
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
The forests of the central Himalayan region are characterized by the dominance of some populous and conspicuous tree species across altitudes that are crucial for ecosystem functions and services. An understanding of such forests with relation to species richness, diversity, structural pattern, and distribution is crucial in maintaining ecosystem stability. Considering this, the present study was undertaken on the four most common forest types dominated with Sal (Shorea robusta), Chir-pine (Pinus roxburghii), and Banj-oak (Quercus leucotrichophora), and mixed-oak (Q. lanuginosa and Q. floribunda) between 300 and 2250 m msl. Across the studied forest stands, we recorded a total of 110 plant species, belonging to 53 families. The most dominating life form was herbaceous (52%), followed by trees (27%), and shrubs (21%). Tree density was recorded maximum (884 ind. ha−1) at the mixed oak stand and minimum (652 ind. ha−1) recorded at the chir-pine forest; however, maximum (51.58 m2 ha−1) total basal area (TBA) was recorded in banj-oak forest and minimum (33.42 m2 ha−1) at mixed-oak forest stand. Tree density, seedling density, and TBA are significantly varied among the forest stands. The temperate oak forests comprised higher species richness and diversity at tree and shrub stratum. However, subtropical sal and chir-pine stand recorded higher regeneration. All stands exhibited high community dependence on forest resources that may increase the vulnerability of forest with respect to human interference, thus deserve immediate attention. Site-specific management and conservation measures have been suggested to provide opportunities for sustainable management of all forest stands.
1. Introduction
Himalayan forests represent complex and diversified ecosystems in the world due to variation in altitude, topography, geographical extent, and climatic conditions (C.M. Sharma et al., Citation2017; Singh & Singh, Citation1992; Sundriyal & Sharma, Citation1996). Spreading over an area of 2400 km in length and 300 km in width the Himalayan region contributes nearly 36% of the total forest area in India (ISFR, Citation2019). Other than maintaining significant biodiversity, the Himalayan forests support diverse ecosystem services to both upstream and downstream communities (Dhyani & Dhyani, Citation2016; Negi et al., Citation2018). The ecosystem services depend on species composition, diversity, richness, distribution pattern, and function, which is believed to be changing over time (Manral et al., Citation2018; Naidu & Kumar, Citation2016). Assessing species composition, diversity, and structure has been a major focus of ecological studies (Bhat et al., Citation2020). Such an evaluation is helpful in improving our understanding of forest ecosystems to improve conservation and management planning (Ahmad et al., Citation2020).
The dynamics of forest structure and function are influenced by various abiotic and biotic parameters (Chawla et al., Citation2008). The nature of forest communities largely depends on the ecological characteristics of sites, species richness, diversity, distribution, abundance, and regeneration status of species (Pandita et al., Citation2019). The population structure expressed in terms of tree girth class distribution provides valuable information on the reproductive capacity of any given stand (Joshi & Chandra, Citation2020; Sinha et al., Citation2018). Besides, an assessment of species diversity indices expresses the stability and sustainability of forest communities (Sarkar & Devi, Citation2014). All these components are essential for the preservation and maintenance of biodiversity. Thus, proper inventorying species composition of any forest stand is essential for its scientific management. Despite significant investigation on Himalayan forests (Bisht & Bhat, Citation2013; C.M. Sharma et al., Citation2017; Gairola et al., Citation2008; Kumar et al., Citation2021; Saxena & Singh, Citation1982; H. Singh et al., Citation2009; G. Singh & Rawat, Citation2012), the database needs a continuous update by undertaking further exploration to decipher various ecosystem properties of forest stands. It is argued that more investigations are needed for diverse forest types distributed across subtropical, temperate, and alpine environments covering evergreen broad-leaved, deciduous, and coniferous types because the majority of Himalayan forests are subject to intense biotic pressure (Bargali et al., Citation2013; C.M. Sharma et al., Citation2017).
Considering this, the present study was undertaken in four broad forest types in subtropical and temperate regions in the central Himalayan region comprising Sal (Shorea robusta), Chir-pine (Pinus roxburghii), and Banj-oak (Quercus leucotrichophora), and mixed-oak (Q. lanuginosa and Q. floribunda). The aim of this study was: (i) to analyze the species richness, diversity, and density of tree, shrub, and herb layers, (ii) to determine the natural regeneration status of the tree species, and (iii) to assess species distribution pattern and size class distribution across these stands. Shorea robusta (sal) dominates below 1000 m, whereas Pinus roxburghii (chir-pine) dominates dry hill slopes between 1000 and 1800 m elevations. Quercus leucotrichophora (banj-oak) dominates at approximately 1600–2200 m, while other oaks Quercus floribunda (tilonj-oak), Q. lanuginosa and Q. semecarpifolia (karshu-oak) dominate above 2000 up to subalpine conditions. We targeted dominating forest communities as such groups are most conspicuous and common, and researches over the past few decades suggest that they are key drivers of ecosystem functioning including species richness, diversity, structure, phenology, tree structure, biomass allocation, primary production, nutrient dynamics, litterfall, and turnover (Airi & Rawal, Citation2017; Gosain et al., Citation2015; Gurarni et al., Citation2010; Lal & Lodhiyal, Citation2016; Zobel & Singh, Citation1997). An effort was also made to compare the data of the present study with those of earlier studies. It is expected that the findings of the study will not only add to the existing quantitative information on such forests but also help to draw sound conclusions and actions to sustainably manage these forest communities.
2. Material and Method
2.1. Study area
The study area falls in Uttarakhand state, Central Himalaya under Nainital and Haldwani forest divisions of Nainital district in between 300 and 2250 m elevation above sea level (). Shorea robusta Gaertn (Sal) stand (Site-I) was located at Chorgaliya between 300 and 350 m elevations, Pinus roxburhii Sarg. (Chir-pine) stand (Site-II) was located at Baldiyakhan at 1450–1650 m, Quercus leucotrichophora (Banj-oak) stand (Site-III) at Kailakhan between 1750 and 1950 m, while Quercus lanuginosa Q. floribunda (Mixed-oak) stand (Site-IV) at Kilbury between 2050 and 2250 m elevation above sea level (). All stands fall under the reserve forest category that bears maximum protection. Site-I and site-II fall under the subtropical region while Site-III and site-IV are under the temperate zone. The mean annual rainfall during 2017–2019 was recorded at 1728.44 mm in the subtropical stands (Site-I and Site-II) and 1702.85 at temperate forest stands (Site-III and Site-IV). The maximum temperature at subtropical and temperate forest stands was recorded at 37.67 and 31.67°C, respectively; however, the minimum temperature was recorded 5.33°C at the temperate region and 7.67°C at subtropical forest stands ()). An examination of soil properties showed a sandy-loam texture and acidic nature across all sites (). The chemical contents of soil varied amongst sites. The soil organic carbon was recorded maximum at Banj-oak forest and minimum at Sal forest, while nitrogen content was assessed maximum at mixed-oak forest and minimum at Sal forest.
Figure 1. Location map of the study area in Uttarakhand, India.
Figure 2. Rainfall and temperature patterns of studied (a) subtropical and (b) temperate forest stands during study period 2017–2019.
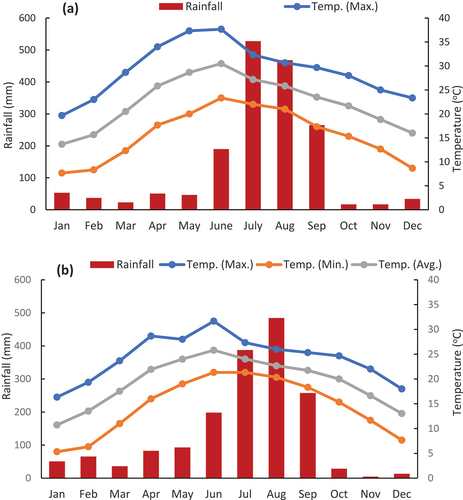
Table 1. Broad characteristics of investigated forest stands in Uttarakhand, Western Himalayan region.
2.2. Sampling designing data collection and analysis
Vegetation analysis
The phytosociological analysis was carried out from June 2017 to December 2017 using a random sampling procedure. To gather quantitative data of different forest layers (tree, shrub, and herbs) standard phytosociological methods were followed (Mueller-Dombois & Ellenberg, Citation1974). We randomly placed a total of 50 quadrates of 10 × 10 m size to assess trees and saplings, 50 sub-quadrates of 5 × 5 m size for seedlings, and 50 sub-quadrates of 1 × 1 m size for herbs at each forest stand. The circumference at breast height (CBH at 1.37 m from the ground) of all the tree individuals was recorded. All individuals with >30 cm CBH were categorized as trees, individuals with <30 cm CBH but height >20 cm as saplings, and individuals with <20 cm height were categorized as seedlings (Sundriyal & Sharma, Citation1996). The data were further analyzed for various phytosociological parameters. All plants were identified to species level by matching the specimens with the herbariums of G.B. Pant Institute and Kumaun University, Nainital, Uttarakhand.
CBH class distribution and regeneration pattern
Population structure and girth class distribution of all species in the forest stands during the same period was conducted using the quadrat method (Deb & Sundriyal, Citation2008). To develop the population structures of tree species, the girth class of each species was analyzed by grouping them into different CBH classes (A: 0–10 cm, B: 10–30 cm; C: 31–60 cm; D: 61–120 cm; E: 120–150 cm; F: 150–180 cm; G: 180–210 cm and H: >210 cm). The regeneration potential of species was analyzed following Uma Shankar (Citation2001).
Diversity indices and distribution pattern of species
The field data were also analyzed for various diversity parameters, i.e., Shannon diversity (H’; Shannon & Wiener, Citation1963), Species Evenness and concentration of dominance. The distribution pattern of species was determined by using the A: F value of species. The ratio indicates regular (<0.025), random (0.025 to 0.050), and contagious (>0.050) distribution pattern.
Shannon–Wiener diversity index was calculated as:
where H′ = Shannon’s diversity index, Ni = number of individuals of species belonging to the ith species and N = total number of individuals in the sample.
Concentration of Dominance was measured following Simpson (Citation1949):
where CD = Concentration of Dominance, Ni is the total number of individuals of species and N is the total number of individuals of all species.
The Species evenness index was calculated following Pielou (Citation1966):
where E = Species evenness index, H′ = Shannon’s diversity index, and S = Total number of species
Statistical analysis
The phytosociological data were analyzed by using Microsoft excel 2010. All the statistical analysis (ANOVA) between different parameter was conducted following IBM-SPSS, 20.
3. Results
3.1. Site characteristics and species richness
The broad characteristics of different investigated forest stands are presented in . The forest stands were distributed between 300 and 2250 m elevations above sea level that covered nearly 62% of the total forest cover of the region. A total of 110 plant species varying from 53 families were recorded across all stands (Annexure 1). Of all species, the maximum represented herbaceous habit (52%), followed by the tree (27%), and shrub (21%) habits. It was interesting to note that total species richness increased with altitude with 19, 45, 46, and 50 species at Sal, Chir-pine, Banj-oak, and mixed-oak stands, respectively. The distribution of species in the tree, shrub, and herbaceous habits was recorded 8, 6, and 5, respectively, at Sal stand, 11, 9, and 25 at Chir-pine stand 12, 9, and 25 at Banj-oak stand, and 13, 12, and 25, respectively, at Mixed-oak stand ().
Table 2. Structural attributes of the forest stands in Uttarakhand, Western Himalaya.
3.2. Stand density and total basal cover
The tree density ranged between 652 and 884 ha−1, being maximum (884 ha−1) at mixed-oak stand, followed by Sal (756 ha−1), Chir pine (672 ha−1), and minimum at Banj-oak stand (652 ha−1; ). The sapling density ranged between 140 ha−1 (Chir-pine forest) and 368 ha−1 (Mixed-oak stand), while the seedling density varied from 174 (Banj-oak forest) to 3086 ha−1 (Sal forest). The total basal area (TBA) was maximum for Banj-oak stand (51.58 m2 ha−1), followed by Sal (47.52 m2 ha−1), Chir-pine (43.64 m2 ha−1), and minimum for Mixed-oak forest (33.42 m2 ha−1). The shrub density was recorded as 1700 to 7176 ha−1, maximum at Sal forest followed by Chir-pine, Mixed-oak, and Banj-oak forest for the herbaceous layer, maximum density was recorded in Banj-oak forest (234.80 m−2), followed by Mixed-oak stands (152.52 m−2), Chir-pine forest (119.85 m−2), and Sal forest (27.37 m−2).
The relative density (RD) of individual species exhibited that at Sal stand, Shorea robusta (RD = 51.52%) and Mallotus philippensis (RD = 36.77%) were dominant and co-dominant status at tree layer, and Clerodendron infortunatum (RD = 46.82%) dominated at shrub layer. At the Chir-pine stand, Pinus roxburghii (RD = 80.36%) exhibited a clear dominance at tree layer, while Eupatorium adenophorum (RD = 51.53%) at shrub layer. Banj-oak stand dominated by Q. lecucotrichophora (RD = 63.19%) at tree stratum and Daphne cannabina (RD = 20.71%) at shrub stratum, however, Mixed-oak stand dominated by Q. lanuginosa (RD = 36.65%) and co-dominated by Q. floribunda (RD = 29.41%) at tree layer, Daphne cannabina (RD = 23.29%) at shrub stratum and Arundinaria falcata (RD = 11.42%) at herb stratum (Annexure 1). The ANOVA analysis between forest stands showed significant variations among seedlings density (F = 46.05; p < 0.01), shrub density (F = 77.02; p < 0.01), herb density (F = 25.23; p < 0.01), total basal area (F = 4.45; p < 0.01), and tree density (F = 3.82; p < 0.05), although the sapling density among forest stands not varied significantly ().
Table 3. ANOVA analysis (F and p value) of vegetation parameter among study sites.
3.3. Species diversity and evenness
The species diversity (H’) ranged between 1.10 and 2.31 for tree layer, 2.18 to 3.14 for shrub layer, and 1.49 to 2.18 for herb layer among different investigated stands (). The species diversity was recorded maximum at mixed oak forest at tree and shrub layer. The concentration of dominance (CD) ranged between 0.24 and 0.66 for the tree layer, 0.11 to 0.30 for the shrub layer, and 0.075 to 0.42 for the herb layer. The species evenness ranged between 0.46 to 0.90, 0.81 to 1.34, and 0.52 to 0.85 for tree, shrub, and herb stratum, respectively (). The maximum evenness was recorded at Mixed-oak stand for tree and shrub layer. The species diversity, evenness, and concentration of dominance of tree were not varied significantly (p = 0.92, 0.08, and 0.09, respectively) among the investigated forest stands.
Table 4. Diversity, dominance and evenness indices for tree, sapling and seedlings stratum for different investigated forest stands.
3.4. Species distribution
The species distribution pattern varied between stands and vegetation layers (). Trees and herbs exhibited a dominance of contagious distribution. Shrubs species showed regular distribution at Sal and mixed-oak stands, and contagious distribution at other stands. At the sal forest, 37.50%, and 62.50% species exhibited random and contagious distribution, respectively, at tree layer, 83.33% and 16.67% species showed random and contagious distribution, respectively, at shrub layer. At Chir-pine stand, 81.82% species showed contagious and 18.82% showed regular distribution in tree layer, and 44.44% species showed random and 55.56% species showed contagious distribution in the shrub layer. At Banj-oak stand, only 8.33% species showed random, 16.67% regular, and 75% species showed contagious distributions in tree layer, while 20% regular, 40% random, and 40% contagious in the shrub layer, However, in the mixed-oak stand 38.46% species showed regular distribution and 61.54% contagious distribution in tree layer, while at shrub layer 36.36% species showed random, 36.36% regular, and 27% species showed contagious distribution ().
Table 5. Distribution pattern of tree, shrub and herb species across forest stands.
3.5. Size class distribution and regeneration pattern
The Sal and Chir-pine stands exhibited a higher density of lower girth class individuals (seedling), although Banj-oak and mixed-oak stands showed a dominance of young trees (; I–IV). At the stands level, respective distribution of seedling, sapling, and tree individuals was recorded 17%, 24%, and 59% at Mixed-oak stand, 17%, 20%, and 67% at Banj-oak stand, 75%, 7%, and 18% at Sal stand, and 68%, 5% and 27% at Chir-pine stand. The study revealed that only 6% of species showed good regeneration while 50% had fair regeneration (). It was also interesting to note that 30% of species were not regenerating at all stands while 14% of species were new to these sites.
Figure 3. (I–IV): Girth class structure of studies forest stands in the Western Himalayan region (CBH classes represents A: 0–10 cm, B: 10–30 cm; C: 31–60 cm; D: 61–120 cm; E: 120–150 cm; F: 150–180 cm; G: 180–210 cm and H: >210 cm).
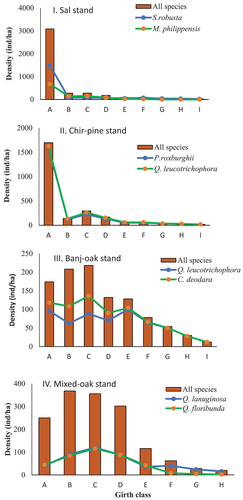
Table 6. Regeneration status of species across forest stands.
3.6. Family composition of species
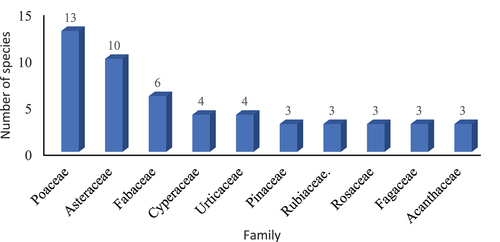
Of the total recorded plant families (53), Poaceae was the most dominant family with 13 species, followed by Asteraceae with 10 species (). Interestingly, 32 families were monotypic and characterized only by single species, while eight families comprised two species, six families three species each, and three families with more than four species.
Figure 4. Dominant plant families (Top 10) of selected forest stands in the Central Himalaya
3.7. Comparison with other Himalayan forests
A comparison of the present study with the tree density and basal area of other Himalayan forests is presented in . Our values of tree density and basal area for the four investigated stands broadly fall within the range for other similar forests.
Table 7. Comparison of present study (density and basal area) with other forest stands of the Himalayan region.
4. Discussion
The study area comprised four dominant forest communities covering an altitudinal range of 300 to 2250 m above sea level, which also coincides with the major human habitation zone in the region. These communities together formed nearly 62% of the total forest cover of the region. A total of 110 plant species varying from 53 families were recorded across all stands, which is significant. The herbaceous life form was most dominant followed by the tree, and shrub habits. The species richness increased with altitude, which can be attributed to variation in climatic, edaphic, and topographic conditions (Singh & Singh, Citation1984). The species richness value of the present investigation reported from 19 to 47 (8–13 for the tree, 6–12 for shrub, and 5–26 for herb), which is higher as compared to the value 13–40 (5–9 for the tree, 5–8 for shrub and 3–22 for herb) reported in the subtropical and temperate forest stands in the western Himalaya region at similar elevation (300–2200 m) zone (Rana et al., Citation1989; Rawat & Singh, Citation1988). Also, species richness of 66 species (12 trees, 19 shrubs, and 34 herbs) has been reported in the temperate forest region of Garhwal Himalaya (Gairola et al., Citation2011), which is much lower as compared to the present study. Low species richness at Sal stand can be attributed to frequent removal of understory vegetation by the Forest department to improve the natural regeneration. Although, higher species richness in the temperate areas can be attributed to better environmental conditions, such as optimal moisture, temperature, rainfall, and shady conditions that support better plant growth (N. Sharma et al., Citation2019).
The Sal stand showed a clear dominance of Shorea robusta (52%), Chir-pine stand of Pinus roxburghii (80%), Banj-oak stand of Q. lecucotrichophora (63%), and Mixed-oak stand of Q. lanuginosa (37%) and Q. floribunda (29%). Therefore, all stands were ideally identified with the name of the dominant species. A dominant species is the most populous species that is found most conspicuously in a particular ecosystem and generally comprises the maximum density and biomass in an ecosystem (Singh & Singh, Citation1992; Sundriyal & Sharma, Citation1996). A comparison of tree density recorded in this study with those reported earlier revealed that Sal stand exhibited either a comparable or higher density value than reported by others (Mittal et al., Citation2020; Rana et al., Citation1989). Similarly, the tree density value recorded for Chir-pine stand is higher than the value (138–575 ind. ha−1) reported by others (Kumar et al., Citation2021; Mittal et al., Citation2020). In the case of Banj-oak and mixed-oak forests, present values are more or less comparable with the value reported by various worker in the similar forests’ types (Kumar & Ram, Citation2005; Pande et al., Citation2014; H. Singh et al., Citation2009).
The reported total basal area in present study in the Sal forest is higher than other similar forests (range 37.37 to 41 m2 ha−1) (Bohra et al., Citation2010). The reported basal area of Chir-pine stand is comparable with the value (25 to 47.2 m2 ha−1) reported by Chaturvedi & Singh, and lower to the value (43–67.14 m2 ha−1) reported in the other Chir-pine forest of western Himalaya (Mittal et al., Citation2020). The total basal area value has been reported as 30.69 to 36.49 m2 ha−1 for oak forests of Himalayan region (Rana et al., Citation1989; Rikhari et al., Citation1991; Saxena & Singh, Citation1982), which clearly reveals that our values of the total basal area are toward the higher side of those reported for other similar forests in the central Himalaya. The diversity index for tree and shrub layers was estimated maximum for mixed-oak stand, while for herb layer it was maximum at Chir-pine stand. The values of species diversity in the present study (1.10–3.14) are well comparable with the reported range 1.16 and 3.40 for different Himalayan forests (Dar et al., Citation2017; Joshi et al., Citation2021; V. Singh et al., Citation2015). The concentration of dominance (CD) ranged between 0.24 and 0.66 for the tree layer, 0.11 to 0.30 for the shrub layer, and 0.075 to 0.42 for the herb layer, these values are comparable to the reported range for other Himalayan forests (Singh & Singh, Citation1992). The evenness values in the present study at tree layer (0.46–0.90) are more or less comparable with the values (0.37–0.91) reported in the other similar forest stands of this region (Sarkar & Devi, Citation2014).
In this study, the maximum species across all three strata (i.e., trees, shrubs, and herbs) exhibited a contagious distribution pattern. This is supported by the finding of Gairola et al. (Citation2011), which shows the contagious distribution is the most common distribution pattern in the temperate forest stands in the Garhwal Himalaya. Absence of regular distribution of species may risk their population in the near future (Shankar, Citation2001). The proportion of DBH in a given population (seedling, sapling, small tree, and large tree) is essential to the prediction of the future status of forest stands (Malik & Bhatt, Citation2016; Sundriyal & Sharma, Citation1996). In the present study, Sal and Chir-pine stands illustrated reverse “J” distribution having maximum individuals in lower girth class and the number decreased in middle and higher girth class thus indicating high regenerative potential, however in the Banj-oak and mixed-oak stands showed a dominance of mature tree, this indicates poor regeneration potential of tree. In this investigations, only 6% of species showed good regeneration and 30% of species were not regenerating, which is a matter of concern. Although 50% of species exhibited had fair regeneration, while 14% of species were new to these forest stands. The dominant and co-dominant species contribute a major part to regeneration at each stand and the presence of new individuals in seedling and sapling stages indicates emergence of new species in these forest stands, which can be attributed the successional status of a given forest stand (Deb & Sundriyal, Citation2008). As such, the girth class structure revealed that these forest stands will exhibit a similar structure of dominant and co-dominant species in near future as well. Although there is a need for proper management and conservation strategies for the maintenance and sustainability of these stands to manage tree species diversity and regeneration.
Conclusions
High dependence on forest resources has increased its vulnerability with respect to human interference. Therefore, an understanding of forest dynamics is crucial for sustainable utilization and management of resources. This study investigated the dynamics of four contrasting forest communities in the Central Himalayan region (namely, Sal, Shorea robusta; Chir-pine, Pinus roxburghii; Banj-oak, Quercus leucotrichophora, and mixed-oak, Q. lanuginosa and Q. floribunda) with relation to species richness, stand density, total basal cover, species diversity, distribution pattern, species girth class distribution, and regeneration status that varied among stands. The study exhibited that temperate oak forest stands were better in terms of species richness and diversity than the subtropical Sal and Chir-pine forests. However, the poor regeneration in these temperate forests stands is a matter of concern. Contrarily, an improved regeneration in Sal and Chir-pine forests explains that these forests can be managed effectively if biotic pressure is reduced. Site-specific variations in species richness, diversity, density, basal area, and regeneration of different forest stands may be attributed to altitudinal and environmental variations, species composition, age structure, successional stage of the forest, and degree of disturbance (Pala et al., Citation2013; Sarkar & Devi, Citation2014; N. Sharma et al., Citation2019).
The data have significant implications for developing a long-term strategy for monitoring, conservation, and management of forest stands. First, forest managers need to develop a better scientific understanding of forest dynamics and various anthropogenic factors affecting all these forest stands. The quantitative analysis undertaken in this study may provide baseline information for further formulating conservation and management strategies of these forest stands. Second, supplement natural reproduction underplanting with seedlings of desirable species can improvize the species status in various forest stands. Such plantations should use dominant, co-dominant, native, and other desirable species. The plantation should be done up to a desired level of stocking. Third, site-specific stand-management options are highly desirable. For example, the Sal stand is maintained for commercial timber production. In this stand, there were many regenerating individuals, however only a few reaches to tree stratum. Local people exploit the stand for meeting their diverse needs that desire to be addressed immediately. The stand also needs proper thinning measures to increase the growth rate of the remaining trees. Chir-pine stand is frequently used for the collection of timber, firewood, charcoal making, and resin collection. The stand is also used for fodder collection and grazing purposes. In view of the highly inflammable nature of Chir-pine needles, such stands are the main causes of forest fires. It greatly impacts forest regeneration. The stand can be planted with broad-leaved species. Besides, the chir-pine needles should be used for some other purposes such as bio briquettes, vermicompost, paper making, boards, panels, and tiles, etc. Oak species are highly valued in the local system in the state. Oak leaves are a good source of fodder to animals during winter months when there is a dearth of fodder. It is amongst the best firewood species. Its leaves are used for animal bedding in winters and such material is used for agriculture field mulching. Oak stands also provide important habitat for a large variety of plants and animals. Fortunately, communities have a good attitude to conserve oak stands, which should be admired and strengthened further. Fourth, although all these forests had a legal status of reserve forests that are meant to meet timber, revenue, and biodiversity conservation. However, nearby communities use these forests for the collection of firewood, fodder, litter, and grazing purposes, which is in conflict with the interest of the Forest department. To promote tree species, regeneration and natural recruitment forests need to be protected from all kinds of biotic interferences. However, strict enforcement of forest legislation may lead to conflicts between the interests of local people and the government; therefore, site-specific strategies for managing each forest stand by involving local communities in forest management is highly desirable. Using strand-specific management options in combination with other scientific approaches may lead to achieving sustainable management of all forest stands.
Supplemental Material
Download MS Word (37.8 KB)Acknowledgments
We are highly thankful to the Ministry of Environment, Forest and climate change (MoEF&CC) for providing financial support, and Director GB Pant National Institute of Himalayan Environment (GBP-NIHE) for providing necessary facilities.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary Material
Supplemental data for this article can be accessed here.
Additional information
Funding
References
- Adhikari, B., Kapkoti, B., Lodhiyal, N., & Lodhiyal, L. S., et al. (2017). Structure and regeneration of Sal (Shorea robusta Gaertn f.) forests in Shiwalik Region of Kumaun Himalaya, India. Indian Journal of Forestry, 40(1), 1–8. https://doi.org/10.54207/bsmps1000-2017-0N83L0
- Ahmad, M., Uniyal, S. K., Batish, D. R., Singh, H. P., Jaryan, V., Rathee, S., Sharma, P., & Kohli, R. K., et al. (2020). Patterns of plant communities along vertical gradient in Dhauladhar Mountains in Lesser Himalayas in North Western India. Science of the Total Environment, 716, 136919. https://doi.org/10.1016/j.scitotenv.2020.136919
- Airi, S., & Rawal, R. S. (2017). Patterns of vegetation composition across levels of canopy disturbance in temperate forests of west Himalaya, India. Biodiversity Research and Conservation, 45(1), 27–33. https://doi.org/10.1515/biorc-2017-0004
- Bargali, K., Bisht, P., Khan, A., & Rawat, Y. S., et al. (2013). Diversity and regeneration status of tree species at Nainital Catchment, Uttarakhand, India. International Journal of Biodiversity and Conservation, 5(5), 270–280. http://doi.org/10.5897/IJBC10.099
- Bhat, J. A., Kumar, M., Negi, A. K., Todaria, N. P., Malik, Z. A., Pala, N. A., & Shukla, G., et al. (2020). Species diversity of woody vegetation along altitudinal gradient of the Western Himalayas. Global Ecology and Conservation, 24, e01302. https://doi.org/10.1016/j.gecco.2020.e01302
- Bisht, A. S., & Bhat, A. B. (2013). Vegetational structure and plant diversity relation in a sub-alpine region of Garhwal Himalaya, Uttarakhnad India. African Journal of Plant Science, 7(8), 401–406. https://doi.org/10.5897/AJPS11.013
- Bohra, C. S., Lodhiyal, L. S., & Lodhiyal, N. (2010). Forest stand structure of Shiwalik region of Nainital district along an altitudinal gradient in Indian central Himalaya. New York Science Journal, 3(12), 82–90.
- Chaturvedi, O. P., & Singh, J. S. (1987). The structure and function of pine forest in Central Himalaya. I. Dry matter dynamics. Annals of Botany, 60(3), 237–252. https://doi.org/10.1093/oxfordjournals.aob.a087442
- Chawla, A., Rajkumar, S., Singh, K. N., Lal, B., Singh, R. D., & Thukral, A. K., et al. (2008). Plant species diversity along an altitudinal gradient of Bhabha Valley in western Himalaya. Journal of Mountain Science, 5(2), 157–177. https://doi.org/10.1007/s11629-008-0079-y
- Dar, J. A., Rather, M. Y., Subashree, K., Sundarapandian, S., & Khan, M. L., et al. (2017). Distribution patterns of tree, understorey, and detritus biomass in coniferous and broad-leaved forests of Western Himalaya, India. Journal of Sustainable Forestry, 36(8), 787–805. https://doi.org/10.1080/10549811.2017.1363055
- Deb, P., & Sundriyal, R. C. (2008). Tree regeneration and seedling survival patterns in old-growth lowland tropical rainforest in Namdapha National Park, north-east India. Forest Ecology and Management, 255(12), 3995–4006. https://doi.org/10.1016/j.foreco.2008.03.046
- Devlal, R., & Sharma, N. (2008). Altitudinal changes in dominance-diversity and species richness of tree species in a temperate forest of Garhwal Himalaya. Life Science Journal, 5(2), 53–57.
- Dhanai, C. S., Panwar, V. P., & Sharma, C. M., et al. (2000). Effect of aspect and soil on the structure of quercus leucotrichophora natural stands in Western Himalaya. Indian Journal of Forestry, 23(3), 349–356.
- Dhyani, S., & Dhyani, D. (2016). Significance of provisioning ecosystem services from moist temperate forest ecosystems: Lessons from upper Kedarnath valley, Garhwal, India. Energy, Ecology and Environment, 1(2), 109–121. https://doi.org/10.1007/s40974-016-0008-9
- Gairola, S., Rawal, R. S., & Todaria, N. P., et al. (2008). Forest vegetation patterns along an altitudinal gradient in sub-alpine zone of west Himalaya, India. African Journal of Plant Science, 2(6), 42–48.
- Gairola, S., Sharma, C. M., Suyal, S., & Ghildiyal, S. K., et al. (2011). Composition and diversity of five major forest types in moist temperate climate of the western Himalayas. Forestry Studies in China, 13(2), 139. https://doi.org/10.1007/s11632-011-0207-6
- Gosain, B. G., Negi, G. C. S., Dhyani, P. P., Bargali, S. S., & Saxena, R., et al. (2015). Ecosystem services of forests: Carbon stock in vegetation and soil components in a watershed of Kumaun Himalaya, India. International Journal of Ecology and Environmental Science, 41(3–4), 177–188.
- Gurarni, D., Arya, N., Yadava, A., & Ram, J., et al. (2010). Studies on plant biodiversity of pure Pinus roxburghii Sarg. forest and mixed pine-oak forest in Uttarakhand Himalaya. New York Science Journal, 3(8), 1–5.
- ISFR (2019). India State of Forest Report 2019. Forest Survey of India, Ministry of Environment, Forest & Climate Change, Government of India.
- Joshi, V. C., & Chandra, N. (2020). Pattern of diversity and regeneration potential along altitudinal gradient in selected forest stands of Kumaun Himalaya. Indian Forester, 146(4), 301–305. http://doi.org/10.36808/if/2020/v146i4/146911
- Joshi, V. C., Sundriyal, R. C., & Arya, D., et al. (2021). Forest floor diversity, distribution and biomass pattern of oak and Chir-pine forest in the Indian Western Himalaya. Indian Journal of Ecology, 48(1), 232–237.
- Kapkoti, B., Adhikari, B., Lodhiyal, N., & Lodhiyal, L. S., et al. (2016). Structure and diversity of Sal forests in government and community management systems in Kumaun region of Central Himalaya. Current World Environment, 11(1), 126. https://doi.org/10.12944/CWE.11.1.16
- Kumar, A., & Ram, J. (2005). Anthropogenic disturbances and plant biodiversity in forests of Uttaranchal, central Himalaya. Biodiversity and Conservation, 14(2), 309–331. https://doi.org/10.1007/s10531-004-5047-4
- Kumar, M., Kumar, A., Kumar, R., Konsam, B., Pala, N. A., & Bhat, J. A., et al. (2021). Carbon stock potential in Pinus roxburghii forests of Indian Himalayan regions. Environment, Development and Sustainability, 23, 12463–12478. https://doi.org/10.1007/s10668-020-01178-y
- Kusumlata, B. N. (1991). Quantitative analysis and regeneration potential of moist temperate forest in Garhwal Himalaya. Indian Journal of Forestry, 14(2), 98–106.
- Lal, B., & Lodhiyal, L. S. (2016). stand structure, productivity and carbon sequestration potential of oak dominated forests in Kumaun Himalaya. Current World Environment, 11(2), 466. https://doi.org/10.12944/CWE.11.2.15
- Malik, Z. A., & Bhatt, A. B. (2016). Regeneration status of tree species and survival of their seedlings in Kedarnath Wildlife Sanctuary and its adjoining areas in Western Himalaya, India. Tropical Ecology, 57(4), 677–690.
- Manral, U., Badola, R., & Hussain, S. A., et al. (2018). Forest composition and structure under various disturbance regimes in the Alaknanda River Basin, Western Himalaya. Mountain Research and Development, 37(3), 310–322. https://doi.org/10.1659/MRD-JOURNAL-D-16-00109.1
- Mittal, A., Singh, N., & Tewari, A., et al. (2020). Quantitative analysis and regeneration status of forest trees species in Kumaun Central Himalaya. Indian Journal of Ecology, 47(2), 507–513.
- Mueller-Dombois, E., & Ellenberg, H. (1974). Aims and methods of vegetation ecology. John Wiley & Sons.
- Naidu, M. T., & Kumar, O. A. (2016). Tree diversity, stand structure, and community composition of tropical forests in Eastern Ghats of Andhra Pradesh, India. Journal of Asia-Pacific Biodiversity, 9(3), 328–334. https://doi.org/10.1016/j.japb.2016.03.019
- Negi, V. S., Joshi, B. C., Pathak, R., Rawal, R. S., & Sekar, K. C., et al. (2018). Assessment of fuelwood diversity and consumption patterns in cold desert part of Indian Himalaya: Implication for conservation and quality of life. Journal of Cleaner Production, 196, 23–31. https://doi.org/10.1016/j.jclepro.2018.05.237
- Pala, N. A., Negi, A. K., Gokhale, Y., Aziem, S., Vikrant, K. K., & Todaria, N. P., et al. (2013). Carbon stock estimation for tree species of Sem Mukhem sacred forest in Garhwal Himalaya, India. Journal of Forestry Research, 24(3), 457–460. https://doi.org/10.1007/s11676-013-0341-1
- Pande, R., Bargali, K., & Pande, N., et al. (2014). Impacts of disturbance on the population structure and regeneration status of tree species in a Central Himalayan Mixed-Oak Forest, India. Taiwan Journal of Forest Science, 29(3), 179–192.
- Pandita, S., Kumar, V., & Dutt, H. C., et al. (2019). Environmental variables vis-a-vis distribution of herbaceous tracheophytes on northern sub-slopes in Western Himalayan ecotone. Ecological Processes, 8(1), 1–9. https://doi.org/10.1186/s13717-019-0200-x
- Pielou, E. C. (1966). Shannon’s formula as a measure of specific diversity: Its use and misuse. The American Naturalist, 100(914), 463–465. https://doi.org/10.1086/282439
- Rana, B. S., Singh, S. P., & Singh, R. P., et al. (1989). Biomass and net primary productivity in Central Himalayan forests along an altitudinal gradient. Forest Ecology and Management, 27(3–4), 199–218. https://doi.org/10.1016/0378-1127(89)90107-2
- Rawat, Y. S., & Singh, J. S. (1988). Structure and function of oak forests in central Himalaya. I. Dry matter dynamics. Annals of Botany, 62(4), 397–411. https://doi.org/10.1093/oxfordjournals.aob.a087673
- Rikhari, H. C., Tewari, J. C., Rana, B. S., & Sharma, S., et al. (1991). Woody vegetation and regeneration status in a mixed oak forest of Kumaun Himalaya. Indian Forester, 117(4), 274–283.
- Sarkar, M., & Devi, A. (2014). Assessment of diversity, population structure and regeneration status of tree species in Hollongapar Gibbon Wildlife Sanctuary, Assam, Northeast India. Tropical Plant Research, 1(2), 26–36.
- Saxena, A. K., & Singh, J. S. (1982). A phytosociological analysis of woody species in forest communities of a part of Kumaun Himalaya. Vegetatio, 50(1), 3–22. https://doi.org/10.1007/BF00120674
- Shankar, U. (2001). A case of high tree diversity in a sal (Shorea robusta)-dominated lowland forest of Eastern Himalaya: floristic composition, regeneration and conservation. Current Science, 81(7), 776–786.
- Shannon, C. E., & Wiener, W. (1963). The mathematical theory of communities. University of Illinois Press.
- Sharma, C. M., Mishra, A. K., Tiwari, O. P., Krishan, R., & Rana, Y. S., et al. (2017). Effect of altitudinal gradients on forest structure and composition on ridge tops in Garhwal Himalaya. Energy, Ecology and Environment, 2(6), 404–417. https://doi.org/10.1007/s40974-017-0067-6
- Sharma, N., Behera, M. D., Das, A. P., & Panda, R. M., et al. (2019). Plant richness pattern in an elevation gradient in the Eastern Himalaya. Biodiversity and Conservation, 28(8–9), 2085–2104. https://doi.org/10.1007/s10531-019-01699-7
- Simpson, E. H. (1949). Measurement of diversity. nature, 163(4148), 688. https://doi.org/10.1038/163688a0
- Singh, G., & Rawat, G. S. (2012). Quantitative analysis of tree species diversity in different oak (Quercus spp.) dominated forests in Garhwal Himalaya, India. Notulae Scientia Biologicae, 4(4), 132–140. https://doi.org/10.15835/nsb448200
- Singh, H., Kumar, M., & Sheikh, M. A., et al. (2009). Distribution pattern of oak and pine along altitudinal gradients in Garhwal Himalaya. Nature and Science, 7(11), 81–85.
- Singh, J. S., & Singh, S. P. (1984). An integrated ecological study of eastern Kumaun Himalaya with emphasis on natural resources Kumaun University, Nainital”. Final Report (HCS/DST/187/76) vols 1-3 (Unpublished).
- Singh, J. S., & Singh, S. P. (1992). Forest of Himalaya, Structure, Functioning and impact of Man. Gyanodaya Prakashan.
- Singh, N., Tamta, K., Tewari, A., & Ram, J., et al. (2014). Studies on vegetational analysis and regeneration status of Pinus roxburghii, Roxb. and Quercus leucotrichophora forests of Nainital Forest Division. Global Journal of Science Frontier Research, 14(3), 41–47.
- Singh, V., Chauhan, D. S., & DaSguPTa, S., et al. (2015). Vegetation status and plant diversity of Chandrabadani Oak forest along anthropogenic disturbance gradient in Garhwal Himalaya, Uttarakhand, India. Journal of Global Agriculture and Ecology, 3(1), 26–37.
- Sinha, S., Badola, H. K., Chhetri, B., Gaira, K. S., Lepcha, J., & Dhyani, P. P., et al. (2018). Effect of altitude and climate in shaping the forest compositions of Singalila National Park in Khangchendzonga Landscape, Eastern Himalaya, India. Journal of Asia-Pacific Biodiversity, 11(2), 267–275. https://doi.org/10.1016/j.japb.2018.01.012
- Sundriyal, R. C., & Sharma, E. (1996). Anthropogenic pressure on tree structure and biomass in the temperate forest of Mamlay watershed in Sikkim. Forest Ecology and Management, 81(1–3), 113–134. https://doi.org/10.1016/0378-1127(95)03657-1
- Verma, A. K., & Garkoti, S. C. (2019). Population structure, soil characteristics and carbon stock of the regenerating Banj-oak forests in Almora, Central Himalaya. Forest Science and Technology, 15(3), 117–127. https://doi.org/10.1080/21580103.2019.1620135
- Zobel, D. B., & Singh, S. P. (1997). Himalayan forests and ecological generalizations: forests in the Himalaya differ significantly from both tropical and temperate forests. Bio Science, 47(11), 735–745.