
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Understanding species richness and diversity patterns and their governing factors in less-to-unexplored regions across Himalaya provide invaluable insights into exploring drivers which shape as well as influence plant community structures. The present investigation explores plant species richness and diversity patterns across different growth forms and its association with environmental parameters along altitudinal gradient (3200 m-4800 m) in alpine regions of west Himalaya, India. A total of 265 plant taxa were documented from study area with higher proportion of herbs (212), followed by shrubs (44) and trees (9). Species richness, diversity, and density patterns were estimated for each growth form along altitude gradients using polynomial regression and an apparent monotonically decreasing trend (p < 0.05) was seen across transects, with highest values for herbs. Beta diversity, estimated for each transect, was low in Darma for herbs exhibiting high species packaging and homogenous composition, and high in Mana showing more scope for occurrence of rare/occasional herbs. Four major distinct altitudinal zones were identified for Uttarakhand alpines using cluster dendrogram, i.e., 3200 to 3500 m, 3600 to 3900 m, 4000 to 4500 m and 4600 to 4800 m with respect to their vegetation composition. NMDS of combined dataset along altitude gradient across transects also exhibited proximity among lower altitudesof transects with similar species composition (like Anaphalis, Danthonia, Geranium, Pedicularis, Potentilla), while high-altitude plots were scattered towards both ends of axesinhabiting specialized plant species (like Gentiana, Nardostachys, Saussurea, Sedum, Swertia). The relationship between vegetation variables (richness, diversity and density) and climate variables was modelled using Pearson’s correlation (P < 0.001) and temperature, precipitation, and solar radiation exhibited positive correlation, while windspeed showed negative correlation. Relative effect of climatic parameters on species composition, analysed by CCA, showed strongest influence of precipitation in vegetation zones with high axes correlation, followed by temperature, isothermality and wind speed, while influence solar radiation was lowest. Thus, under the current climate change scenario, any change in these factors may alter the composition of these high-altitude area and threaten the unique flora as well as the fauna dependent on it. Hence, any effort made towards conservation would eventually benefit a significant proportion of Himalayan biodiversity.
Significance statement
The alpine regions of Himalaya have rich plant diversity and vary along with elevational gradient. Environmental parameters have played significant role on plant distribution and diversity. Studies on relation between the plant diversity and environmental factors are limited. Keeping in view the study has been conducted to analyse the plant diversity with possible environmental parameters. The observed variation in temperature and precipitation patterns along altitude gradient significantly correlates with the species richness and diversity patterns. The linear correlation states altitude as the variable which best explains the variations in richness and diversity patterns.
Introduction
Mountains are recognized as biologically diverse ecosystems supporting high proportions of flora and fauna and harbouring a large number of protected areas (Mallen-Cooper & Pickering, Citation2008). Owing to the different topography, micro and macroclimatic conditions, mountain ecosystems consist of a range of habitats inhabited by variable life-forms. Recent decades have witnessed a noticeable increment in biodiversity research especially in these cold biomes at high elevations. Several studies have demonstrated characteristic relationships between altitude, species richness patterns and composition, varying from hump-shaped with maximum richness in mid-altitudes (Sharma et al., Citation2014; Vetaas & Grytnes, Citation2002) to monotonically decreasing (Ghafari et al., Citation2018; Kala & Mathur, Citation2002), depending on whether parts or whole gradient is taken into account (Rahbek, Citation1995). A variety of environmental factors change over short distances along an altitude gradient, such as temperature, precipitation, solar radiation, and wind speed which directly or indirectly influence plant diversity and its composition (Cui et al., Citation2019; Khuroo et al., Citation2010; Körner, Citation2007; Laiolo et al., Citation2018). Owing to this, plant occurrence and composition in mountain system are more influenced by environmental conditions (Körner, Citation2007; Rawal et al., Citation2018), thus making altitude a suitable gradient for studying spatial variation in diversity and vegetation composition as well as the effects of climate change.
Himalaya, the youngest mountain ecosystem, is globally recognized as biodiversity hotspot and inhabits a rich repository of unique biodiversity (Carpenter, Citation2005; Mallen-Cooper & Pickering, Citation2008). The vast mountain range (over 2500 km long and 80 to 300 km wide), rising from low-lying plains and reaching upto 8000 m asl, provides altitude range exhibiting wide bioclimatic regimes (Vetaas & Grytnes, Citation2002). The two broad climate regimes of Himalaya are monsoonal and non-monsoonal. While the great Himalayan belt occurs under the Indian monsoon regime characterized by glacial features like lateral and median moraines, the arid mountainous tracts in the extreme north and parallel to the Himalayan range represent the non-monsoonal regime. These high-altitude biomes exhibit extreme climatic conditions such as diurnal temperature fluctuations, scanty rainfall, and high wind velocity and are completely snowbound for months. The soil is poor in organic matter, nitrogen content, sandy or sandy-loam in texture and a pH range of 7–11 (Körner, Citation2007; Negi, Citation2002). Owing to these extreme conditions, the region exhibitsa wide variety of ecological traitsalong with a number of specialized habits such as cushion forming, diminutive and bushy (Bhattarai et al., Citation2010; Pandey et al., Citation2018). Alpine vegetation starts from around 3200 m with lower elevations (3200–3500 m) forming temperate alpine forests dominated by tree species such asAbies pindrow, Cedrus deodara and Pinus wallichiana. Towards higher elevations, tree species are unable to cope with environmental stress and grow shorter and often in lower densities and marks the treeline above which they are unable to grow. This zone mainly comprises alpine scrubs dominated by bushy habit with prostrate, woody branches and long deep roots like Caragana spp., Juniperus spp. and Lonicera spp., followed by alpine meadows in higher elevations dominated by herbaceous growth form including forbs, sedges, tussock, and non-tussock grasses.The mounting anthropogenic pressure and over exploitation of natural resources have made the Himalayan ecosystem one of the world’s most fragile and threatened ecosystems (Bisht et al., Citation2022; Chandra Sekar & Srivastava, Citation2009; Murti, Citation2001). Any change or disturbance in the biotic and abiotic components of this unique ecosystem is bound to cause imbalance in the ecosystems. Due to remoteness, inaccessibility and harsh climatic conditions these high-altitude areas have largely escaped the attention of ecologists, geographers and natural resource managers. It is only in the recent decades that investigations have been taken up in high-altitude regions in Western Himalaya detailing the plant diversity and composition (Acharya et al., Citation2011; Raina & Sharma, Citation2012; Rawat et al., Citation2010, Citation2010; Rawat, Citation2005).
Under the global climate change scenario, a comprehensive understanding of vegetation distribution and dynamics at the upper elevational limit of vascular plants across all life forms context for management planning and development of strategies for achieving is needed along with their responses to environmental parameters. Further, it provides necessary sustainable management of the ecosystem (Negi et al., Citation2018). Keeping in view the above considerations and the vast gap areas, the present study aims to assess diversity and distribution patterns of different life forms along altitude gradient in alpine regions of Uttarakhand, west Himalaya. This study has considered multiple representative altitude transects spread across entire alpines of Uttarakhand Himalayas for providing answers to following questions: (i) how does species richness, diversity and density patterns in various life forms vary along altitude gradient? (ii) Does vegetation compositiondiffer considerably with altitude as well across regions? (iii) How climatic factors influence plant species abundance in high-altitudes?
Material and methods
The study was conducted in high-altitude (>3000 m) alpine regions of Uttarakhand, west Himalaya. Positioned in the rain shadow of the Himalaya, the region remains unaffected by Indian monsoons, thus representing high-altitude non-monsoonal regime and exhibit extreme climate conditions, such as diurnal fluctuations in temperatures (−45ºC to 40ºC), scanty and erratic rainfall (<600 mm), high wind velocity and snowfall (Chandra Sekar et al., Citation2020). Furthermore, region consists of arid mountainous tracts constituted of sediments of Tethyan sea bed and sandy-loam with appreciable amount of clay and poor organic content. The vegetation found in the areas can be segregated into temperate forests with open canopy and stunted growth (scrubs and meadows) (Murti, Citation2001). After several field visits and consultation of relevant literatures (Aswal & Mehrotra, Citation1994; Murti, Citation2001; Negi, Citation2002; Vashistha et al., Citation2011), a study area map was extracted and six altitudes transects were selected for vegetation assessment (see S1 Table):
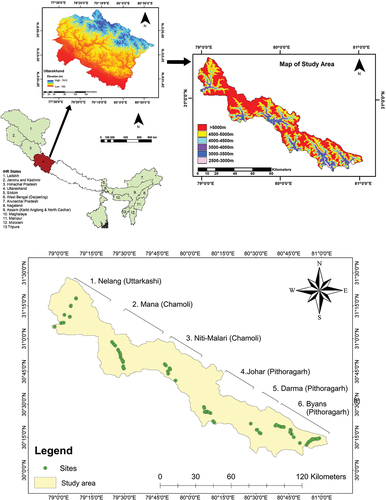
Figure 1. Study area (Source: http://srtm.csi.cgiar.org) and locations of seven studied altitude transects in alpine regions of Uttarakhand.
Sample design and vegetation assessment
Intensive survey trips were carried out during 2014 to 2018 in selected transects in order to prepare a baseline of plant species across all life forms (i.e., trees, shrubs and herbs). Collection of plant specimen was done in accordance with the permission of State Forest Department received from Chief Wildlife Warden (Letter no. 917/5-6/DDN/dated 16 September 2013) and complying with the IUCN Policy Statement Involving Species at Risk of Extinction. To support reproducibility, voucher specimens for all collected specimens were deposited in the herbarium of GBP National Institute of Himalayan Environment, Kosi, Almora (Acronym- GBP). Information regarding collected plants list, their voucher specimen (Accession no.) and collector has been provided in Supplementary File (S1 Table) along with the complete list of plants species. Specimens were collected as per the methods given in Jain and Rao (Citation1977) and identified using local and regional floras (Deva & Naithani, Citation1986; Gaur, Citation1999; Naithani, Citation1984; Osmaston, Citation1927; Pusalkar & Singh, Citation2012) and earlier herbarium records (i.e., Botanical Survey of India, Dehradun, Kolkata; Forest Research Institute – Dehradun).
Selected representative transects were investigated for compositional patterns of vegetation using stratified random sampling. Beginning from 3200 m, each transect was divided into 100 m altitude bands upto its respective upper elevational limits to carry out vegetation assessment along altitude gradient. Within each altitude band, three random sample plots of 0.25 ha (50 × 50 m2) were established. In each sample plot, random sub-plots were laid, i.e., 10 (10 × 10 m2) for trees, 5 (5 × 5 m2) for shrubs and saplings, and 10 (1 × 1 m2) for herbs and seedling. For tree species, individuals with <10 cm girth and <20 cm height were considered as seedlings, those between 10–30 cm girth and >20 cm in height as saplings and with >30 cm girth at breast height of 1.37 m as trees. Number of individuals of plant species per quadrat was taken, pooled and analysed as per standard phytosociological approaches (Gairola et al., Citation2014; Goswami & Das, Citation2018; Mishra, Citation1968; Mueller-Dombois & Ellenberg, Citation1974; Rawat & Everson, Citation2012; Zou et al., Citation2013) and Important Value Index was calculated as per (Floeter et al., Citation2004).
IVI = Relative Density + Relative Frequency + Relative Abundance
Species richness, S = Total number of species
Shannon diversity index, H; and Beta diversity (β) was calculated using Whittaker (Citation1960) formula as given in Mena and Vázquez‐Domínguez (Citation2005):
where α is the mean number of species per altitude belt, and s is the total number of species recorded across the study system (i.e., altitude transect). The Shannon provides the diversity of species in a community (i.e., altitudinal basis).
For each of the transect, values of selected environmental variables (average temperature, average precipitation, average solar radiation, average wind speed, average isothermality), which represent different direct and indirect gradients important for plant distribution (Williams et al., Citation2012), were obtained along selected altitude ranges from WorldClim version 2.1 climate data for 2000–2018 [https://www.worldclim.org/data/worldclim21.html] a spatial resolution of 30s (~1 km2).
Data processing
To numerically model distribution of different habit along altitude gradients, quadratic models were fitted between altitude and species distributions indices (richness, diversity, density). Selection of the quadratic model was made based on the model performance that was primarily evaluated by computing statistically significant r2 values (P < 0.05). Moreover, to assess distribution of sample means from observations and model simulations, analysis of variance (ANOVA) was carried out, particularly, estimated F-value of each model was compared with critical F-value. The result section highlights those models for which a significantly high r2 value was noted at P < 0.05. Subsequently, results from ANOVA for these models were elaborated.
Non-metric multidimensional scaling (NMDS) and permutational multivariate analysis of variance (PERMANOVA, 999 permutations) were performed using R-vegan package (version 2.5–3) function “metaMDS,” to assess the community composition along various altitude transects, ordination plots were produced. Abbreviated scientific names of plant taxa used is documented in S2 Table. In order to select an optimum dimension for NMDS analyses for both herbs and shrubs, stress-dimension analyses were carried out and dimension 2 was used for both NMDS analyses of shrubs and herbs using “Bray-Curtis” distance. urther, influence of selected climatic parameters (temperature, precipitation, solar radiation, wind speed, isothermality) on species richness, diversity and density was evaluated using Pearson’s correlation coefficient method to check significant relationships among selected parameters. Further, Canonical Correspondence Analysis (CCA) was carried out to assess relationships between environmental parameters and species composition in study transects. The community data matrices of plant species for all transects were used with the constrained matrices composing of environmental parameters, (i) altitude, (ii) temperature, (iii) precipitation, (iv) solar radiation, (v) isothermality, (vi) wind speed. The nature of relationships between soil parameters and vegetation distribution was depicted through ordination diagram. The CCA analyses were carried out using the R-Vegan package (version 2.5–3) function “CCA” and results were analysed with respect to inertia, ranks and eigen values of constrained matrices and parameters.
Results
Floristic diversity pool
A total of 265 vascular plants belonging to 155 genera and 55 families were observed along the investigated altitude range (i.e., 3200–4800 m) of Uttarakhand alpines (S2Table). Among these, nine were gymnosperms distributed in six genera and three families, while the rest belong to angiosperms. Asteraceae was the most dominant family having 30 plant taxa followed by Rosaceae (21) and Ranunculaceae (18). In terms of growth forms, herbaceous habit (81.7%) was dominant, followed by shrubs (14.9%) and trees (3.4%). In relation to 500 m altitude zones, plant population was maximumin the lowest zone, i.e., 3000–3500 m (72% of total flora), followed by 3501–4000 m (62%), 4001–4500 m (38%) and 4501–4800 m (9%) , thus exhibiting a continuous decline in the number of plant species with increasing altitude. There was a more rapid decline in distribution of higher level of taxa (genera and family) with altitude as compared to that of species . The species to genera (S/G) value calculated for the entire landscape was 1.1 for trees, 2.0 for shrubs and 1.7 for herbs, while, species to family (S/F) values were 1.8 for trees, 3.1 for shrubs and 5.0 for herbs. Furthermore, across the altitude zones, S/G and S/F values declined monotonically from lower to higher zones for trees and shrubs. However (S/F) ratio in 3501–4000 m zone was higher (3.9) as compared to that in 3000–3500 m (3.6) for herbs .
Table 1. Floristic diversity pool (S- Species, G- Genus, F- Family) in high-altitude alpine region of Uttarakhand along altitude gradient.
At quadrate level, species richness was maximum in Johar, for all growth forms, ranging, while Shannon diversity Index (H’) was highest in Niti-Malari for tress (1–1.05), Johar for shrubs (0.45–0.91) and Byans 1 for herbs (0.91–1.33) (S3 Figure). Broad range of compositional attributes of vegetation in study transects depicted higher densities of trees in Byans 1 &2, i.e., 20–240 ind/ha, while shrub and herb densities were higher in Darma (1200–3000ind/ha) and Byans 1 (31000–115000ind/ha), respectively (S4 Figure). Beta diversity values calculated for each life form along altitude gradient exhibited an increasing trend in all transects, thus indicating scattered and heterogeneous plant composition especially at high altitudes. Apart from this, beta diversity contributed significantly to transect level species richness of Byans, Johar, Mana and Nelang for herbs and shrubs, but very little for trees. However, in Darma tree species change rapidly while herbs and shrubs change slowly and in Niti-Malari, shrub species exhibited rapid change .
Table 2. Beta diversity variations across life forms and transects.
Altitudinal relationship of compositional features
Considering the alpine treeline area broadly varies from 3000–4000 m in the west Himalayas, treeline in present study was identified at 3200–3900 m with 1–3 tree species per plot, 3–11 shrubs per plot and 9–30 herbs per plot). Patterns of species richness and diversity of different growth forms across altitude rangewere modelled and a 2nd order polynomial regression showed a significant fit (p < 0.05, , with both richness and diversity monotonically decreasing with increasing altitude. While the decrease in tree diversity was steep at altitude zone 3600–3800 m with no species observed above 4000 m, it was moderate for herbs and shrubs upto the upper hard boundaries. Furthermore, density distribution of different growth formsalong altitude gradientfollowed a similar decreasing pattern. For different size classes (i.e., trees, saplings, seedlings), density decreased at higher altitudes drastically, ranging from 10–540 ind/h for trees, 10–80 ind/h for saplings and 100–1000 ind/h for seedlings ( (S2 Figure). As for shrub and herb densities in relation to altitude, a relatively moderate but significant decline (p < 0.05) was seen (. Based on the Important Value Index (IVI) of plant taxa in each altitude, dominant and codominant species across three growth forms are listed in . Cluster dendrogram for the study area identified four major altitudinal zones based on species composition, i.e., 3200 to 3500 m, 3600 to 3900 m, 4000 to 4500 m and 4600 to 4800 m (. Lowest altitude zone (3200–3500 m) inhabited tree species such as Pinus wallichiana, Abies pindrow, Prunus cornuta and Cedrus deodara, while Betula utilis, Juniperus semigloboosa and Salix denticulata were present in 3600–3900 m zone. Among shrubs, the species of Astragalus, Berberis, Cotoneaster and Salix were dominant in lower altitude zones, while Cassiope fastigiata, Ephedra gerardiana, E. intermedia, Juniperus indica and Rhododendron anthopogon are mostly dominated in high-altitudes (. Moreover, types of dominant herbaceous plants highly varied across altitude gradient. At lower altitude zones several common species such as Arenaria bryophylla, Bistorta affinis, Danthonia cachemyriana, Geranium wallichianum, Oxyria digyna, Oxytropis lapponica, Potentilla spp., Taraxacum officinale and Thymus linearis were present in fairly high proportions, whereas higher altitudes exhibited clear dominance of Bistorta vivipara, Carex nivalis, Gentiana stipitata, Geranium wallichianum, Juncus concinnus and Meconopsis aculeata.
Figure 2. Species richness, diversity and density trends along altitude gradients in different life forms [A] Trees, [B] Shrubs, [C] Herbs in alpine of Uttarakhand, west Himalaya.
![Figure 2. Species richness, diversity and density trends along altitude gradients in different life forms [A] Trees, [B] Shrubs, [C] Herbs in alpine of Uttarakhand, west Himalaya.](/cms/asset/2e1f8005-295a-4f30-88d7-ca70e0d7bc81/tgel_a_2163606_f0002_b.gif)
Figure 3. Hierarchical cluster dendrogram showing distinct altitudinal zones with respect to the species composition in alpine region of Uttarakhand, west Himalaya.
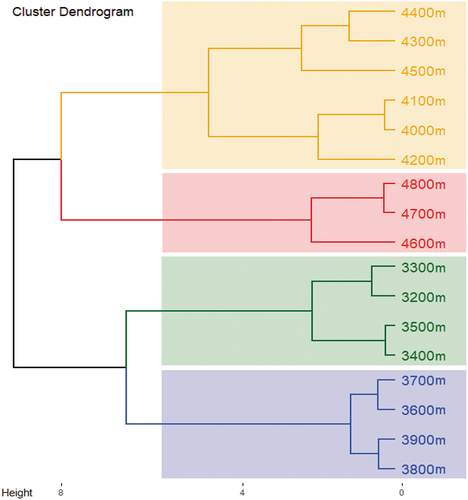
Table 3. Dominant and co-dominant taxa along altitude gradient in studied alpine regions of Uttarakhand, west Himalaya.
Vegetation composition across different transects
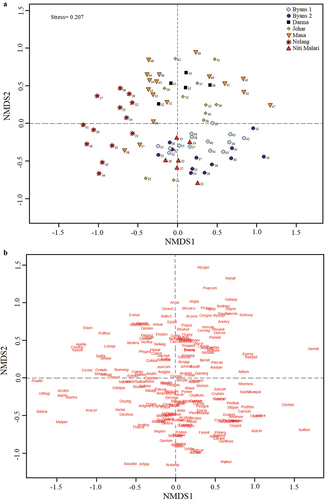
NMDS of vegetation abundance and combined dataset of studied transects further exhibited a strong gradient separating sample (plots) and species between transects as well as altitude range along the first two axis (NMDS1 and NMDS2) (. Separation along the first axis explained 40% of variation while second axis explained 20%, thus underscored the affinity between different altitudinal vegetation zones. Across all transects, distinct clusters denoting vegetations zones were identified characterized by specific species. Nelang transect formed a cluster towards the negative end of first axis, with plotsat higher altitude (3900–4400 m) in more proximity with each other inhabiting speciessuch as Aconogonum tortuosum, Melica persica, Poa tibetica, Sisymbrium brassiciforme, Urtica hyperborea. Lower and mid-altitude plots of Byans 1 (3200–3700 m) and Byans 2 (3200–3800 m) as well as NitiMalari transect plots formed a separate cluster towards negative end of second axis, inhabited by similar species (such as Carex nivalis, Delphinium brunonianum, Primula denticulata, Rheum webbianum, U. dioca). On contrary, higher altitude plots in Byans 1 (>3800 m) & Byans 2 (>3900 m) formed a distinct cluster towards positive end of first axis characterised by species such as Christolea himalayensis, Leontopodium brachyactis, Ranunculus palmatifidus, Saxifraga flagellaris. For Johar, study plots were scattered across the four quadrants, thus exhibited large variations in species composition along altitude gradient. Lower (3200–3500 m) and higher altitude plots (4400–4800 m) in Mana formed distinct cluster with Darma transect showing proximity in their species composition inhabiting Cotoneaster spp., Convolvulus arvensis, Polygonum polystachyum, Rosa macrophylla. Mid-altitude zones in Mana, i.e., 3600–3800 m, showed similarity with Nelang, while 3900–4300 m zone was scattered along positive end of first axisalong with species like Galium asperuloides Nardostachys jatamansi, Picrorhiza kurroa, Poa compressa and Saussurea simpsoniana. Overall, sample plots in lower altitudes of transects were more clustered towards the middle of both axes, inhabiting common plant species. Most of the sample plots in higher altitudes occupied extreme ends (positive and negative) of both axes, thus showing increase in rarity in species composition moving from low to high altitudes.
Figure 4. Non-metric Multi-Dimensional Scaling (NMDS) representing species composition among sample plots along altitude gradients of study transect.
Relationship of vegetation composition with environmental variables
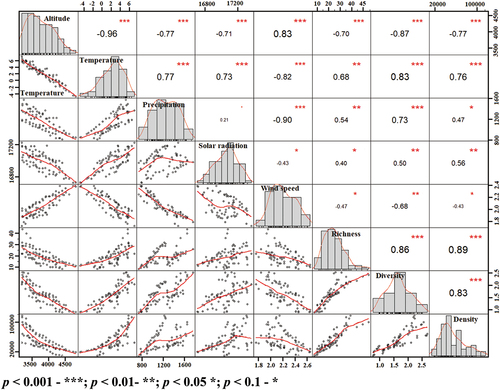
Correlation analysis among climate variables and plant diversity indices is depicted in along with Pearson’s correlation coefficient and significance level of relation. As established earlier also, altitude has a significant effect on density, diversity, and richness of plant species (p < 0.001), with an apparent decrease in density (r= -0.77), diversity (r = −0.87), and richness (r = −0.70). Similarly, there was strong negative correlation (p < 0.001) of temperature (r = −0.96), precipitation (r = −0.77), and solar radiation (r = −0.71) with altitude as well, while it was positively significant with wind speed (r = 0.83). Apart from altitude, temperature and precipitation also made a strong gradient influencing plant species richness and diversity in alpine regions . Increase in temperature as well as precipitation favoured comparatively more number as well as variety of species in study area, which is evident with significantly positive correlation coefficient values (p <0.01). As indicated by r values, solar radiation and wind speed exhibited significant (p < 0.05) but weak correlation with richness and diversity indices, thus exhibiting little influence on presence and types of plant taxa in alpine.
Figure 5. Pearson’s Correlation between plant richness, density and diversity indices with climate variables.
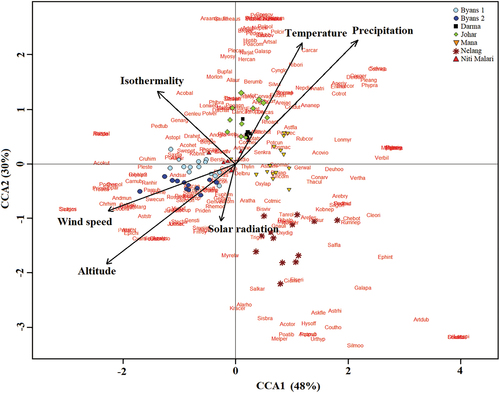
The relative effect of climatic parameters on plant species composition, analysed by CCA, showed strongest influence of precipitation in forming altitudinal zones having specific vegetation composition with high axes correlation (), followed by temperature, isothermality, and wind speed, while influence of solar radiation was lowest. Total inertia of combined data set was 13.02 where constrained climatic parameters explained 48% of total variances on first axis and 30% on second axis. Among altitude transects, in Byans 1 & 2 due to the high wind speed, low temperature and precipitation species like Christolea himalayensis, Gentiana stipitata, Leontopodium brachyatis, Rhodiola bupleuroides, Saussurea gossypiphora, S. simpsoniana and Sibbaldia parviflora. Contrary to this, species like Anaphalis nepalensis, A. triplinivalis, Berberis umbellata, Carum carvi, Cotoneaster rotundifolius and Ribes orientale favoured lower altitude transects like Darma and Johar with high temperature and precipitation. Occupying positive end of first axis and negative end of second axis, Nelang transect exhibited very little dependence on temperature, precipitation and wind speed. In context of plant species, the site thus occupies herbs like Aconogonum tortusum, Rumex nepalensis, Urtica hyperborea and Verbascum thapsus. Furthermore, isothermality of study site favoured growth and abundance of species like Aconitum balfourii, Gentiana argentea, G. leucomelaena and Pedicularis tubiformis.
Figure 6. Canonical Correspondence Analysis of combined data set: ordination of sample pots and vegetation abundance constrained by their relationships to environmental variables. Significant parameters are shown as biplot vector.
Discussion
The results of our study are based on data gathered during a time period from 2014 to 2018 of field work. A baseline floristic survey of study area (field and literature based) documented a total of 932 plant taxa belonging to 371 genera in 76 families Chandra Sekar et al., Citation2022. Hence, in comparison with previous studies (Negi, Citation2002), the high-altitude alpine zones (3200–4800 m) of Uttarakhand accounts for 58% alpine flora of west Himalayas. While comparing with pan-Himalayas (Rana & Rawat, Citation2017), the study area inhabits 10% species, 16% genera, and 32% families. Furthermore, 47 plants were under different threat categories as per IUCN and CAMP (Ved et al., Citation2003). Among these, Dactylorhiza hatagirea, Nardostachys jatamansi and Picrorhiza kurroa were “Critically Endangered,” while Aconitum heterophyllum, Angelica glauca, Ephedra gerardiana, Fritillaria roylei, Saussurea obvallata and Podophyllum hexandrum were “Endangered.” Hence, any effort made towards conservation ofthis high-altitude zone would eventually benefit a significant proportion of Himalayan biodiversity including flora as well as fauna. Dominance of herbaceous growth form in terms of richness, diversity as well as density is apparent due to extreme climatic conditions in the region which causes poor assemblages of trees and shrubs (generally high stature species). Apart from this, dominance of families such as Asteraceae and Rosaceae can be attributed to their specialized morphology, and broad ecological amplitude enabling them to establish their abundance in different communitieswith extreme conditions (Abbas et al., Citation2019).
The patterns of S/G along altitude gradient are used to describe speciation and diversification rates (Floeter et al., Citation2004). The general altitudinal decrease in S/G as well as S/F recorded in our study indicates their phylogenetic dispersion towards higher altitudes. On comparing the overall S/G values of different life forms, the higher ratio for shrubs indicates more intense diversification within the genera as compared to trees and herbs. Similarly, within family herbs exhibit significantly more intense phylogenetic clumping as compared to others. The increasing values of beta diversity calculated along altitude gradient for each transect indicates heterogenous plant composition, thus exhibiting more scope for occurrence of rare taxa as compared to lower altitudes (Rawal et al., Citation2018). This can be attributed to the variable levels of environmental as well as anthropogenic pressure prevailing in the different study areas. Relatively high level of disturbance and poor species pool leads to lower beta diversity in herbs, while it exhibits a reverse trend for trees. Among the studied transects, Darma with low β diversity exhibits high species packaging and homogenous composition in case of herbs, while becomes highly heterogeneous for trees (high β diversity). Similarly, with high beta diversity of herbs in Mana transect there is more scope for occurrence of rare/occasional herbaceous species, which contributes for higher heterogeneity of species composition.
Several studies have demonstrated characteristic relationships between altitude and patterns of species richness, diversity, and compositional features (such as density, total basal area) along a gradient (Acharya et al., Citation2011; Rawat et al., Citation2010; Sharma & Raina, Citation2013). In present study also, density and altitude exhibited highly significant relationship (p < 0.05, F>Fcrit) in entire study zone. This signifies the strong dependence of floristic diversity and density with altitude, irrespective of the growth stage or life forms in the alpine region. Our results are congruent with several former studies (Kala & Mathur, Citation2002; Pandey et al., Citation2018; Rawal et al., Citation2018; Sharma et al., Citation2014; Sinha et al., Citation2018; Vetaas & Grytnes, Citation2002; Wang et al., Citation2006). Reduction of plants in higher altitude may be attributed to eco-physiological constraints, such as extremely low temperature, precipitation and solar radiation ( which results in short growing season paving way for plant species tolerant to extreme conditions, a phenomenon known as “ecological filtering.” These fluctuations also influence soil texture, nutrients and stability of substrate which in turn are responsible for maintaining vegetation composition (Sanchez-Gonzalez & Lopez-Mata, Citation2005). Hence, forming distinct vegetation zones along altitude, with tree species occupying the lowest zone along with woody anddeep-rooted shrubs like Caragana spp., Juniperus spp. and Lonicera spp., followed by meadows at higher elevations dominated by herbaceous growth form including forbs, sedges, tussock and non-tussock grasses.
Ordination analysis (NMDS) clearly exhibited variation in vegetation composition among transects (. While temperature was the major factor responsible for variation along altitude, among transects precipitation had the strongest influence on vegetation composition, followed by temperature, wind speed and isothermality as depicted by CCA analysis. Byans and Niti-Malari regions with cold, dry climate and high wind speed had similar species composition. Nelang region formed a separate cluster due to its variable climate such as comparatively low temperature, precipitation, isothermality and wind speed. Darma and Johar fell in the same cluster with comparatively warm and wet climate experiencing slower winds and optimum isothermality.
While comparing to earlier literature (Chandra Sekar et al., Citation2015; Garbyal et al., Citation2007; Lohani et al., Citation2013; Mitra et al., Citation2017; Rawat, Citation2005; Uniyal et al., Citation2002), the density of Aconitum heterophyllum, Allium stracheyi, Bergenias tracheyi, Carum carvi, Hippophae salicifolia, Rhododendron anthopogon, R.campanulatum, Swertia ciliata and Thymus linearis was lesser, while that of Juniperus communis, J. indica, Polygonatum verticillatum density was increased. This can be attributed to the irregular harvesting of former species for fuel and medicinal purposes. The population of Betula utilis highly decreased in the area due to its utilization for fuelwood in higher Himalayan regions (Mitra et al., Citation2017). Climatic barriers, limiting tree habits, resulting limited density coupled with high extraction as fuelwood may be the reason of population reduction. Furthermore, trees and saplings of B. utilis are available, but there are no seedlings recorded in 3500 m. Thus, protection of these saplings is need of the hour, so that the population of B. utilis can be saved. In view of importance of the tree, the local considered it as “sacred” and various conservation activities were observed. The people of the local areas must aware about the importance of plants and some good initiatives (i.e., J. semiglobosa conservation in Johar) should be replicated in all other regions, to conserve the plant diversity. Furthermore, density and diversity are also influenced by frequency of anthropogenic disturbances, i.e., road construction, habitation, commercial plantation and agricultural performs also results in replacement of natural vegetation with man-made ecosystem.
Under current global climate change scenarios, a number of studies have projected changes in vegetation distribution and upward shifts in plant boundaries (Kuhn et al., Citation2016; Rai et al., Citation2010; Rosset et al., Citation2010; Telwala et al. Citation2013). These shifts tend to change the shape of ecosystems of high-altitude regions. However, shifting rate varies with species depending on their altitudinal and geographical distribution ranges, and their accumulative potential (Klimes et al., Citation2003). As species distribution range decrease with increasing altitude, species with narrow amplitude of altitudinal distribution and endemics are more affected by environmental or anthropogenic disturbances. While climate change (natural or anthropogenic) threatens high-altitude flora, it also impacts a significant portion of useful biodiversity and traditional livelihood of local people. Hence it becomes pivotal to carry out empirical studies pertaining to potential extent and magnitude of current and future shifts in altitudinal limits of plants in response to climate change in order to draw conclusive conservation and management strategies (Mehta et al., Citation2020; Pandey et al., Citation2019). Our study documents the plant species diversity and composition of remote areas of Himalaya which are known to be natural laboratories to study the responses of plants to climate change. While our study is more focussed on plant diversity, it can be expanded to broader aspects such as phenological and physiological studies of alpine plants and their relation with climatic and edaphic factors (Majeed et al., Citation2022) and opt more towards plant distribution models (Bayat et al., Citation2021; Jahani & Saffariha, Citation2020; Jahani et al., Citation2021; Mosaffaei et al., Citation2020).
Conclusion
This study is the first attempt to document and analyse floristic composition of high-altitude alpine regions panning across Uttarakhand, west Himalaya, along altitude gradient and determine major climate drivers influencing vegetation patterns. Hierarchical clustering as well as ordination analysis (NMDS and CCA) performed also showed concordant results, underscoring altitudinal vegetation zonation throughout study area as well as its relationship with environmental variables.Phytosociological studies undertaking multiple transects and life forms in high altitude alpine regions are very scarce especially in Indian Himalayan region. In this context, the present study also attempts to discuss and compare distribution patterns of vegetation across the longitudinal gradients along multiple altitudes transect. In this context, the present study gives comparable ecological data sets to discuss distribution pattern of seven altitudinal transects. Species richness and diversity along the altitude gradient in alpine region exhibited a monotonic decrease, both overall and by growth forms, with maximum values in the lower altitude zones (3200–3500 m). Beta diversity also increased with altitude thus exhibiting higher heterogeneity and presence of unique species adapted to survive the extreme environmental conditions. Among growth forms, beta diversity was generally higher for herbs and shrubs as compared to tress indicating homogeneity in tree populations at high-altitude alpines. The observed variations in temperature and precipitation patterns along altitude gradient significantly correlates with the species richness and diversity patterns, with high temperature and precipitation supporting plant richness and diversity. The linear correlation states altitude as the variable which best explains the variations in richness and diversity patterns.
Acknowledgements
The authors thankfully acknowledge the facilities provided by Director, G.B. Pant National Institute of Himalayan Environment (NIHE), Kosi-Katarmal, Almora for undertaking this work. We are gratefully acknowledging the financial assistance received from Council of Scientific and Industrial Research [EMR No. 38 (1346)/12/EMR-II], Kailash Sacred Landscape Conservation and Development Initiative and National Mission on Himalayan Studies (NMHS), Ministry of Environment, Forest and Climate Change, Government of India (GBPNI/NMHS-2018-19/SG 11) and National Mission on Sustaining the Himalayan Ecosystem (NMSHE), TF-3, DST, New Delhi. Permissions for Herbarium and library consultation received from different organizations, i.e., Botanical Survey of India, Forest Research Institute, Wildlife Institute of India, National Botanical Research Institute; Permission of State Forest Department received from Chief Wildlife Warden (Letter no. 917/5-6/DDN/dated 16 Sep. 2013); logistic and field guidance received from DG, ITBP (Letter no. 2766 dated 14.08.2014) are highly acknowledged. Dr. Poonam Tripathi, International Centre for Integrated Mountain Development (ICIMOD), Nepal is gratefully acknowledged for preparing study area map. The inputs received from anonymous reviewers aided in improving contents of this manuscript, we highly thank them.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Abbas, Z., Alam, J., Khan, S. M., Hussain, M., & Abbasi, A. M. (2019). Diversity, ecological feature and conservation of a high montane flora of the Shigar valley (Karakorum Range) Baltistan region, northern Pakistan. Pakistan Journal of Botany, 51(3), 985–1000. https://doi.org/10.30848/PJB2019-3(23)
- Acharya, B. K., Chettri, B., & Vijayan, L. (2011). Distribution pattern of trees along an elevation gradient of Eastern Himalaya, India. Acta Oecologica, 37(4), 329–336. https://doi.org/10.1016/j.actao.2011.03.005
- Aswal, B. S., & Mehrotra, B. N. (1994). Flora of Lahaul-Spiti (an alpine in north-west Himalaya). Bishen Singh Mahendra Pal Singh Dehradun.
- Bayat, M., Burkhart, H., Namiranian, M., Hamidi, S. K., Heidari, S., & Hassani, M. (2021). Assessing biotic and abiotic effects on biodiversity index using machine learning. Forests, 12(4), 461. https://doi.org/10.3390/f12040461
- Bhattarai, S., Chaudhary, R. P., Quave, C. L., & Taylor, R. S. (2010). The use of medicinal plants in the trans-himalayan arid zone of Mustang district, Nepal. Journal of Ethnobiology and Ethnomedicine, 6(1), 1–11. https://doi.org/10.1186/1746-4269-6-14
- Bisht, M., Chandra Sekar, K., Mukherjee, S., Thapliyal, N., Bahukhandi, A., Singh, D., Bhojak, P., Mehta, P., Upadhyay, S., & Dey, D. (2022). Influence of anthropogenic pressure on the plant species richness and diversity along the elevation gradients of Indian Himalayan high-altitude protected areas. Frontiers in Ecology and Evolution, 10, 751989. https://doi.org/10.3389/fevo.2022.751989
- Carpenter, C. (2005). The environmental control of plant species density on a Himalayan elevation gradient. Journal of Biogeography, 32(6), 999–1018. https://doi.org/10.1111/j.1365-2699.2005.01249.x
- Chandra Sekar, K., Lalit, G., Aseesh, P., & Srivastava, S. K. (2015). A note on distribution of juniperus semiglobosa in Uttarakhand, India. Indian Journal of Forestry, 38(1), 79–80. https://doi.org/10.54207/bsmps1000-2015-0XY6MS
- Chandra Sekar, K., Pandey, A., Giri, L., Joshi, B. C., Bhatt, D., Bhojak, P., Dey, D., Thapliyal, N., Bisht, K., Bisht, M., & Negi, V. S. (2022). Floristic diversity in Cold Desert regions of Uttarakhand Himalaya, India. Phytotaxa, 537(1), 1–62.
- Chandra Sekar, K., & Srivastava, S. K. (2009). Flora of the Pin Valley National Park, Himachal Pradesh. Botanical Survey of India. Ministry of Environment and Forests.
- Cui, Y., Bing, H., Fang, L., Wu, Y., Yu, J., Shen, G., Jiang, M., Wang, X., & Zhang, X. (2019). Diversity patterns of the rhizosphere and bulk soil microbial communities along an altitudinal gradient in an alpine ecosystem of the eastern Tibetan Plateau. Geoderma, 338, 118–127. https://doi.org/10.1016/j.geoderma.2018.11.047
- Deva, S., & Naithani, H. B. (1986). The Orchid Flora of North-West Himalaya. Print &Media Associates.
- Floeter, S. R., Ferreira, C. E. L., Dominici‐arosemena, A., & Zalmon, I. R. (2004). Latitudinal gradients in Atlantic reef fish communities: Trophic structure and spatial use patterns. Journal of Fish Biology, 64(6), 1680–1699. https://doi.org/10.1111/j.0022-1112.2004.00428.x
- Gairola, S., Rawal, R. S., Todaria, N. P., & Bhatt, A. (2014). Population structure and regeneration patterns of tree species in climate- sensitive subalpine forests of Indian western Himalaya. Journal of Forestry Research, 25, 343–349.
- Garbyal, S. S., Grover, A., Aggarwal, K. K., & Babu, C. R. (2007). Traditional phytomedicinal knowledge of Bhotias of Dharchula in Pithoragarh. Indian Journal Traditional Knowledge, 6(2), 360–364.
- Gaur, R. D. (1999). Flora of the District Garhwal, North–West Himalaya (with ethnobotanicalnotes). TransMedia.
- Ghafari, S., Ghorbani, A., Moameri, M., Mostafazadeh, R., & Bidarlord, M. (2018). Composition and structure of species along altitude gradient in Moghan-Sabalan rangelands, Iran. Journal of Mountain Science, 15(6), 1209–1228. https://doi.org/10.1007/s11629-017-4820-2
- Goswami, N., & Das, D. (2018). Status study of medicinal plants at Purnapani forest of Jhargram in West Bengal. IOSR Journal of Pharmacy and Biological Sciences, 13(4), 1–12.
- Jahani, A., Goshtasb, H., & Saffariha, M. (2020). Tourism impact assessment modeling of vegetation density for protected areas using data mining techniques. Land Degradation & Development, 31(12), 1502–1519.
- Jahani, A., & Saffariha, M. (2021). Human activities impact prediction in vegetation diversity of Lar National Park in Iran using artificial neural network model. Integrated Environmental Assessment and Management, 17(1), 42–52.
- Jain, S. K., & Rao, R. R. (1977). Field and Herbarium methods. Today and Tomorrow Publishers.
- Kala, C. P., & Mathur, V. B. (2002). Patterns of plant species distribution in the Trans‐Himalayan region of Ladakh, India. Journal of Vegetation Science, 13(6), 751–754. https://doi.org/10.1111/j.1654-1103.2002.tb02104.x
- Khuroo, A. A., Weber, E., Malik, A. H., Dar, G. H., & Reshi, Z. A. (2010). Taxonomic and biogeographic patterns in the native and alien woody flora of Kashmir Himalaya, India. Nordic Journal of Botany, 28(6), 685–696. https://doi.org/10.1111/j.1756-1051.2010.00750.x
- Klimes, L. (2003). Life-forms and clonality of vascular plants along an altitudinal gradient in E Ladakh (NW Himalayas). Basic and Applied Ecology, 4(4), 317–328. https://doi.org/10.1078/1439-1791-00163
- Körner, C. (2007). The use of ‘altitude’ in ecological research. Trends in Ecology & Evolution, 22(11), 569–574. https://doi.org/10.1016/j.tree.2007.09.006
- Kuhn, E., Lenoir, J., Piedallu, C., & Gégout, J. C. (2016). Early signs of range disjunction of submountainous plant species: An unexplored consequence of future and contemporary climate changes. Global Change Biology, 22(6), 2094–2105. https://doi.org/10.1111/gcb.13243
- Laiolo, P., Pato, J., Obeso, J. R., & Coulson, T. (2018). Ecological and evolutionary drivers of the elevational gradient of diversity. Ecology letters, 21(7), 1022–1032. https://doi.org/10.1111/ele.12967
- Lohani, N., Tewari, L. M., Kumar, R., Joshi, G. C., Ch, J., Kumar, S., & Upreti, B. M. (2013). Population studies, habitat assessment and threat categorization of Polygonatumverticillatum (L.) Allioni in Kumaun Himalaya. Journal of Ecology and the Natural Environment, 5(5), 74–82. https://doi.org/10.5897/JENE12.042
- Majeed, M., Lu, L., Haq, S. M., Waheed, M., Sahito, H. A., Fatima, S., Aziz, R., Bussmann, R. W., Tariq, A., Ullah, I., & Aslam, M. (2022). Spatiotemporal distribution patterns of climbers along an abiotic gradient in jhelum district, Punjab, Pakistan. Forests, 13(8), 1244. https://doi.org/10.3390/f13081244
- Mallen-Cooper, J., & Pickering, C. M. (2008). Linear declines in exotic and native plant species richness along an increasing altitudinal gradient in the Snowy Mountains, Australia. Austral Ecology, 33(5), 684–690. https://doi.org/10.1111/j.1442-9993.2008.01835.x
- Mehta, P., Chandra Sekar, K., Bhatt, D., Tewari, A., Bisht, K., Upadhyay, S., Negi, V. S., & Soragi, B. (2020). Conservation and prioritization of threatened plants in Indian Himalayan Region. Biodiversity and Conservation, 29(6), 1723–1745. https://doi.org/10.1007/s10531-020-01959-x
- Mena, J. L., & Vázquez‐domínguez, E. (2005). Species turnover on elevational gradients in small rodents. Global Ecology and Biogeography, 14(6), 539–547. https://doi.org/10.1111/j.1466-822X.2005.00189.x
- Mishra, R. (1968). Ecology work book. Oxford and IBH Publishing Co.
- Mitra, M., Kumar, A., Adhikari, B. S., & Rawat, G. S. (2017). Fuelwood resources and their use pattern by Bhotia community in Niti valley, Western Himalaya. Botanica Orientalis: Journal of Plant Science, 11, 1–6. https://doi.org/10.3126/botor.v11i0.21025
- Mosaffaei, Z., Jahani, A., Chahouki, M. A. Z., Goshtasb, H., Etemad, V., & Saffariha, M. (2020). Soil texture and plant degradation predictive model (STPDPM) in national parks using artificial neural network (ANN). Modeling Earth Systems and Environment, 6(2), 715–729.
- Mueller-Dombois, D., & Ellenberg, H. (1974). Aims and methods of vegetation ecology. Wiley.
- Murti, S. K. (2001). Flora of alpines of Western Himalaya 1 (Monocotyledons). Botanical Survey of India.
- Naithani, B. D. (1984). Flora of Chamoli (Vol. 1, p. 2). Botanical Survey of India, Howrah.
- Negi, S. S. (2002). Alpines of India. Indus Publishing.
- Negi, V. S., Giri, L., & Chandra Sekar, K. (2018). Floristic diversity, community composition and structure in Nanda Devi National Park after prohibition of human activities, Western Himalaya, India. Current Science, 115(6), 1056–1064. https://doi.org/10.18520/cs/v115/i6/1056-1064
- Osmaston, A. E. (1927). A Forest Flora for Kumaon. Government press, UnitedProvinces.
- Pandey, A., Chandra Sekar, K., Joshi, B., & Rawal, R. S. (2019). Threat assessment of high-value medicinal plants of cold desert areas in Johar valley, Kailash Sacred Landscape, India. Plant Biosystems-An International Journal Dealing with All Aspects of Plant Biology, 153(1), 39–47. https://doi.org/10.1080/11263504.2018.1448010
- Pandey, A., Rai, S., & Kumar, D. (2018). Changes in vegetation attributes along an elevation gradient towards timberline in Khangchendzonga National Park, Sikkim. Tropical Ecology, 59(2), 259–271.
- Pusalkar, P. K., & Singh, D. K. (2012). Flora of Gangotri National Park, Western Himalaya, India. Botanical Survey of India.
- Rahbek, C. (1995). The elevational gradient of species richness: A uniform pattern? Ecography, 18(2), 200–205. https://doi.org/10.1111/j.1600-0587.1995.tb00341.x
- Rai, H., Nag, P., Upreti, D. K., & Gupta, R. K. (2010). Climate warming studies in alpine habitats of Indian Himalaya, using lichen based passive temperature-enhancing system. Nature and Science, 8(12), 104–106.
- Raina, A. K., & Sharma, N. (2012). Species composition and diversity of tree species along an altitudinal gradient in Sewa catchment of north-western Himalayas, Jammu and Kashmir, India. Environment Conservation Journal, 13(1–2), 173–179.
- Rana, S. K., & Rawat, G. S. (2017). Database of Himalayan plants based on published floras during a century. Data, 2(4), 36–44. https://doi.org/10.3390/data2040036
- Rawal, R. S., Rawal, R., Rawat, B., Negi, V. S., & Pathak, R. (2018). Plant species diversity and rarity patterns along altitude range covering treeline ecotone in Uttarakhand: Conservation implications. Tropical Ecology, 59(2), 225–239.
- Rawat, G. S. (2005). Alpine meadows of Uttaranchal. Bishen Singh Mahendra Pal Singh.
- Rawat, S. Y., & Everson, C. S. (2012). Ecological status and uses of Juniper species in the cold desert environment of the Lahaul Valley, North-western Himalaya, India. Journal of Mountain Science, 9(5), 676–686.
- Rawat, Y. S., Vishvakarma, S. C. R., Oinam, S. S., & Kuniyal, J. C. (2010). Diversity, distribution and vegetation assessment in the Jahlmanal watershed in cold desert of the Lahaul valley, north-western Himalaya, India. iForest-Biogeosciences and Forestry, 3(3), 65–71. https://doi.org/10.3832/ifor0532-003
- Rosset, V., Lehmann, A., & Oertli, B. (2010). Warmer and richer? Predicting the impact of climate warming on species richness in small temperate waterbodies. Global Change Biology, 16(8), 2376–2387. https://doi.org/10.1111/j.1365-2486.2010.02206.x
- Sánchez-gonzález, A., & López-Mata, L. (2005). Plant species richness and diversity along an altitudinal gradient in the Sierra Nevada, Mexico. Diversity & Distributions, 11(6), 567–575. https://doi.org/10.1111/j.1366-9516.2005.00186.x
- Sharma, N., & Raina, A. K. (2013). Composition, structure and diversity of tree species along an elevational gradient in Jammu province of north-western Himalayas, Jammu and Kashmir, India. Journal of Biodiversity and Environmental Sciences, 3(10), 12–23.
- Sharma, P., Rana, J. C., Devi, U., Randhawa, S. S., & Kumar, R. (2014). Floristic diversity and distribution pattern of plant communities along altitudinal gradient in Sangla Valley, Northwest Himalaya. The Scientific World Journal, 20, 1–11. https://doi.org/10.1155/2014/264878
- Sinha, S., Badola, H. K., Chhetri, B., Gaira, K. S., Lepcha, J., & Dhyani, P. P. (2018). Effect of altitude and climate in shaping the forest compositions of Singalila National Park in Khangchendzonga Landscape, Eastern Himalaya, India. Journal of Asia-Pacific Biodiversity, 11(2), 267–275. https://doi.org/10.1016/j.japb.2018.01.012
- Telwala, Y., Brook, B. W., Manish, K., & Pandit, M. K. (2013). Climate-induced elevational range shifts and increase in plant species richness in a Himalayan biodiversity epicentre. Plos One, 8(2), e57103. https://doi.org/10.1371/journal.pone.0057103
- Uniyal, S. K., Awasthi, A., & Rawat, G. S. (2002). Current status and distribution of commercially exploited medicinal and aromatic plants in upper Gori valley, Kumaon Himalaya, Uttaranchal. Current Science, 82(10), 1246–1252.
- Vashistha, R. K., Rawat, N., Chaturvedi, A. K., Nautiyal, B. P., Prasad, P., & Nautiyal, M. C. (2011). Characteristics of life-form and growth-form of plant species in an alpine ecosystem of North-West Himalaya. Journal of Forestry Research, 22(4), 501–506. https://doi.org/10.1007/s11676-011-0194-4
- Ved, D. K., Kinhal, G. A., Ravikumar, K., Prabhakaran, V., Ghate, U., Vijayashankar, R., & Indresha, J. H. (2003). Conservation assessment and management prioritisation. FRLHT.
- Vetaas, O. R., & Grytnes, J. A. (2002). Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal. Global Ecology and Biogeography, 11(4), 291–301. https://doi.org/10.1046/j.1466-822X.2002.00297.x
- Wang, W., Wang, Q., Li, S., & Wang, G. (2006). Distribution and species diversity of plant communities along transect on the northeastern Tibetan Plateau. Biodiversity & Conservation, 15(5), 1811–1828. https://doi.org/10.1007/s10531-004-6681-6
- Whittaker, R. H. (1960). Vegetation of the Siskiyou mountains, Oregon and California. Ecological monographs, 30(3), 279–338. https://doi.org/10.2307/1943563
- Williams, K. J., Belbin, L., Austin, M. P., Stein, J. L., & Ferrier, S. (2012). Which environmental variables should I use in my biodiversity model? International Journal of Geographical Information Science, 26(11), 2009–2047. https://doi.org/10.1080/13658816.2012.698015
- Zou, Y., Sang, W., Bai, F., & Axmacher, J. C. (2013). Relationships between plant diversity and the abundance and α-diversity of predatory ground beetles (Coleoptera: Carabidae) in a mature Asian temperate forest ecosystem. Plos One, 8(12), e82792.