
Abstract
The seeds of Norway spruce (Picea abies) are important food for the Great Spotted Woodpecker (Dendrocopos major), especially in winter, when other resources are limited. However, little is known about preferences of woodpeckers in terms of the location of anvils, in which seeds of Norway spruce are extracting from cones. In this study, we documented preferences of the Great Spotted Woodpecker for anvil placement sites and checked whether there are differences between males and females in this respect. The research was conducted in primeval oak-lime-hornbeam stands of the Białowieża National Park, E Poland. To determine preferences of woodpeckers regarding the species and condition of trees in terms of the location of anvils in our research we took into consideration available resources of trees. Altogether during our study, we collected data on 358 anvils. They were most often located on large trees, having a much larger trunk diameter than the average diameter of trees from resources. We found the greatest number of anvils on oak (30%), which was also the most preferred tree species by woodpeckers. Anvils were located the most often on branches (72%), on dead parts of trees (67%) and at great heights (on average over 19 m above ground). We found no differences between males and females in the selection of anvil placement sites.
Introduction
The Great Spotted Woodpecker (Dendrocopos major) is the most common and the most omnivorous among all European woodpecker species. In some regions, conifer seeds are an essential component of its diet (Hogstad Citation1971; Michalek & Miettinen Citation2003). They are particularly important in periods when other resources (i.e. food of animal origin) are limited, e.g. in winter (Hogstad Citation1971; Osiejuk Citation1998). The Great Spotted Woodpecker most often feeds on seeds of Scots pine (Pinus silvestris) and Norway spruce (Picea abies), however, occasionally it may also feed on seeds of other tree species (Hordowski Citation1995; Michalek & Miettinen Citation2003; Nicolai Citation2011; Dylewski et al. Citation2017; Dylewski & Myczko Citation2019). Woodpeckers extract seeds from cones using anvils, in which they put a cone after removing it from a tree. Such a place has to provide a stable wedging of a cone, which prevents it from falling out during feeding. Most often cracks and crevices in the bark or fractures formed in branches as a result of breaking off or decay of their parts become anvils. The size of a cone is the primary factor determining the choice of a specific location for an anvil, because the cone has to fit into it (Hordowski Citation1995; Kędra & Mazgajski Citation2001). Therefore, one woodpecker can use many anvils of different sizes, which are selected depending on the dimensions of picked cones (Hordowski Citation1995). In addition, too small, naturally formed crevices can be enlarged by woodpeckers to fit the size of cones hammered into them (Hordowski Citation1995; Michalek & Miettinen Citation2003). Individual anvils may be used by woodpeckers with varying intensity. Some of them are used regularly and frequently, while others only occasionally or even just once (Kędra & Mazgajski Citation2001; Gorman Citation2015). Among the European woodpeckers, it is mainly the Great Spotted Woodpecker that uses anvils, although other species, e.g. the White-backed Woodpecker Dendrocopos leucotos can also use them occasionally (Cramp Citation1985; Gorman Citation2015).
Although foraging behaviour of the Great Spotted Woodpecker was often studied, the topic of anvils was rarely discussed and mainly concerned anvils used for Scots pine cones in managed, pine-dominated forests (Skoczylas Citation1961; Osiejuk Citation1991; Hordowski Citation1995; Kędra & Mazgajski Citation2001). Whereas the area of our study was the Białowieża Forest, the best-preserved natural European lowland forest, which provides excellent conditions for the observation of organisms in their natural habitats, and the research conducted there also allows us to draw conclusions regarding the evolution of various adaptations and behaviours of animals under natural conditions. The Great Spotted Woodpecker is very common there and reaches density up to 2.0 pair/10 ha (Wesołowski et al. Citation2015a).
The first of the objectives of our survey was to characterise the locations of anvils used by the Great Spotted Woodpecker to extract seeds from Norway spruce cones. This tree species is present in all types of tree stands in the Białowieża Forest, both as a dominant species in coniferous (spruce-pine or spruce) stands and as an admixture in oak-lime-hornbeam, ash-alder or swampy alder stands (Pawlaczyk Citation2009). Therefore, seeds of Norway spruce are important food for the Great Spotted Woodpecker in the Białowieża Forest, constituting in oak-lime-hornbeam stands during some autumn-winter seasons almost the sole component of its diet (Stański Citation2014).
The next aim of our study was to determine the preferences of the Great Spotted Woodpecker for anvil placement sites. Primeval stands of the Białowieża Forest are characterised by an unusual tree species richness and possess diversified vertical and horizontal structure, therefore, there is a large number of potential places where anvils can be located, and some trees may be particularly preferred on account of their species or condition. In order to determine these preferences, trees selected for the location of anvils were compared with available resources. We hypothesised that trees with large dimensions (i.e. large trunk diameter at breast height) would be especially preferred because they possess many potential places to locate anvils (crevices, dead branches).
Moreover, the objective was also to check whether males and females of the Great Spotted Woodpecker locate anvils in places with the same parameters. Foraging niche differentiation between sexes has been found in some European woodpecker species, such as the Middle Spotted Woodpecker Leiopicus medius (Pasinelli Citation2000), the White-backed Woodpecker (Stenberg & Hogstad Citation2004), or the Three-toed Woodpecker Picoides tridactylus (Pechacek Citation2006). Intersexual niche partitioning and differences in foraging behaviour were also revealed in the case of the Great Spotted Woodpecker. Osiejuk (Citation1994) revealed that females compare to males were more mobile and used more techniques and microhabitats during foraging. Taking into account the results of the above-mentioned studies showing that foraging places may be different for males and females, we predicted that anvil placement sites would differ between both sexes. To our knowledge, this is the first publication comparing the use of anvils by two sexes of the Great Spotted Woodpecker.
Materials and methods
Study area
The study was carried out in the Białowieża Forest (52°29ʹ – 52°57ʹN; 23°31ʹ – 24°21ʹE), a large forest complex located at the border between Poland and Belarus. The best preserved Polish part of this forest is protected as the Białowieża National Park (hereafter BNP) and covers 105 km2. The essential part of BNP has never been logged and possesses many characteristics of a primeval forest, such as multi-storey, mixed-species and uneven-aged tree stands with large amounts of dead wood (Tomiałojć Citation1991; Bobiec Citation2002). The study area (about 10 km2) was located in BNP in the primeval oak-lime-hornbeam stands, Tilio-Carpinetum, which is the dominant type of forest there, covering c. 45% of its total area. These stands are the most structurally diversified forest types with three canopy layers and many tree species. The most numerous tree species in these stands are lime Tilia cordata, hornbeam Carpinus betulus, Norway spruce, pedunculate oak Quercus robur, maple Acer platanoides, ash Fraxinus excelsior and elms Ulmus spp. Live spruces constitute about 10% of all trees in oak-lime-hornbeam stands, with old trees being the most common because the species is currently poorly regenerating there.
Data collection
Data on anvils were collected from 1999 to 2009 (except 2002). Anvils were searched for during random walks in the study area, where the sounds of woodpeckers hammering on cones were helpful. We recorded data only when the Great Spotted Woodpecker was spotted while extracting seeds from Norway spruce cones and only such data were included in the analysis. We did not record parameters of anvils that were detected solely on the basis of cones left in them or cones present under a tree. Taking such sites into account would increase the number of easily detectable anvils located in visible places, i.e. on the trunks and low on trees. When the Great Spotted Woodpecker was found feeding on a cone, the sex of a foraging bird was determined and parameters of an anvil were recorded: (1) tree species, (2) tree condition (alive or dead), (3) tree diameter at breast height (DBH), (4) part of the tree used (trunk or branch), (5) condition of the part used (alive or dead), (6) substrate diameter: up to 5 cm, >5–10 cm, >10–20 cm, >20 cm, (7) height above ground.
In order to estimate the DBH, the circumference of the trunk was measured (using a tape measure), on the basis of which the DBH was calculated. The diameter of a site where an anvil was located was visually assessed using the woodpecker body size as a reference, whereas the height of an anvil location was measured using a Suunto clinometer or the height of an observer as a reference.
Habitat measurements
To determine the trees preferred by woodpeckers as an anvil location, we measured from 1999 to 2003 the habitat on 82 square sample plots (0.25 ha each) randomly selected in the study area. On these plots we recorded all standing trees, noting their species, condition (alive or dead) and DBH (). The average number of measured trees on the plot was 89.7 (±27.9 standard deviation).
Table I. Tree species selected for the anvils by males, females and all woodpeckers
Data analysis
Data from different years were pooled. During our study, we collected data on 358 anvils: 119 used by males, 155 by females, and 84 by birds whose sex was not recorded. The average number of found anvils during one year was 36, although it ranged from 1 in 1999 to 94 in 2004. Such difference was caused by extreme variation in Norway spruce seeds production in particular years (Wesołowski et al. Citation2015b). Thus, the Great Spotted Woodpecker in seasons, when seeds were abundant, foraged mainly on them, whereas in seasons with low availability of Norway spruce cones chose other food resources (Stański Citation2014).
Analyses were performed and characteristics were presented for all anvils together, anvils used by males, and anvils used by females. To determine preferences of woodpeckers regarding the species and condition of trees in terms of the location of anvils, selection indices were calculated. They were calculated as a percentage of trees on which anvils were located in relation to a percentage of available trees (Manly et al. Citation2002). Trees with a DBH of at least 13 cm (the minimum DBH of trees on which anvils were located in our study) were considered as available trees. If the selection index was greater than one, it indicated preference, if the index was less than one, it indicated negative selection. Moreover, for each selection index, we constructed 95% simultaneous confidence limits according to Manly et al. (Citation2002). We changed negative lower limits to 0.00, because negative values of confidence limits are not possible. The selection index was considered to be statistically significant if the calculated confidence limits did not contain the value 1 (Manly et al. Citation2002).
In order to check if the thickness of trees selected for the location of anvils differs from the available trees, we used the Mann–Whitney U-test. The analysis was performed for all birds together, without sex breakdown, and only for the most commonly used tree species.
To assess whether there are differences in anvil location between males and females, we applied the Generalised Linear Model (GLM) with binomial error distribution and the logit link function. The sex was considered as a binary response variable, with 0 indicating females and 1 indicating males. The model included a set of variables that characterised the location of an anvil. Tree species, tree condition (alive or dead), the type of place (trunk or branch), its condition (alive or dead) and the diameter were included as categorical explanatory variables, whereas DBH and the height of the anvil location – as continuous explanatory variables. Statistical analyses were carried out using STATISTICA 12.0. Results with p < 0.05 were considered statistically significant and all presented probability values are two-tailed.
Results
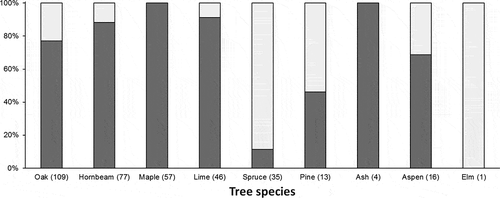
The Great Spotted Woodpecker used nine tree species for anvil location (eight in the case of males and nine in the case of females). In general, both sexes used individual tree species with similar intensity. The largest number of anvils were located on oaks (about 30%), but many of them were also found on hornbeams and maples, whereas the least on ashes and elms. The selection indices showed statistically significant preferences for oaks and maples in all analysed groups (males, females and “all birds”). Limes and hornbeams, although numerous in the stand, were avoided ().
Both males and females located most anvils on living trees but the selection indices showed a strong preference for dead trees (). Anvils were more frequently found on dead trees in the case of coniferous species (Norway spruce, Scots pine) compared to deciduous ones ().
Figure 1. Percentage distribution of anvils on alive (dark colour) and dead (pale colour) trees from particular species. Numbers in brackets indicate sample size
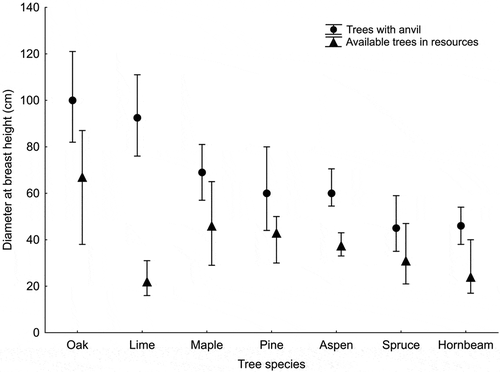
Trees with anvils had a significantly larger DBH than available trees in the case of all analysed species: oaks (Mann–Whitney U test; Z = 8.98; p < 0.001), hornbeams (Z = 9.70; p < 0.001), maples (Z = 6.78; p < 0.001), limes (Z = 10.64; p < 0.001), Norway spruces (Z = 3.96; p < 0.001), Scots pines (Z = 2.50; p = 0.012) and aspens (Z = 5.51 p < 0.001; ).
Figure 2. Median diameter at breast height (DBH) of trees with anvils compared to available trees in resources from particular species. Whiskers indicate 25–75% quartile ranges
Most anvils (about 70%) were located on branches and dead parts of trees (above 60%). The largest number of anvils (over 40%) were located on parts of trees between 5 and 10 cm wide, while the least on parts below 5 cm wide. Almost 80% of the anvils were located above 15 m (). The average height of anvil location was 19.4 m for all birds, 19.5 m for males and 19.2 m for females. The lowest location of an anvil was at a height of 0.3 m, and the highest at 41 m.
Table II. Number (N) and percentage distribution (%) of anvils in relations to: type of substrate, its condition, diameter and height
Detailed information about DBH of trees with anvils and height of anvil location in relation to tree species is shown in .
Table III. The diameter at breast height (DBH) of trees with anvils and height of anvil location in relation tree species
GLM showed no differences between males and females in the selection of anvil placement sites ().
Table IV. Results of generalised linear model assessing the differences between males and females of the Great Spotted Woodpecker in the selection of anvil placement sites
Discussion
Our study showed that anvils where seeds were extracted from Norway spruce cones by the Great Spotted Woodpeckers were most often located on large trees, at great heights and usually on dead branches. On the other hand, anvils commonly considered to be “typical”, i.e. located at low heights on tree trunks (Skoczylas Citation1961; Osiejuk Citation1991; Hordowski Citation1995) constituted a small percentage of all anvils found.
Tree species particularly preferred by woodpeckers for anvil location were oaks and maples. Both of these species reach considerable sizes in BPN and therefore have many natural cracks, crevices and dead branches, which are perfect for the location of anvils. In scientific publications, there is no information on the use of maples as anvil sites, while frequent selection of oak trees has been noted by other authors. For example, out of 13 anvils found by Osiejuk (Citation1991), 12 were on oaks. Hordowski (Citation1995) in his research conducted in coniferous and mixed stands with a predominance of Scots pine showed that in this respect oak was the most frequently used tree after the alder. A significant contribution of anvils on oaks (29%) was also observed in forests located on the outskirts of the Kampinos National Park (near Warsaw), despite the fact that Scots pine definitely dominated in the stands, and its contribution was about 90% (Kędra & Mazgajski Citation2001).
Many anvils in BNP were located on hornbeams and limes, however, according to the selectivity indices, these tree species were avoided by woodpeckers. This was due to the presence of a large number of young trees of these species in the stands, which, due to their age, did not have places suitable for anvil location.
Anvils in the BNP oak-lime-hornbeam forests were most often located on large trees, having a much larger DBH than the average DBH of available resources. This is due to the fact that such trees have a large number of places suitable for anvils, such as dead branches or natural crevices (Izdebska Citation2010; Michałowska Citation2010). There are also more of these places on dead trees compared to living ones, which explains why the former were used more than three times as often as their resources would indicate. Moreover, our study showed that anvils in the case of coniferous trees were much more often located on dead trees (as much as 90% in the case of spruce) compared to deciduous species. Live Norway spruces and Scots pines, characterised by hard wood saturated with resin, usually have no crevices or other places suitable for locating cones, whereas such places are most often found in the case of dead specimens of these species.
Our study revealed that most anvils were located in dead parts of trees, which, apart from the fact that it is possible to find there suitable locations for cones, is attributable to the fact that rotten wood can easily be enlarged by woodpeckers to adapt anvils to their needs. Such enlarged anvils can account for the majority of anvils in a given area (Hordowski Citation1995).
The anvils were located at great heights in BNP, on average over 19 m above ground and almost 80% of them were placed above 15 m above ground, i.e. much higher than reported by other authors: in the Kampinos Forest, the anvils were located on average at a height of about 10 m (Skoczylas Citation1961), in the Przemyskie Foothills – about 8.6 m (Hordowski Citation1995), on the Vistula Spit – 7.6 m (Osiejuk Citation1991). Such a great height of placing anvils in BPN may be due to the fact that spruces growing there reach considerable sizes, so cones are located quite high on them. Thanks to the location of anvils at a considerable height, flights between them and the source of feed do not require overcoming large differences in height, which would be an inefficient feeding strategy (Osiejuk Citation1991). In addition, the great height probably means more places suitable for locating cones (i.e. more medium-sized dead or rotten branches), as well as better protection against predators, whose high diversity occurs in BPN (Tomiałojć Citation1991).
In our study, we revealed that about 70% of the anvils were located on branches and 30% on trunks, unlike other authors: Hordowski (Citation1995) found 80% of anvils on the trunks, Osiejuk (Citation1991) – two-thirds of all anvils, while Skoczylas (Citation1961) – up to 100%.
The differences in the location of anvils in BNP and the results obtained by the above-mentioned authors are primarily due to different habitats of the study areas. Skoczylas (Citation1961), Osiejuk (Citation1991), Hordowski (Citation1995) carried out their research in forests where the Scots pine had a significant or even dominant contribution and its cones were hammered in the analysed anvils. The stands in these forests are not as diverse in terms of structure, size and number of tree species as in the Białowieża National Park. For this reason, the latter offers a wide spectrum of potential places where anvils can be located. The differences in the location of anvils are certainly due to the fact that the cited studies concerned Scots pine cones and those carried out in the BNP concerned Norway spruce cones. These cones differ significantly in size, with which the anvil size is closely correlated (Hordowski Citation1995; Kędra & Mazgajski Citation2001). There are also different factors that determine the choice of cones of these species in order to obtain seeds. The Great Spotted Woodpecker avoids Scots pine cones with large apophyses and prefers medium-sized cones with smaller apophyses (Myczko & Benkman Citation2011). In the case of Norway spruce, this woodpecker species chooses medium cones (between 90 and 150 mm) avoiding smaller and larger cones (Dylewski et al. Citation2017). The choice of cones with such parameters can be explained by a compromise – larger cones may be more difficult to transport and place in an anvil, whereas small cones contain fewer seeds, many of which are underdeveloped, which makes feeding on them less profitable (Ehrenberg et al. Citation1955).
In our study, we found no differences between males and females in the selection of anvil placement sites. Both sexes used individual tree species with similar intensity, preferred dead and large diameter trees and placed anvils at similar heights. Our findings suggest that in terms of anvil selection intersexual competition does not exist. However, intersexual competition for food in the case of the Great Spotted Woodpecker may exist and some mechanisms that reduce it, have been demonstrated by other authors. For example, Osiejuk (Citation1994) revealed that females of this species used more foraging techniques, more microhabitats and they were more mobile than males. Moreover, this author showed that differences in foraging behaviour between sexes were more pronounced in periods of limited food resources, i.e. in winter with low availability of Scots pine cones (Osiejuk Citation1998). On the other hand, Hogstad (Citation1978) found that both males and females of the Great Spotted Woodpecker showed a similar foraging pattern, but in order to minimise the intersexual competition both sexes use separate feeding territories in the non-breeding period.
The mechanism that minimises the intersexual competition can also be collecting food of different sizes by males and females, as observed in the Hairy Woodpecker Dendrocopus villosus (Kilham Citation1965). Such competition in the case of feeding on Norway spruce seeds would manifest itself in choosing cones of different sizes and thus anvils of different parameters by males and females. However, there is no information in the literature about differences in the phenotypic selection of cones between sexes of woodpeckers. On the other hand, the phenotypic selection of cones minimising the competition has been demonstrated for different species, i.e. the Great Spotted Woodpecker, which prefers feeding on larch cones with thinner peduncles and the red squirrel, which prefers feeding on small and medium larch cones (Dylewski & Myczko Citation2019).
In conclusion, we have demonstrated that anvils in the Białowieża National Park were most often located on large trees, at great heights and usually on dead branches. The preferred tree species for locating anvils by woodpeckers were oaks and maples.
Geolocation information
Poland, Białowieża National Park, coordinates 52°43ʹN; 23°50ʹE.
Acknowledgements
We thank the authorities of the Białowieża National Park for their help during our research. The study was supported by the Siedlce University of Natural Sciences and Humanities.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Bobiec A. 2002. Living stands and dead wood in the Białowieża forest: Suggestions for restoration management. Forest Ecology and Management 165:125–140. DOI: 10.1016/S0378-1127(01)00655-7.
- Cramp S. 1985. The Birds of the Western Palearctic. Vol. 4. Oxford: Oxford University Press.
- Dylewski Ł, Myczko Ł. 2019. Great spotted woodpecker (Dendrocopos major) and red squirrel (Sciurus vulgaris) prefer different con features of European larch (Larix decidua). Biologia 74:515–519. DOI: 10.2478/s11756-018-00181-z.
- Dylewski Ł, Yosef R, Myczko Ł. 2017. Difference on cone size preferences between two coniferous species by Great Spotted Woodpecker (Dendrocopos major). PeerJ 5:e3288. DOI: 10.7717/peerj.3288.
- Ehrenberg C, Gustafsson Å, Forshell C, Simak M. 1955. Seed quality and the principles of forest genetics. Hereditas 41:291–366. DOI: 10.1111/j.1601-5223.1955.tb02998.x.
- Gorman G. 2015. Foraging signs and cavities of some European woodpecker (Picidae): Identifying the clues that lead to establishing the presence of species. Denisia 36 Neue Serie 164:87–97.
- Hogstad O. 1971. Notes on the winter food of the great spotted woodpecker, Dendrocopos major. Sterna 10:233–241.
- Hogstad O. 1978. Sexual dimorphism in relation to winter foraging and territorial behaviour of the three-toed woodpecker Picoides tridactylus and three Dendrocopos species. Ibis 120(2):198–203. DOI: 10.1111/j.1474-919X.1978.tb06775.x.
- Hordowski J. 1995. Charakterystyka kuźni dzięcioła dużego (Dendrocopos major) w lasach mieszanych Pogórza Przemyskiego [Characteristics of the Great Spotted Woodpecker (Dendrocopos major) anvils in mixed forests of the Pogórze Przemyskie plateau]. Notatki Ornitologiczne 36:374–377.
- Izdebska M. 2010. Ile jest martwego na żywym? Obumieranie gałęzi na żywych dębach i ich znaczenie dla ptaków [How much of dead in alive? Dead branches and limbs of alive oaks and their importance for birds]. MS thesis, Siedlce University of Natural Sciences and Humanities, Siedlce.
- Kędra AH, Mazgajski TD. 2001. Factors affecting anvils utilization by Great Spotted Woodpecker Dendrocopos major. Polish Journal of Ecology 49:79–86.
- Kilham L. 1965. Differences in feeding behaviour of male and female hairy woodpeckers. Wilson Bulletin 77(2):134–145.
- Manly BFJ, McDonald LL, Thomas DL, McDonald TL, Erickson WP. 2002. Resource selection by animals. Statistical design and analysis for field studies. Dordrecht: Springer. DOI: 10.1007/0-306-48151-0.
- Michalek KG, Miettinen J. 2003. Dendrocopos major Great Spotted Woodpecker. BWP Update 5(2):101–184.
- Michałowska E. 2010. Obumieranie grabów i ich rola dla dzięciołów w Białowieskim Parku Narodowym [Dying hornbeams and their role for woodpeckers in the Białowieża National Park]. MS thesis, Siedlce University of Natural Sciences and Humanities, Siedlce.
- Myczko Ł, Benkman CW. 2011. Great spotted woodpeckers Dendrocopos major exert multiple forms of phenotypic selection on Scots pine Pinus sylvestris. Journal of Avian Biology 42:429–433. DOI: 10.1111/j.1600-048X.2011.05326.x.
- Nicolai B. 2011. Beobachtungen an einer Schmiede des Buntspechtes Dendrocopos major in Halberstadt. Ornithologische Jahresberichte des Museum Heineanum 29:77–84.
- Osiejuk T. 1991. Behawior żerowiskowy młodych dzięciołów dużych (Dendrocopos major) w lecie [Foraging behavior of young Great Spotted Woodpeckers (Dendrocopos major) in summer]. Notatki Ornitologiczne 32:27–36.
- Osiejuk T. 1994. Sexual dimorphism in foraging behaviour of the Great Spotted Woodpecker Dendrocopos major during winters with rich crops of Scotch pine cones. Ornis Fennica 71:144–150.
- Osiejuk T. 1998. Study on the intersexual differentiation of foraging niche in relation to abundance of winter food in Great Spotted Woodpecker Dendrocopos major. Acta Ornithologica 33:135–141.
- Pasinelli G. 2000. Sexual dimorphism and foraging niche partitioning in the Middle Spotted Woodpecker Dendrocopos medius. Ibis 142(4):635–644. DOI: 10.1111/j.1474-919X.2000.tb04463.x.
- Pawlaczyk P. 2009. Forest communities. In: Okołów C, Karaś M, Bołbot A, editors. Białowieża National Park. Know it - Understand it - Protect it. Białowieża: Białowieski Park Narodowy. pp. 37–58.
- Pechacek P. 2006. Foraging behavior of Eurasian Tree-toed Woodpeckers (Picoides tridactylus alpinus) in relation to sex and season in Germany. Auk 123(1):235–246. DOI: 10.1093/auk/123.1.235.
- Skoczylas R. 1961. Dynamika liczebności, rozmieszczenie pionowe i zachowanie terytorialne dzięcioła pstrego dużego (Dryobates maior L.) w borze sosnowym [Number changes, vertical distribution and territorial behaviour of Great Spotted Woodpecker (Dryobates major) in pine forest]. Ekologia Polska 14:229–243.
- Stański T. 2014. Porównanie miejsc żerowania i gniazdowania dzięcioła średniego Dendrocopos medius (L., 1758) i dzięcioła dużego Dendrocopos major (L., 1758) w pierwotnych grądach Białowieskiego Parku Narodowego [Comparison of foraging and nesting sites between the Middle Spotted Woodpecker Dendrocopos medius (L., 1758) and the Great Spotted Woodpecker Dendrocopos major (L., 1758) in a primeval lime-oak-hornbeam forest of Białowieża National Park]. Ph.D. thesis, Siedlce University of Natural Sciences and Humanities, Siedlce.
- Stenberg I, Hogstad O. 2004. Sexual dimorphism in relation to winter foraging in the white-backed woodpecker (Dendrocopos leucotos). Journal of Ornithology 145(4):321–326. DOI: 10.1007/s10336-004-0045-6.
- Tomiałojć L. 1991. Characteristics of old growth in the Bialowieza Forest, Poland. Natural Areas Journal 11(1):7–18.
- Wesołowski T, Czeszczewik D, Hebda G, Maziarz M, Mitrus C, Rowiński P. 2015a. 40 years of breeding bird community dynamics in a primeval temperate forest (Białowieża National Park, Poland). Acta Ornithologica 50:95–120. DOI: 10.3161/00016454AO2015.50.1.010.
- Wesołowski T, Rowiński P, Maziarz M. 2015b. Interannual variation in tree seed production in a primeval temperate forest: Does masting prevail? European Journal of Forest Research 134:99–112. DOI: 10.1007/s10342-014-0836-0.