
Abstract
Over the last few decades, due to its relevant function in male reproduction assessment, important molecular achievements have been made in the molecular characterization of estrogen receptor genes in various species. Our work focuses on a male seasonal breeder, the bioindicator Podarcis sicula, because of its peculiar gonadal anatomy, similar to that of humans. Based on the cloned lizard’s gene sequence fragment of estrogen receptor beta, esr2 (GenBank JN705543.1), we found DNA binding domain identity of 99% as well as a homologous sequence with humans. Furthermore, in order to better illustrate how this gene is regulated in the lizard’s reproductive system organs, we investigated the transcriptional activity of esr2 in brain and testis tissues during mating and winter stasis phases of the reproductive cycle. Quantitative real time-polymerase chain reaction (qRT-PCR) analyses performed on male gonadal tissues demonstrate a significant increase in esr2 expression during mating compared to the winter stasis period, while in the brain, esr2 shows the opposite trend. Next, we provide morphological evidence of the detrimental effect on spermatogenesis of a pure anti-estrogen treatment (ICI 182,780) and the corresponding effect on esr2 expression in lizard specimens during the mating period which, upon treatment, was found to be no different from the expression levels in winter stasis both in the brain and in the testis. In this study, we explore the potential use of Podarcis sicula as a model for human testis development and maturation, as well as esr2 expression for toxicological screening in one-testis gonadectomy.
Introduction
Estrogen receptors, like all members of the nuclear receptor superfamily, share a common domain organization consisting of a region located in the central part of the gene that binds DNA (DNA binding domain, DBD) which is highly conserved; a domain involved in binding with the ligand, in the dimerization and transactivation (ligand binding domain, LBD); variable N-terminal domain and C-terminal regions; and a hinge region between the DBD and LBD showing phylogenetic steroid receptor conservation (Gronemeyer & Laudet Citation1995; Mahfouz et al. Citation2016). Estrogen receptor transcriptional activity is induced by estrogens in response to changing demands due to factors such as female and male reproductive status, cognitive function, mood and environmental pollution (Zhang et al. Citation2009; Tohyama et al. Citation2016; Cooke et al. Citation2017). In previous work in this field, estrogen receptor alpha (ER-ALFA, esr1) and estrogen receptor beta (ER-BETA, esr2) were immunodetected, and their cDNA was cloned and quantified in many vertebrates (Katsu et al. Citation2004; Verderame & Limatola Citation2010; Zhang et al. Citation2017). Further, in teleost fish a third estrogen receptor class was identified and named gamma (ER-GAMMA) (Hawkins et al. Citation2000), while in some sharks the presence of only one estrogen receptor form was found (Katsu et al. Citation2010). All cloned receptors responded similarly to estrogen level variations whether with down- or up-regulation of expression in relation to tissue, gender, and the season for each species. However, they responded differently to estrogenic endocrine disruptors (Greytak & Callard Citation2007; Tohyama et al. Citation2016). Signal transduction through these nuclear receptors leads to dramatic changes in gene expression programs in different vertebrate cell types, typically due to their binding to DNA or to transcription modulators (Guerriero & Ciarcia Citation2001; Guerriero Citation2009; Guerriero et al. Citation2018b). esr1 and esr2, in particular, exhibit different biological roles and functions, as demonstrated by knock-out experiments in mice (Blatter & Mahoney Citation2015). The proposed estrogen receptor functions of esr2, in particular, include antiproliferative action, regulation of apoptosis, control of antioxidant gene expression, modulation of immune responses and DNA repair (Nikolos et al. Citation2018). Furthermore, esr2 is already known to be a key element in the signal transduction pathway for endocrine disruptor compounds (Tohyama et al. Citation2016). Xenobiotics, as well as substances able to compete with estrogens in the binding with esr2, even if they were usually unable to activate it, are known as anti-estrogens (Greytak & Callard Citation2007; Verderame et al. Citation2014; Tohyama et al. Citation2016; Walker & Gore Citation2017). Some of the most used anti-estrogens in cancer therapy are tamoxifen (Nolvadex) and raloxifene (Evista), which act as agonists or antagonists, depending on the target cells (Zhang et al. Citation2007; Arevalo et al. Citation2011; Jensen et al. Citation2018). Another known anti-estrogen is the ICI 182,780 (trade names Faslodex, Fulvestrant), which only exhibits anti-agonistic action toward estrogens and is thus considered a pure anti-estrogen (Alfinito et al. Citation2008; Guerriero et al. Citation2012; Gao et al. Citation2016). It is able to cross the blood-brain barrier, penetrate into brain and hypothalamic tissues, and affect known neuroendocrine functions. The esr2 gene has been studied in different tissues in reptiles including brain and gonads (Katsu et al. Citation2004, Citation2010; Verderame et al. Citation2014; Mahfouz et al. Citation2016), but its neuronal expression has never been investigated in relation to reproductive events although its importance in the regulation of reproduction is unquestionable. The esr2 gene coding for ER-BETA has been cloned and characterized in many fishes, birds, amphibians and reptiles (Katsu et al. Citation2004, Citation2010; Katsutoshi et al. Citation2007; Verderame & Limatola Citation2010; Zhang et al. Citation2014, Citation2017). The study of its expression in the brain may allow the monitoring of species. Specifically, the expression of esr2 can be used as a biomarker and its assessment may be considered a useful component of strategies for the conservation of species at risk of extinction (Guerriero et al. Citation2018b). This is because gametogenesis is under the control of the hypothalamic-pituitary system, which is substantially similar among the various vertebrate classes (for a review see Guerriero Citation2009), and their hormones. Sex steroid hormones, along with the pituitary gonadotropins, are able to modulate reproductive events, thanks to the presence of specific nuclear receptors and microRNAs (Zhang et al. Citation2009; Cao et al. Citation2018). The importance of the linkage between these structures is evident especially in seasonal breeding species, where environmental stimuli are integrated at the level of the central nervous system with the internal hormonal system and with endogenous antioxidants to trigger reproductive behavior (Guerriero et al. Citation2003, Citation2004, Citation2012, Citation2018b; Guerriero Citation2007). The use of seasonal breeding animals allows analysis of the physiological changes of the gonad and the monitoring of possible molecular “pathways” of the brain-gonad axis that regulate the progression of events for spermatozoon and oocyte formation (Guerriero Citation2009; Guerriero et al. Citation2018b). Previous research in our laboratories has shown that in the lizard Podarcis siculus (Podarcis sicula) testis, the estrogen receptor protein is at the highest levels during winter stasis, whereas the lowest gonadal levels occur during maximum activity or mating (Ciarcia Citation1993). The data suggest a seasonal modulation by the brain along the hypothalamic-hypophysis-gonad axis. For instance, results of testis estrogen measurements demonstrated more elevated levels during the mating phase. These results support the fact that the highest amount of local binding of estrogen occurs in winter stasis whereas estrogen plasma levels are higher in the mating phase (Andò et al. Citation1992). Evidence has been found in Podarcis testis demonstrating the direct influence of estrogens on the function and maturation of germ cells (Chieffi & Varriale Citation2004). Recently, our laboratories reported the Podarcis sicula esr2 sequence (GenBank JN705543.1). In the current study we report the characterization and expression of the esr2 dynamic in the brain and gonad of the male lizard. We describe the bioindicator Podarcis sicula followed through a decade of study, and the possibility of employing it, from a comparative toxicological perspective, as phylogenetically close to mammals, having similar anatomical germinal cell compartments. Finally, to expand our understanding of the role of Podarcis sicula as a bio-sentinel we address ICI 182,780 treatment to identify in the brain, as well as in the testis, its toxicological effect on germinal cells. These predictable answers will help us both to understand the brain-gonad feedback in normal status, with a possible role of Podarcis sicula testis gonadectomy, and to identify variability within the established brain-gonad dynamics when there is interference by anti-estrogen compounds.
Materials and methods
Wall lizard maintenance and experimental design
Sexually mature male wall lizards (total number 200 with n = 10 per each seasonal collection examined/10 years, weight range 10–14 g) identified as Podarcis sicula were collected (wild-caught) in the reference site of Monti di Lauro, Avellino (I), during two periods of the year: mating period, when there is full spermatogenetic activity (May), and winter stasis when the spermatogenesis is quiescent (November), in the period 2008–2017. The lizards were captured and the experiments were carried out in accordance with the ethical provisions enforced by the National Committee of the Italian Ministry of Health on in vivo experimentation (Department for Veterinary Public Health, Nutrition and Food Safety, SCN/2D/2000/9213). For 10 days before the experiments, the captured animals were housed in large terraria containing rocks, sticks and water. The animals were fed with mealworms Tenebrio molitor and fresh fruit ad libitum. All efforts were made to minimize animal suffering and to reduce the number of specimens used. Since they are heterotherms, they were euthanized at 4°C. For each specimen, brain and gonad were aseptically sampled. Some testes were dissected out and weighed, and only one plunged in Steve’s solution for histological studies with (n = 2) and without treatment with ICI 182,780 (n = 2), whereas the others, together with other tissues and organs, were quickly frozen at −80°C for molecular analysis.
Treatment with the estrogen receptor down-regulator ICI 182,780
The treatment took place during the mating period (May). Animals (n = 5 of mating collection/10 years) were subjected to a thermo-photoperiodic regime typical of the winter stasis period with a 16:8 h dark:light cycle and exposed to a controlled temperature of 10°± 1.5°C. Specimens were injected every 24 h with a subcutaneous dose of anti-estrogen ICI 182,780 (Tocris, catalog number 1047; M.W. 606.77; 6 mg/0.25 mL almond oil/animal). In parallel, control animals (n = 2) were injected with almond oil only (0.25 mL/animal/day). Collection of tissues and organs from treated animals occurred on the 21st day after the beginning of treatment, with three animals in each of the years from 2008–2017.
Histological analysis of the testis
Only one testis of three euthanized lizards for each condition (mating with and without ICI 182,780, and stasis) was fixed in Steve’s solution and subsequently embedded in paraffin-celloidin according to Peterfi (Mazzi Citation1977). Morphological staining was carried out on histological sections (7 μm thick) using Galgano’s trichome, and histological analysis was done using a Nikon-MicroPhot-FXA light microscope.
RNA extraction and cDNA synthesis
Total RNA was extracted by homogenization tissue in Trizol (Invitrogen), according to the manufacturer’s instructions. Briefly, testes and brain (50 mg) from euthanized Podarcis sicula (n = 5) of each phase of the reproductive cycle – mating and winter stasis – and in mating treated with ICI 182,780, during the years 2008–2017, were homogenized in 1 mL of Trizol reagent buffer, and then the homogenized samples were kept at room temperature for 15 min. A volume of 0.2 mL of chloroform per 1 mL of Trizol reagent was added. The samples were vortexed vigorously for 15 s, then incubated for 3 min at room temperature and centrifuged for 15 min at 4°C at 12,000 × g. After centrifugation, the upper aqueous layer was transferred to a fresh tube, then RNA was precipitated after mixing with isopropyl alcohol. Isopropyl alcohol (0.5 mL) was added to 1 mL of Trizol reagent used in the initial homogenization. Afterward, samples were incubated for 10 min at 30°C and centrifuged at 4°C for 10 min at 12,000 × g. The RNA pellet was detected after removing supernatant, then washed with 1 mL of 75% ethanol, centrifuged at 7500 × g for 5 min at 4°C. The RNA pellet was air-dried for 10 min, then resuspended in 100 µL diethylpyrocarbonate (DEPC)-treated water and stored at −80°C. The quality and the amount of purified RNA were assessed using spectrophotometer measurements at 260 and 280 nm (only samples with a ratio 260/280 ≥ 1.8 were accepted and further processed) and by electrophoresis under denaturing conditions on 1% agarose gel according to Raven et al. (Citation1979). To avoid the amplification of contaminant genomic DNA, we purified the total RNA from genomic DNA with an Ambion DNA-free kit. Total cDNA was synthesized from total RNA in duplicates for each sample to minimize reaction variations. One hundred nanograms of RNA in 20 μL reaction volume was used for each cDNA synthesis reaction using MMLV reverse transcriptase (Promega ImpProm II kit) according to the manufacturer’s instructions.
DNA sequence alignment and phylogenetic tree
The partial estrogen receptor beta (ER-BETA) protein sequence from Podarcis sicula (AFD18855.1) was aligned to all of the reference proteins available in the National Center for Biotechnology Information (NCBI) database by using protein-protein BLAST (BLASTp). Identified sequences were aligned and used to build a phylogenetic tree by using COBALT.
Quantitative real time-polymerase chain reaction (qRT-PCR)
Quantification of esr2 mRNA by RT-PCR was performed on five Podarcis sicula of each examined condition and the analyses were performed on two different tissues: testis and brain. 12S rRNA was used as a reference gene. qRT-PCR was carried out in a OneStep Plus Real-Time PCR System (Applied Biosystems), using SYBR Green (Applied Biosystems). Each reaction was performed in a 20-μL reaction volume containing 1 μL of a 1:5 dilution of cDNA preparation, 10 μL of 2x SYBR_ Green qPCR Kit (Applied Biosystems), and 5 μL 0.8 μM of each primer (PRIMM Biotech Products and Services, Milan, Italy). The specificity of the qRT-PCR was verified by sequencing the reaction product. Negative controls were performed on samples without cDNA in the reaction mix. Melting curves from each reaction were analyzed and all showed only one peak at the same Tm, indicating the absence of primer dimers or side products. The primers were designed on the esr2 sequence (JN705543.1) using the Primer3 software. The forward primer was 5ʹ-AAGAGAGCGCTGTGGCTATC-3ʹ and the reverse primer was 5ʹ-CAGTGCATTGACCAGCAGTT-3ʹ, giving a product size of 150 base pairs (bp). The PCR for the genes included an initial denaturation step at 95°C for 5 min, followed by 45 amplification cycles consisting of denaturation at 95°C for 30 s, annealing at 60°C for 30 s and an extension at 72°C for 60 s, plus a final extension at 72°C for 1 min. To ensure that a single product was amplified, melt curve analysis was performed on the PCR products at the end of each run. The PCR products were further assayed on a 2% agarose gel. The relative expression value was calculated as a fold change with the formula 2- ∆∆Ct (Livak & Schmittgen Citation2001).
Statistical analysis
Multiple group data were analyzed using one-way analysis of variance (ANOVA). The Student’s t-test was used to compare means between the groups. Statistically, significant differences are defined outside of the 95% confidence interval. Data are shown as a mean ± standard error of the mean (SEM). Statistical parameters are reported in the respective figures and figure legends.
Results
DNA sequence alignments
shows the sequence alignment of Podarcis sicula protein with the four best protein matches in the BLAST database of other species that belong to different families of the order Squamata, class Reptilia – Phyton bivittatus (Pythonidae), Plestiodon finitimus (Scincidae), Anolis carolinensis (Polychrotidae) and Elaphe quadrivirgata (Colubridae) – together with the human sequence showing phylogenetic relationships based on multiple sequence alignment and conserved protein domains. displays identities (%) with Podarcis sicula partial sequence (SmartBLAST) as well as the conserved domains of esr2. In particular, Podarcis sicula DBD shows 99% sequence identity with the human sequence. The LBD is also highly conserved between Podarcis sicula and Homo sapiens, showing 83% sequence identity.
Figure 1. (a) Phylogenetic tree of Podarcis sicula ER-BETA partial protein sequence and the four best sequence matches together with the human sequence, showing phylogenetic relationships based on multiple sequence alignment and conserved protein domains. (b) SmartBLAST identities (%) with Podarcis sicula partial sequence are indicated. The values 99% and 83% are the sequence identities of human DBD and LBD domains, respectively, vs Podarcis sicula. (c) Multiple sequence alignment of the sequences aligned in (a). The red color indicates identical residues (Cobalt). Conserved domains are indicated: DBD in blue and LBD in green.

shows the detailed multiple sequence alignment of the selected proteins, and the DBD and LBD are indicated.
Expression levels of esr2 during the reproductive stages
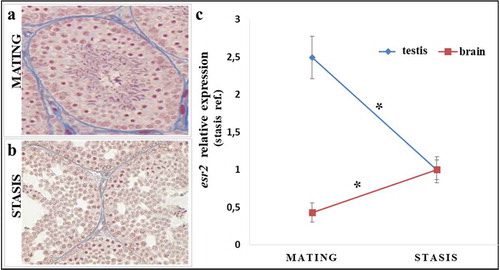
depicts the typical testicular organization during the mating period. The seminiferous tubules are well developed and all the germinal cells in different stages of maturation are clearly evident, including many sperm. shows representative sections of testis during the winter stasis phase; in the testis, the spermatogenetic processes appear substantially slowed down, with a reduced germinal epithelium, large lumen and gaps between the germ cells. Many degenerations are found in the seminiferous tubules, especially at spermatids; only a few very rare sperm are present, which will not be used for fertilization. After confirming the reproductive phase, we investigated the expression of the esr2 gene by qRT-PCR both in the testis and in the brain (). We included brains in our analysis to assess the possible impact of esr2 on the crosstalk brain-gonadal axis. Finally, we merged the data obtained in the various years considered to improve statistical power in the analysis of the combined pool of data. The qRT-PCR analysis in the gonadal tissues showed a significant increase of estrogen receptor beta gene expression during the mating phase compared to the winter stasis phase. Differently from gonads, the expression of esr2 in the brain is significantly higher in the winter stasis phase compared to the reproductive phase.
Figure 2. Histological variation in the stages of spermatogenesis (left) and expression of the esr2 gene by qRT-PCR both in the testis and in the brain (right) in Podarcis sicula: (a) mating period: in testis tubules all spermatogenetic stages, including many sperm, are present; (b) winter stasis period: all spermatogenetic stages are present but quiescient, with only a few sperm; (c) qRT-PCR expression analysis of esr2 gene of Podarcis sicula in testis and brain samples, in the mating period (May) and winter stasis period (November) at the reference site. The relative expression of esr2 is indicated as -fold change of the mean expression relative to the winter stasis period for testis and brain samples ± SEM (*p < 0.05). Reference gene, 12S rRNA gene.
Treatment with an estrogen receptor down-regulator (ICI 182,780)
In order to better understand the role of esr2 in the regulation of gonadal activity, we treated animals with the anti-estrogen ICI 182,780. highlights the histological organization of gonadal tissue upon treatment with the anti-estrogen. Treatment of animals with estrogen receptor down-regulator during the mating period of the reproductive cycle induces histological changes. In the testis, some seminiferous tubules show a slowdown of spermatogenesis processes, with a reduced lumen and few sperm. Severe testicular changes occurred, including an arrest of spermatogenesis at the secondary spermatocyte level. Treatment with ICI 182,780 induces a reorganization of the seminiferous tubules comparable to the winter stasis period.
Figure 3. Histological organization of gonadal tissue and expression levels of esr2 upon treatment with the anti-estrogen ICI 182,780 during the mating period (a) Seminiferous tubules show reduced lumen and few sperm; (b,c) qRT-PCR expression analysis of esr2 gene of Podarcis sicula brain (b) and testis (c) specimens, in the mating phase (May) and upon treatment in the mating period with the anti-estrogen ICI 182,780. The relative expression of esr2 is indicated as -fold change of the mean expression relative to the stasis period ± SEM (*p < 0.05). Reference gene, 12S rRNA gene.
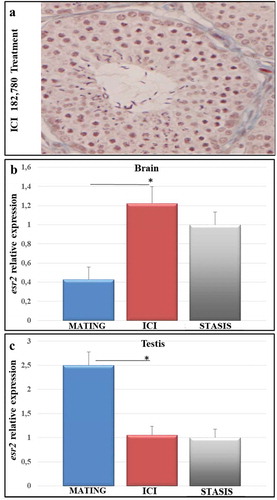
We also analyzed the expression levels of esr2 upon treatment with the estrogen receptor down-regulator ICI 182,780, both in the brain () and in male gonads () of lizards collected in the period 2008–2017.
The anti-estrogen inhibits the expression of esr2 in the brain, showing levels not statistically different from those in the winter stasis period, in line with the histological results. Along the same line, in the male gonads, treatment with ICI 182,780 is shown to be effective as evidenced by expression levels in the treated animals being lower than in animals in the mating period, but injected with vehicle only; and these lower levels were also not significantly different from those of the winter stasis period.
Discussion
In this study, we first analyzed the partial sequence of the mRNA for esr2, which we identified in the lizard Podarcis sicula (deposited under the accession number GenBank JN705543.1) looking for sequence homologies and confirming the evolutionarily well-conserved domains. Observing in detail the amino acid sequence of esr2 of Podarcis sicula, we clearly recognize the three classical domains of the estrogen receptor superfamily (Guerriero et al. Citation2009; Mahfouz et al. Citation2016). Estimating the evolutionary distance between the partial sequence of esr2 of the Podarcis sicula lizard and the sequences of esr2 deposited in the BLAST database, it emerges, as expected, that the sequence esr2 of Podarcis sicula is placed among the reptiles. The sequence alignments also indicated that the lizard sequence is close to the birds’ orthologs but, interestingly, closer to the mammals than to amphibians and fishes (data not shown). In particular, an overall 76% sequence identity between human and lizard esr2 sequences emerged. It is interesting, then, to emphasize how the conserved DNA-binding domain shows an identity of 99% with Homo sapiens as well as to the closest species, clearly indicating a strict correlation of structure/function in DNA binding and recognition of target genes.
Further, we demonstrate that Podarcis sicula estrogen receptor is actively transcribed in the brain as in another endocrine tissue already reported, the testis (Ciarcia Citation1993; Verderame et al. Citation2014). The lowest gene expression levels were detected in the brain with the concomitantly highest esr2 transcription levels in testis during mating. Moreover, comparison of esr2 gene expression during the mating period compared to the winter stasis of the reproductive phase showed upregulation in the testis and downregulation in the brain. Those seasonal variations in gene expression followed the estradiol pattern in Podarcis sicula testis (Ciarcia Citation1993), and this result is in agreement with various studies showing that testes produce estradiol and that esr2 is very sensitive to estradiol changes – as, for example, in mammal rat testis, where ER-BETA is detected in spermatogenetic cells of various stages and in Sertoli cells, suggesting that estrogens directly affect germ and somatic cells during testicular development and spermatogenesis (Oliveira et al. Citation2001; Cooke et al. Citation2017). Podarcis is one bioindicator species for estradiol exposure altering esr2 expression as detected by in situ hybridization studies by Verderame et al. (Citation2014), and estrogen receptor mRNA using real-time PCR in primary cultures of lizard testis cells (Cardone et al. Citation1998). Our laboratories have demonstrated that there are close dynamics between brain and testis and that seasonal correspondence in the main phases of the reproductive cycle follows the feedback of the brain-gonad axis. Podarcis sicula, as a seasonal breeder, allowed us to better characterize how the gene esr2 is regulated in the brain of this lizard.
In order to study the expression levels of esr2 from Podarcis sicula, we investigated, by qRT-PCR, the esr2 expression in both the gonads and the brain of adult male lizards collected during the winter stasis and the mating period. The histological analysis of the tissues allowed verification of the expected tissue organization of the relative reproductive period and the effect of anti-estrogen treatments.
The analysis of the expression of esr2 in the reptile Podarcis sicula, by qRT-PCR, highlighted an interesting seasonal dynamic between the brain and the gonads, as in mammalian species (Pentikainen et al. Citation2000; Cooke et al. Citation2017). In the lizard, levels of estradiol show seasonal variations in accordance with the reproductive cycle via ER-BETA (Chieffi & Varriale Citation2004). The estrogen receptor levels are modulated by a series of complex processes that involve the control of expression and post-transcriptional modifications (Guerriero et al. Citation2005b, Citation2009; Greytak & Callard Citation2007; Tohyama et al. Citation2016; Cooke et al. Citation2017).
Our present and previous results indicate that the testis should be considered a valid tissue model for endocrine disruption assessment. This idea may be supported by the high conservation degree of the sequences of the DBD and LDB domains and by the spatial organization of germ cells. An interesting confirmation is provided by the data of Cohen et al. (Citation2012) in the lizard Anolis, that show a similar neuro-distribution in the brain between vertebrates as also confirmed by Guerriero et al. (Citation2009) and Mahfouz et al. (Citation2016). The work by Cohen et al. (Citation2012) examines reptile brain ER-BETA distribution, documenting a similar pattern to that found in birds and mammals, which suggests that this receptor may perform similar functions across multiple vertebrate taxa, whereas lizard testis coexpresses androgen receptor and estrogen receptor alpha and beta (Verderame et al. Citation2014). Further studies are needed to elucidate the general vertebrate mechanism underlying esr2 actions and fertility, but there is strong evidence of ER-BETA predominance within the seminiferous tubules in the early gametogenesis event (O’Donnel et al. Citation2001).
In recent years, many high-persistence molecules, defined as emerging contaminants, have become of concern because of their ability to disrupt the endocrine system and their recalcitrance in the environmental matrix (Santos et al. Citation2010). Particular importance has been given to pharmaceutical products derived not only from human but also from veterinary use. Among the therapeutic classes, antibiotics, epileptic agents and sex hormones are predominant (Santos et al. Citation2010). Steroids such as the synthetic hormone 17 alpha estradiol (main hormone of the small contraceptive) and the natural hormone 17 beta estradiol (used as a drug in hormone replacement therapy) are added to the antisteroids; in particular the antiestrogens, based on their mechanism of action, are classified into impeded antagonists, such as estradiol, which interacts with ER but rapidly dissociates; aromatase inhibitors, such as anastrozole, which block the conversion of androgens into estrogens; the triphenylethylene antagonists which in turn are divided into two families: the selective ER modulators, such as the drug Nolvadex, and the selective destroyers of ER, such as Faslodex whose bioactive molecule is ICI 182,780. Our attention has focused on ICI 182,780. It is known that estradiol administration stimulates proliferation of spermatogonia in otherwise inactive testes in teleosts, amphibians and reptiles, and in each of these classes, tamoxifen and ICI 182,780 prevent this stimulation process (Minucci et al. Citation1995; Guerriero et al. Citation2000; Chieffi et al. Citation2002). On the basis of the human species’ esr2 identity with Podarcis sicula, we treated the bioindicator Podarcis sicula with ICI 182,780. ICI 182,780 is a 7-alkylsulfinyl, an analog of endogenous estrogen 17-beta-estradiol. It binds to estrogen receptors with high affinity, preventing binding with estrogens. In this way it prevents the receptor’s dimerization and promotes its degradation, causing the abrogation of the transcription of the sensitive estrogen genes (Morris & Wakeling Citation2002). The damage caused by ICI 182,780 at the brain level is already known (Chieffi et al. Citation2002; Alfinito et al. Citation2008; Guerriero et al. Citation2012; Gao et al. Citation2016) and can be studied following the sensitivities of several different biomarkers and approaches (Guerriero et al. Citation2003, Citation2005a,Citationb; Bartiromo et al. Citation2013; Guerriero et al. Citation2018a,Citation2018b; D’Errico et al. Citation2018). In particular, morphological studies have shown that ICI 182,780 changes the hypophysis histomorphometrically with an increase and/or reduction of its weight (Gao et al. Citation2016), and increases the concentration of calcium channels (Zhao et al. Citation2006). Biochemical studies have allowed detection of alterations of the lactate dehydrogenase pattern (Nunez & McCarthy Citation2003) and extracellular phosphorylation of the kinases ERK1/2 (Wong et al. Citation2003). Indeed, through molecular studies it was possible to demonstrate that ICI 182,780 acts on the antioxidant defense, whereby after treatment with ICI 182,780 there is an increase in the concentration of the PHGPx\gpx4 mRNA, indicating the presence of a damage shelter (Guerriero et al. Citation2012), and this alters estrogen receptor expression in mammals (Gao et al. Citation2016). Our present results indicate that anti-estrogen treatment has a detrimental effect on spermatogenesis too via esr2 brain expression as already detected in the mammalian species by Oliveira et al. (Citation2001). This allowed us to verify that by interfering with the maturation process with an endocrine disruptor treatment such as ICI 182,780 for 21 days, Podarcis sicula gonads and brain obtained esr2 levels closer to winter stasis. Assessing changes in estrogen receptor expression in animals exposed and not exposed to endocrine disruptor compounds is critical for our understanding of the role of these receptors in endocrine disruptors in the natural environment (Rie et al. Citation2005; Guerriero Citation2011; Guerriero et al. Citation2014; Verderame et al. Citation2014). In this study, not only has a histological alteration been noted, but also the return of expression levels of esr2 to values not significantly different from winter stasis for both the gonads and the brain. Thus, the role of esr2 in the control of reproductive function suggests its use as a biomarker of anti-estrogen damage.
Conclusions
In the current study, we show that the esr2 protein sequence of the lizard Podarcis sicula is evolutionarily very conserved, with a high sequence identity with the human ortholog. Furthermore, we report the transcription activity of esr2 by qRT-PCR in brain and testis in the main reproductive phases, i.e. during the period of maximum activity of gonads, and in winter stasis. The observed pattern will help us to both detect the normal feedback within the established brain-gonad dynamics when there is interference by anti-estrogen compounds, and to identify variability within this dynamic.
These findings will allow the use of esr2 as a biomarker in the bioindicator Podarcis sicula for biomonitoring environmentally restored sites, and further allow the development of quick and efficient tests by gonadectomy in pollution biomonitoring programs. This methodology may also be used for making predictions of risk based on future mathematical models. Certainly, the high conservation of the sequence of esr2 and the general cytological organization close to that of humans strongly suggest the use of Podarcis sicula as an excellent model to monitor the sexual maturation state following exposure to various substances, thus opening new horizons for the conservation of species, including the human species.
Acknowledgements
The authors gratefully thank Dr. Emidio Sivieri, Biomedical Engineer at Children’s Hospital Of Philadelphia, Section on Newborn Care, at Pennsylvania Hospital, Philadelphia (USA), for critical revision of the manuscript; Ms. Dhouha Mabrouk for careful proofreading and editing of the manuscript; and the Interdepartmental Research Center for Environment (I.R.C.Env.), Federico II University, Napoli, Italy for logistical help.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Alfinito PD, Chen X, Atherton J, Cosmi S, Deecher DC. 2008. ICI 182,780 Penetrates brain and hypothalamic tissue and has functional effects in the brain after systemic dosing. Endocrinology 149:5219–5226. DOI:10.1210/en.2008-0532.
- Andò S, Ciarcia G, Panno ML, Imbrogno E, Tarantino G, Buffone M, Beraldi E, Angelini F, Botte V. 1992. Sex steroids levels in the plasma and testis during the reproductive cycle of lizard Podarcis s. sicula Raf. General and Comparative Endocrinology 85:1–7.
- Arevalo MA, Santos-Galindo M, Lagunas N, Azcoitia I, Garcia-Segura LM. 2011. Selective estrogen receptor modulators as brain therapeutic agents. Journal of Molecular Endocrinology 46:R1–R9.
- Bartiromo A, Guignard G, Barone Lumaga MR, Barattolo F, Chiodini G, Avino R, Guerriero G, Barale G. 2013. The cuticle micromorphology of in situ Erica arborea L. Exposed to Long-Term Volcanic Gases. Environmental and Experimental Botany 87:197–206.
- Blatter MS, Mahoney MM. 2015. Changes in estrogen receptor signaling alter the timekeeping system in male mice. Behavioural Brain Research 294:43–49.
- Cao C, Ding Y, Kong X, Feng G, Xiang W, Chen L, Yang F, Zhang K, Mingxing CM, Wang P, Zhang B. 2018. Reproductive role of miRNA in the hypothalamic-pituitary axis. Molecular and Cellular Neuroscience 88:130–137.
- Cardone A, Angelini F, Varriale B. 1998. Autoregulation of estrogen and androgen receptor mRNAs and downregulation of androgen receptor mRNA by estrogen in primary cultures of lizard testis cells. General and Comparative Endocrinology 110:227–236.
- Chieffi P, Colucci D’Amato GL, Guarino F, Salvatore G, Angelini F. 2002. 17-b Estradiol induces spermatogonial proliferation through mitogen-activated protein kinase (extracellular signal-regulated kinase 1) activity in the lizard (Podarcis s. sicula). Molecular Reproduction and Development 61:218–225.
- Chieffi P, Varriale B. 2004. Estrogen receptor β localization in the lizard (Podarcis s. sicula) testis. Zygote 12:39–42.
- Ciarcia G. 1993. Control of testicular activity in the lizard, Podarcis s. sicula Raf. In: Facchinetti F, Henderson IW, Pierantoni R, Polzonetti-Magni AM, editors. Cellular communication in reproduction. Bristol: Journal of Endocrinology, Ltd. pp. 131–140.
- Cohen RE, Roach J, Wade J. 2012. The distribution of estrogen receptor β mRNA in male and female green anole lizards. Brain Research 1430:43–51.
- Cooke PS, Nanjappa MK, Ko C, Prins GS, Hess RA. 2017. Estrogen in male physiology. Physiological Reviews 97:995–1043.
- D’Errico G, Vitiello G, De Tommaso G, Abdel-Gawad FKH, Brundo MV, Ferrante M, De Maio A, Trocchia S, Bianchi AR, Ciarcia G, Guerriero G. 2018. Electron Spin Resonance (ESR) for the study of eactive Oxygen Species (ROS) on the isolated frog skin (Pelophylax bergeri): A non-invasive method for environmental monitoring. Environmental Research 165:11–18. DOI:10.1016/j.envres.2018.03.044.
- Gao H, Xue Y, Cao L, Liu Q, Liu C, Shan X, Wang H, Gu Y, Zhang Y. 2016. ESR1 and its antagonist fulvestrant in pituitary adenomas. Molecular and Cellular Endocrinology 1e10. 443:32–41.
- Greytak SR, Callard GV. 2007. Cloning of three estrogen receptors (ER) from killifish (Fundulus heteroclitus): Differences in populations from polluted and reference environments. General and Comparative Endocrinology 150:174–188.
- Gronemeyer H, Laudet V. 1995. Transcription factors 3: Nuclear receptors. Protein Profile 2:1173–1308.
- Guerriero G. 2007. Seasonal steroids variations and maturity stages in the female chub, Leuciscus cephalus L. (Pisces, Cyprinidae). Italian Journal of Zoology 74:317–324.
- Guerriero G. 2009. Vertebrate sex steroid receptors: Evolution, ligands, and neurodistribution. Annals of the New York Academy of Sciences 1163:154–168.
- Guerriero G. 2011. Hormones in animal biology. Italian Journal of Zoology 2. Editorial. 78:133.
- Guerriero G, Brundo MV, Labar S, Bianchi AR, Trocchia S, Rabbito D, Palumbo G, Abdel-Gawad FK, De Maio A. 2018a. Frog (Pelophylax bergeri, Gunther 1986) endocrine disruption assessment: Characterization and role of skin poly(ADP-ribose) polymerases. Environmental Science and Pollution Research 25: 18303. DOI:10.1007/s11356-017-0395-2.
- Guerriero G, Ciarcia G. 2001. Progesterone receptor: Some viewpoints on hypothalamic seasonal fluctuations in a lower vertebrate. Brain Research Reviews 37:172–177.
- Guerriero G, D’Errico G, Di Giaimo R, Rabbito D, Olanrewaju OS, Ciarcia G. 2018b. Reactive oxygen species and glutathione antioxidants in the testis of the soil bi-osentinel Podarcis sicula (Rafinesque 1810). Environmental Science and Pollution Research 25: 18286. DOI:10.1007/s11356-017-0098-8.
- Guerriero G, Di Finizio A, Ciarcia G. 2003. Oxidative defenses in the sea bass, Dicentrarchus labrax. In: Dunn JF, Swartz HM, editors. Handbook of oxygen transport to tissue. XXIV. Advances in Experimental Medicine and Biology. Berlin, Germany: Springer. Vol. 530, pp. 681–688.
- Guerriero G, Ferro R, Ciarcia G. 2005b. Correlation between plasma levels of sex steroids and spermatogenesis during the sexual cycle in the chub, Leuciscus cephalus L. (Pisces: Cyprinidae). Zoological Studies 44:228–233.
- Guerriero G, Ferro R, Russo GL, Ciarcia G. 2004. Vitamin E in early stages of sea bass (Dicentrarchus labrax) development. Comparative Biochemistry and Physiology A. Molecular and Integrative Physiology 138:435–439.
- Guerriero G, Prins GS, Birch L, Ciarcia G. 2005a. Neurodistribution of androgen receptor immunoreactivity in the male frog, Rana esculenta. Annals of the New York Academy of Sciences 1040:332–336.
- Guerriero G, Roselli CE, Ciarcia G. 2009. The amphibian (Rana esculenta) brain progesterone receptor: Relationship to plasma steroids andVvitellogenic cycle during the gonadal recovery phase. Annals of the New York Academy of Sciences 1163:407–409.
- Guerriero G, Roselli CE, Paolucci M, Botte V, Ciarcia G. 2000. Estrogen receptors and aromatase activity in the hypothalamus of the female frog, Rana esculenta. Fluctuations throughout the reproductive cycle. Brain Research 880:92–101.
- Guerriero G, Trocchia S, Abdel-Gawad FK, Ciarcia G. 2014. Roles of reactive oxygen species in the spermatogenesis regulation. Front Endocrinology 5:56.
- Guerriero G, Trocchia S, Ferrara C, Ciarcia G. 2012. Lizard neuroendocrine disruptor assessment by phospholipid hydroperoxide glutathione peroxidase (GPX4/PHGPx) expression. Experimental and Clinical Endocrinology 120:P5. DOI:10.1055/s-0032-1330090.
- Hawkins MB, Thornton JW, Crews D, Skipper JK, Dottiee A, Thomas P. 2000. Identification of a third distinct estrogen receptor and reclassification of estrogen receptors in teleosts. Proceedings of the National Academy of Sciences of the United States of America 97:10751–10756.
- Jensen M-B, Krarup JF, Palshof T, Ejlertsen B. 2018. Two years of tamoxifen or no adjuvant systemic therapy for patients with high-risk breast cancer: Long-term follow-up of the Copenhagen breast cancer trial. Acta Oncologica 57:26–30.
- Katsu Y, Bermudez DS, Braun EL, Helbing C, Miyagawa S, Gunderson MP, Kohno S, Bryan TA, Guillette LJ, Iguchi T. 2004. Molecular cloning of the estrogen and progesterone receptors of the American alligator. General and Comparative Endocrinology 136:122–133.
- Katsu Y, Taniguchi E, Urushitani H, Miyagawa S, Takase M, Kubokawa K, Tooi O, Oka T, Santo N, Myburgh J, Matsuno A, Iguchi T. 2010. Molecular cloning and characterization of ligand- and species-specificity of amphibian estrogen receptors. General and Comparative Endocrinology 168:220–230.
- Katsutoshi I, Kazuhiko M, Kazunori F. 2007. Molecular cloning of two estrogen receptors expressed in the testis of the Japanese Common Goby, Acanthogobius flavimanus. Zoological Science 24:986–996.
- Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C (T)). Methods 25:402–408.
- Mahfouz A, Boudewijn PF, Lelieveld A, Grefhorst A, van Weert Lisa TCM, Mol IM, Sips HCM, van den Heuvel JK, Datson NA, Visser JA, Reinders MJT, Meijer OC. 2016. Genome-wide coexpression of steroid receptors in the mouse brain: Identifying signaling pathways and functionally coordinated regions. Proceedings of the National Academy of Sciences of the United States of America 113:2738–2743.
- Mazzi A. 1977. Manuale di tecniche istologiche e istochimiche. Padova, Italy: Piccin.
- Minucci S, Vitiello II, Marmorino C, di Matteo L, Baccari GC. 1995. Mast cell-Leydig cell relationships in the testis of the lizard Podarcis s. sicula Raf: Thermal manipulation, ethane 1,2-dimethane sulphonate (EDS) and sex hormone treatment. Zygote 3:259–264.
- Morris C, Wakeling A. 2002. Fulvestrant (‘Faslodex’)–A new treatment option for patients progressing on prior endocrine therapy. Endocrine-Related Cancer 9:267–276.
- Nikolos F, Thomas C, Bado I, Gustafsson JÅ. 2018. ERβ Sensitizes NSCLC to chemotherapy by regulating DNA damage response. Molecular Cancer Research 16:233–242.
- Nunez JL, McCarthy MM. 2003. Estradiol exacerbates hippocampal damage in a model of preterm infant brain injury. Endocrinology 144:2350–2359.
- O’Donnel L, Robertson KM, Jones ME, Simpson ER. 2001. Estrogen and spermatogenesis. Endocrine Reviews 22:289–318.
- Oliveira CA, Carnes K, Franca LR, Hess RA. 2001. Infertility and testicular atrophy in the antiestrogen treated adult male rat. Biology of Reproduction 65:913–920.
- Pentikainen V, Erkkila K, Suomalainen L, Parvinen M, DunKel L. 2000. Estradiol acts as a germ cell survival factor in the human testis in vitro. Journal of Clinical Endocrinology and Metabolism 85:2057–2067.
- Raven N, Crkvenjakov R, Boedtker H. 1979. Identification of procollagen mRNAs transferred to diazobenzyloxymethyl paper from formaldehyde agarose gels. Nucleic Acids Research 6:3559–3567.
- Rie MT, Kitana N, Lendas KA, Won SJ, Callard IP. 2005. Reproductive endocrine disruption in a sentinel species (Chrysemys picta) on Cape Cod, Massachusetts. Archives of Environmental Contamination and Toxicology 48:217–224.
- Santos LH, Araujo AN, Fachini A, Pena A, Delerue-Matos C, Montenegro MC. 2010. Ecotoxicological aspects related to the presence of pharmaceuticals in the aquatic environment. Journal of Hazardous Materials 175:45–95.
- Tohyama S, Miyagawa S, Lange A, Ogino Y, Mizutani T, Ihara M, Tanaka H, Tatarazako N, Kobayashi T, Tyler CR, Iguchi T. 2016. Evolution of estrogen receptors in ray-finned fish and their comparative responses to estrogenic substances. Journal of Steroid Biochemistry and Molecular Biology 158:189–197.
- Verderame M, Angelini F, Limatola E. 2014. Spermatogenic waves and expression of AR and ERs in germ cells of Podarcis sicula. International Journal of Zoology Article ID 965617:8.
- Verderame M, Limatola E. 2010. Molecular identification of estrogen receptors (ERα and ERβ) and their differential expression during VTG synthesis in the liver of lizard Podarcis sicula. General and Comparative Endocrinology 168:231–238.
- Walker DM, Gore AC. 2017. Epigenetic impacts of endocrine disruptors in the brain. Front Neuroendocrinology 44:1–26.
- Wong JK, Le HH, Zsanovszky A, Belcher SM. 2003. Estrogens and ICI 182,780 (Faslodex) modulate mitosis and cell death in immature cerebellar neurons via rapid activation of p44/p42 mitogen-activated rrotein kinase. Journal of Neuroscience 23:4984–4995.
- Zhang H, Chen F, Li GL, Ding YY, Tao ZR, Li JJ, Zhong SL, Lu LZ. 2014. Molecular cloning, expression, and regulation of estrogen receptors in pigeon oviduct epithelial cells. Genetics and Molecular Research 13:1926–1937.
- Zhang H, Xie M, Schools GP, Feustel PF, Wang W, Lei T, Kimelberg HK, Zhou M. 2009. Tamoxifen mediated estrogen receptor activation protects against early impairment of hippocampal neuron excitability in an oxygen/glucose deprivation brain slice ischemia model. Brain Research 1247:196–211.
- Zhang R, Yin Y, Sun L, Yan P, Zhou Y, Wu R, Wu X. 2017. Molecular cloning of ESR2 and gene expression analysis of ESR1 and ESR2 in the pituitary gland of the Chinese alligator (Alligator sinensis) during female reproductive cycle. Gene 623:15–23.
- Zhang Y, Milatovic D, Aschner M, Feustel PJ, Kimelberg HK. 2007. Neuroprotection by tamoxifen in focal cerebral ischemia is not mediated by an agonist action at estrogen receptors but is associated with antioxidant activity. Experimental Neurology 204:819–827.
- Zhao L, O’Neill K, Brinton RD. 2006. Estrogenic agonist activity of ICI 182,780 (Faslodex) in hippocampal neurons: Implications for basic science understanding of estrogen signaling and development of estrogen modulators with a dual therapeutic profile. Journal of Pharmacology and Experimental Therapeutics 319:1124–1132.