help in designing strategies for management. The monitoring of protected area is important due to their geographical uniqueness and the presence of good populations of threatened species of plants and animals. Woody communities were segregated and studied for species diversity along an elevational gradient and it was observed that the density of tree species increased with increasing altitude, whereas the species diversity was declined. The timberline forests are considered most climate sensitive ecotone and limiting factor for survival of tree growth in Himalayan ecosystem (Schickhoff, Citation2005). B. utilis widespread broadleaf timberline species which formed a continuous tree line in the core zone of NDBR. The timberline reaches at higher ranges in the north aspect (4200 m) as compared to south aspect (3600 m). The seedling and sapling growth of B. utilis was found higher in north aspect where soil moisture is expected high due to the increased elevation. Besides, north aspect in Himalaya usually receives quite low amount of the monsoon rain and depends on snowmelt water from permanent snow cover which maintain soil moisture during growing season (Shrestha et al., Citation2007). The continuous regeneration of B. utilis in both, the mixed forest at lower elevations and pure forest at higher elevations reported in earlier study by Schickhoff et al. (Citation2015) could be attributed to adequate soil moisture and nutrient at higher elevations for successful growth and establishment of the B. utilis at north aspect (Carlton & Bazzaz, Citation1998). The overall population structure of plant species showed highest contribution of seedlings to the total density in both the forests and aspects. This indicates good regeneration of tree species inside the core zone of Biosphere Reserve.
The species richness and plant diversity indicated large variation with respect to increasing altitude at both pure and mixed B. utilis forests in timberline zone of VoFNP and NDNP. A total of 4 tree species were recorded in the pure forest of B. utilis while 11 species were found in the mixed forest with dominant B. utilis at south and north aspects of timberline. The density of B. utilis was found higher in the north aspect in both the forest types of NDNP as compared to VoFNP. The distribution pattern of the species at timberline zone controlled by low air temperature, soil temperature and moisture during growing season at regional and global scale (Dimri & Dash, Citation2011; Korner, Citation2012; Paulsen & Körner, Citation2014). Considering the major compositional attributes, an increase in altitude and aspects (North & South) came with an important factor to control tree density and the total basal area for all the sites. Population structure studies are important as they reveal the dominance status of species and development within the community (Gairola et al., Citation2014). The structural characteristics are also important to explain niche requirement of species, spatial heterogeneity, temporal dynamics of understory vegetation and regeneration dynamics of dominant tree species (Chen & Franklin, Citation1995; Chen et al., Citation1993). All the timberline forest exhibited expanding population structure except North aspect of NDNP. Along the NW-SE gradient, north-facing slopes exhibit higher ranges of timberline zone (4200 m asl) from deciduous B. utilis to evergreen R. campanulatum dominated forest in NDBR. Accumulation of individuals in the sapling class and sharp decline toward both higher tree classes and lower seedling classes was apparent for other associated species (i.e., T. baccata, A. pindrow, C. deodara) of B. utilis in most of the forest sites indicates towards the replacement in tree size classes from sapling stage is not proportional and the population may decline in the long-term. Few other species i.e., Salix sikkimensis, Sorbus foliolosa other than B. utilis enjoy long-term persistence at lower elevations, Rhododendron campanulatum at higher elevations with more or less expanding population structures. The significance of individual population structure of any species should be having higher value as the environmental conditions at timberline forests do not permit most species to follow the normal life cycles (Gairola et al., Citation2014).
The presence of a sufficient number of seedling, sapling and mature trees in a given population indicate successful regeneration (Saxena & Singh, Citation1984). Seedling and sapling density did not exhibit uniform patterns for different sites and any strong relation with altitude however the higher tree seedling was found for B. utilis at upper limit (3600–4000 m asl) of the Biosphere Reserve. Dominant tree species (B. utilis) had good regeneration in the timberline while co-dominant species (T. baccata, S. foliolosa, V. grandiflorum and C. deodara) showed low regeneration in the mixed forest of both core areas of the Reserve. The regeneration of B. utilis was found good under open canopy forests whereas regeneration of T. baccata and S. foliolosa was found good under close canopy forest. The recruitment of seedling and survival of B. utilis and A. pindrow in timberline were found to be closely associated to annual change in snow accumulation, span of snow melting and precipitation during early stage of plant growth as well as adequate intensity of light (Wang et al., Citation2004). The maximum germination of seedling was observed during rainy season while highest mortality was noticed in winter season at both sites. Regeneration status of dominant tree species in mixed B. utilis forests was quite low as compared to pure patches of B. utilis. Higher elevation exhibit open canopy and are exposed to direct sunlight which is considered suitable for growth of seedling and sapling of B. utilis, while closed canopy forest with abundant leaf litter in the surface and low light intensity limit the recruitment of seedling and saplings of B. utilis in the mixed forest (Carlton & Bazzaz, Citation1998; Catovsky & Bazzaz, Citation2000; Mori & Takeda, Citation2004) and might also be affected by other site-specific abiotic factors viz., soil temperature, topography, slope, livestock grazing, and herbivory as revealed in earlier studies (Cairns & Moen, Citation2004; Grime, Citation2001; Korner, Citation1998). In some places mortality of the high girth classes of B. utilis was noticed (>20 tree ha −1) due to fungal infection. Aspect also influences seedling establishment, inhibiting regeneration success on South facing slopes exposed to high level of solar radiation resulted in maximum utilization of biresources i.e., fuelwood, timber, bark extraction etc. In some of the study sites, higher accumulation of seedlings at high altitude would indicate that fluctuations in climate might play an important role in synchronizing these patterns (Gairola et al., Citation2014).
The pure forest of B. utilis is also affected as compared to the mixed forest by the defoliators since the growth and establishment of species found better in the North aspect. The density of herbaceous vegetation was found less in the present study as compared to data reported in the earlier studies (Kala, Citation2005; Kala et al., Citation1998; Nautiyal et al., Citation2001) and may be largely attributed to all these kind of pressure in the timberline. The impact of herbivory in timberline particularly on B. utilis is an indicator of response of change in microclimate other biotic and abiotic factors in the region that could create conducive climatic conditions for proliferation of the different insect species in the region. Changes in environmental variables would also result changes in species distributional and affect their spatial relocation. In relation to altitude, expansion of plant species at timberline zone is an important indicator of climate change triggered by warming of the globe (Holtmeier, Citation2009; Pauli et al., Citation2007; Theurillat & Guisan, Citation2007). The seedlings and saplings of Betula utilis have been observed to extend their ranges and augment in the density of pure and mixed B. utilis forests to the extent of 268 ha and 224 ha in the timberline ecotone of NDBR, respectively (Maletha, Citation2017). Though, the long term scientific data to support timberline advancement or shift is not available, however local people’s perception/ responses & evidences clearly indicates that climate change impact is happening and visible in forest ecosystem in high elevation particularly in timberline and adjoining areas. Therefore, the present study suggests for long-term ecological monitoring to assess the impact of climate change on species composition and shifting of species to higher altitudes. Appropriate policies and management strategies are needed for socio-economic development of local inhabitant as well as other people involved in Cordyceps and MAPs collection from alpine meadows of NDBR.
5. Conclusion
Timberline ecotone among the regions in the Western Himalaya most sensitive to climate change and are characterized by a high level of endemism i.e., of plants and animals that occur no where else. Betula utilis being native species of timberline which is most prominent and represent significant ecological boundary where the upper most limit of forest/ sub-alpine forest vegetation terminates, beyond this, alpine pastures begin and therefore has been identified as sensitive eco-zone to the climate change process. The present findings provide an insight into timberline structure, composition and regeneration status which will be helpful for developing a conservation and sustainable management strategies of protection of Betula utilis which is one of the ecologically, socio-culturally, religiously and economically important species along with its other associated species in timberline zone in the wake of present and future climate change impact. The result of the present study also help to redesign a management plan and conservation policies based on scientific research and requirement of the dependent communities. Long term ecological research sites need to be established in high altitude timberline areas to monitor the environmental changes and their impacts and to development sustainable adaptation/mitigation measures. This network of people, scientist and conservationists should be linked to UNESCO’s global network of GLOCHAMORE/GLOCHAMOST partners for sustainable conservation of biodiversity as well as the protection of some keystone species.
Acknowledgments
The authors are thankful to Director, GBPNIHE, Kosi-Katarmal, Almora for providing facilities to undertake this work. We are extremely thankful to DST-SERB (SERB/SR/SO/PS/33/2011) for financial support for conducting this study. We are also thankful to the Chief Wildlife warden, Govt. of Uttarakhand, Conservator of Forest and Director, NDBR for their support and cooperation during the study in the VoFNP and NDNP (NDBR).
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Acharya, K. P., Vetaas, O. R., & Birks, H. (2011). Orchid species richness along Himalayan elevational gradients. Journal of Biogeography, 38(9), 1821–1833. https://doi.org/10.1111/j.1365-2699.2011.02511.x
- Allen, C. D., & Breshears, D. D. (1998). Drought-induced shift of a forest–woodland ecotone: Rapid landscape response to climate variation. Proceedings of the National Academy of Sciences, 95(25), 14839–14842. https://doi.org/10.1073/pnas.95.25.14839
- Bharti, R. R., Adhikari, B. S., & Rawat, G. S. (2012). Assessing vegetation changes in timberline ecotone of Nanda Devi National Park, Uttarakhand. International Journal of Applied Earth Observation and Geoinformation, 18, 472–479. https://doi.org/10.1016/j.jag.2011.09.018
- Bhattarai, K. R., & Vetaas, O. R. (2006). Can Rapoport’s rule explain tree species richness along the Himalayan elevation gradient, Nepal? Diversity & Distributions, 12(4), 373–378. https://doi.org/10.1111/j.1366-9516.2006.00244.x
- Bhattarai, K. R., Vetaas, O. R., & Grytnes, J. A. (2004). Fern species richness along a central Himalayan elevational gradient, Nepal. Journal of Biogeography, 31(3), 389–400. https://doi.org/10.1046/j.0305-0270.2003.01013.x
- Cairns, D., & Moen, J. (2004). Herbivory influences tree lines. Journal of Ecology, 92(1019), 1024. https://doi.org/10.1111/j.1365-2745.2004.00945.x
- Carlton, G. C., & Bazzaz, F. A. (1998). Regeneration of three sympatric birch species on experimental hurricane blowdown microsites. Ecological Monographs, 68(1), 99–120. https://doi.org/10.1890/0012-9615(1998)068[0099:ROTSBS]2.0.CO;2
- Catovsky, S., & Bazzaz, F. A. (2000). The role of resource interaction and seedling regeneration in maintaining a positive feedback in hemlock stands. Journal of Ecology, 88(1), 100–112. https://doi.org/10.1046/j.1365-2745.2000.00428.x
- Chawla, A., Yadav, P. K., Uniyal, S. K., Vats, S. K., Kumar, S., & Ahuja, P. S. (2012). Long-term ecological and biodiversity monitoring in the Western Himalaya using satellite remote sensing. Current Science, 102(8), 1143–1156.
- Chen, J., & Franklin, J. F. (1995). Growing-season microclimate gradients from clear cut edges into old-growth Douglas-fir forests. Ecological Applications, 5(1), 74–86. https://doi.org/10.2307/1942053
- Chen, J., Franklin, J. F., & Spies, T. A. (1993). Contrasting microclimatic patterns among clearcut, edge, and interior area of old-growth Douglas-fir forest. Agricultural and Forest Meteorology, 63(3/4), 219–237. https://doi.org/10.1016/0168-1923(93)90061-L
- Curtis, J. T. (1959). The vegetation of Wisconsin. An ordination of plant communities. University Wisconsin Press.
- Dimri, A. P., & Dash, S. K. (2011). Wintertime climatic trends in the western Himalayas. Climate Change, 111(3–4), 775–800. https://doi.org/10.1007/s10584-011-0201-y
- Gairola, S., Rawal, R. S., Todaria, N. P., & Bhatt, A. (2014). Population structure and regeneration patterns of tree species in climate-sensitive subalpine forests of Indian western Himalaya. Journal of Forestry Research, 25(2), 343–349. https://doi.org/10.1007/s11676-014-0463-0
- Gaur, R. D. (1999). Flora of the district Garhwal North West Himalaya with ethnobotanical notes. Transmedia.
- Grabherr, G., Gottfried, M., & Pauli, H. (1994). Climate effects on mountain plants. Nature, 369(6480), 448. https://doi.org/10.1038/369448a0
- Grace, J., Berninger, F., & Nagy, L. (2002). Impacts of climate change on the tree line. Annals of Botany, 90(4), 537–544. https://doi.org/10.1093/aob/mcf222
- Grime, J. P. (2001). Plant strategies, vegetation process, and ecosystem properties. Wiley.
- Hajra, P. K., & Balodi, B. (1995). Plant wealth of Nanda Devi biosphere reserve. Flora of India. Series-4. BSI, Dehra Dun.
- Holtmeier, F. K. (2009). Mountain timberlines: Ecology, patchiness, and dynamics. In Advances in global change research (pp. 36). Springer Science & Business Media.
- Holtmeier, F. K., & Broll, G. (2005). Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Global Ecology and Biogeography, 14(5), 395–410. https://doi.org/10.1111/j.1466-822X.2005.00168.x
- Holtmeier, F. K., & Broll, G. (2007). Treeline advance – Driving processes and adverse factors. Landscape Online, 1, 1–21. https://www.landscape-online.org/index.php/lo/issue/view/LO.200701
- Kala, C. P. (2005). Indigenous uses, population density and conservation of threatened medicinal plants in Protected Areas of the Indian Himalayas. Conservation Biology, 19(2), 368–378. https://doi.org/10.1111/j.1523-1739.2005.00602.x
- Kala, C. P., Rawat, G. S., & Uniyal, V. K. (1998). Ecology and conservation of the valley of flowers National Park, Garhwal Himalaya (Report). Wildlife Institute of India.
- Kanka, R., Kollar, J., & Barancok, P. (2005). Monitoring of climatic change impacts on alpine vegetation in the Tatry MTS – First approach. Ekologia (Bratislava), 24(4), 411–418. https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.629.5739&rep=rep1&type=pdf
- Khan, M. L., Rai, J. P. N., & Tripathi, R. S. (1987). Population structure of some tree species in disturbed and protected subtropical forests of north-east India. Acta Oecologica, 8(3), 247–255. https://scholar.google.com/scholar?hl=en&as_sdt=0%2C5&q=Population+structure+of+some+tree+species+in+disturbed+and+protected+subtropical+forests+of+north-east+India.+Acta+Oecologica%2C+8%283%29%2C+247%E2%80%93255&btnG=
- Kharkwal, G., Mehrotra, P., Rawat, Y. S., & Pangtey, Y. P. S. (2005). Phytodiversity and growth form in relation to altitudinal gradient in the Central Himalayan (Kumaun) region of India. Current Science, 89(5), 873–878. https://www.jstor.org/stable/24111035
- Korner, C. (1998). A re-assessment of high elevation treeline positions and their explanation. Oecologia, 115(4), 445–459. https://doi.org/10.1007/s004420050540
- Korner, C. (2003). Ein morphologiebedingter Wärmemangel bestimmt die Waldgrenze. In M. Winiger (Ed.), Carl Troll: Zeitumstände und Forschungsperspektiven. Kolloquium im Gedenken an den 100. Geburtstag von Carl Troll (pp. 114–119). Colloquium Geographicum 26.
- Korner, C. (2012). Alpine treelines: Functional ecology of the global high elevation tree limits. Springer.
- Kullman, L. (1998). Tree-limits and montane forests in the Swedish Scandes: Sensitive biomonitors of climate change and variability. Ambio, 27(4), 312–321. https://www.jstor.org/stable/4314741
- Kumar, P. (2012). Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using Maxent modelling: Limitations and challenges. Biodiversity and Conservation, 21(5), 1251–1266. https://doi.org/10.1007/s10531-012-0279-1
- Maletha, A.(2017). Ecological assessment of timberline vegetation with special reference to Betula utilis d.don forests in Nanda Devi Biosphere Reserve, Western Himalaya. Ph.D Thesis, Kumaun University.
- Misra, R. (1968). Ecology work book. Oxford and IBH Publishing Calcutta.
- Mori, A., & Takeda, H. (2004). Effects of undisturbed canopy structure on population structure and species coexistence in an old-growth subalpine forest in central Japan. Forest Ecology and Management, 200(1–3), 89–101. https://doi.org/10.1016/j.foreco.2004.06.036
- Naithani, B. D. (1984). Flora of Chamoli (Vols. 1-2). Botanical Survey of India. New Delhi, India.
- Nautiyal, S., Maikhuri, R. K., Rao, K. S., & Saxena, K. G. (2001). Medicinal plant resources in Nanda Devi Biosphere Reserve in the Central Himalayas. Journal of Herbs, Spices & Medicinal Plants, 8(4), 47–64. https://doi.org/10.1300/J044v08n04_06
- Pauli, H., Gottfried, M., Hohenwallner, D., Reiter, K., & Grabherr, G. (2004). The GLORIA field manual – Multi-summit approach. European Communities.
- Pauli, H., Gottfried, M., Reiter, K., & Klettner, C. G. (2007). Signals of range expansions and contractions of vascular plants in the high Alps: Observations (1994–2004) at the GLORIA master site Schrankogel, Tyrol, Austria. Global Change Biology, 13(1), 147–156. https://doi.org/10.1111/j.1365-2486.2006.01282.x
- Paulsen, J., & Körner, C. (2014). A climate-based model to predict potential treeline position around the globe. Alpine Botany, 124(1), 1–12. https://doi.org/10.1007/s00035-014-0124-0
- Phondani, P. C., Maikhuri, R. K., Negi, V. S., Rawat, L. S., Bahuguna, A., & Chamoli, K. P. (2010). Effect of provenance variation and temperature on seed germination of Himalayan Silver Birch (Betula utilis D.Don) in Central Himalaya. National Academy Science Letters, 33(7 and 8), 221–226. http://www.nasi.org.in
- Price, M. F., & Haslett, J. R. (1995). Climate change and mountain ecosystems. In N. J. R. Allan (Ed.), Mountains and risk: Current issues in environmental studies (pp. 73–97). Manohar Publishers and Distributors.
- Rai, I. D., Bharti, R. R., Adhikari, B. S., & Rawat, G. S. (2013). Structure and functioning of timberline vegetation in the western Himalaya: A case study. In N. Wu, G. S. Rawat, S. Joshi, M. Ismail, & E. Sharma (Eds.), High-altitude rangelands and their interfaces in the Hindu Kush Himalayas (pp. 91–107). Kathmandu.
- Samant, S. S. (1993). Diversity and status of plants in Nanda Devi Biosphere Reserve. In scientific and ecological expedition to Nanda Devi. (pp 54–85). New Delhi: Army Headquarters.
- Saxena, A. K., & Singh, J. S. (1984). Tree population structure of certain Himalayan forest associations and implications concerning their future composition. Vegetatio, 58(2), 61–69. https://doi.org/10.1007/BF00044928
- Schickhoff, U. (2005). The upper timberline in the Himalayas, Hindu Kush and Karakorum: A review of geographical and ecological aspects. In G. Broll & B. Keplin (Eds.), Mountain ecosystems. Studies in treeline ecology (pp. 275–354). Springer.
- Schickhoff, U., Bobrowski, M., Böhner, J., Bürzle, B., Chaudhary, R. P., Gerlitz, L., Heyken, H., Lange, J., Müller, M., Scholten, T., Schwab, N., & Wedegärtner, R. (2015). Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth System Dynamics, 6(1), 245–265. https://doi.org/10.5194/esd-6-245-2015
- Shankar, U. (2001). A case of high tree diversity in a sal (Shorea robusta) dominated lowland forest of Eastern Himalaya: Floristic composition, regeneration and conservation. Current Science, 81(7), 776–786. https://www.jstor.org/stable/24106397
- Shannon, C. E., & Weaver, W. (1949). The mathematical theory of communication. University of Illinois Press.
- Shrestha, B. B., Ghimire, B., Lekhak, H. D., & Jha, P. K. (2007). Regeneration of treeline birch (Betula utilis D. Don) forest in a trans-Himalayan dry valley in Central Nepal. Mountain Research and Development, 27(3), 259–267. https://doi.org/10.1659/mrdd.0784
- Silori, C. (2001). Biosphere reserve management in theory and practice: Case of Nanda Devi Biosphere Reserve, western Himalaya, India. Journal of International Wildlife Law & Policy, 4(3), 205–219. https://doi.org/10.1080/13880290109353987
- Singh, C. P., Panigrahy, S., Thapliyal, A., Kimothi, M. M., Soni, P., & Parihar, J. S. (2012). Monitoring the alpine treeline shift in parts of the Indian Himalayas using remote sensing. Current Science, 102(4), 559–562. https://www.jstor.org/stable/24084105
- Srivastava, S. (1999). Management plan for valley of flowers national park. Wildlife Preservation Organisation.
- Theurillat, J. P., & Guisan, A. (2007). Potential impacts of climate change on vegetation in the European Alps: A review. Climatic Change, 50(1/2), 77–109. https://doi.org/10.1023/A:1010632015572
- Ved, D. K., Kinhal, G. A., Ravikumar, K., Prabhakaran, V., Ghate, U., Vijaya Shankar, R., & Indresha, J. H. (2003). Conservation assessment and management prioritization for the medicinal plants of Jammu & Kashmir, Himachal Pradesh & Uttaranchal. Foundation for Revitalization of Local Health Traditions.
- Wang, T., Liang, Y. H., Ren, B., Yu, D., Ni, J., & Ma, K. P. (2004). Age structure of Picea schrenkiana forest along an altitudinal gradient in the central Tianshan Mountains, northwestern China. Forest Ecology and Management, 196(2–3), 267–274. https://doi.org/10.1016/j.foreco.2004.02.063
- Weiser, G., & Tuasz, M. (2007). Synopsis. In G. Wieser & M. Tuaszeds (Eds.), Trees at their upper limit: Tree life limitations at the alpine tree line (pp. 219–223). Springer.