
Abstract
Chondrichthyan egg cases are important elements for species-specific identification and also provide a valuable aid in determining a species spatial distribution, as well as for defining spawning areas. Considering the absence of a general key for the identification of the egg cases of the Mediterranean Chondrichthyes, this work aims to fill this gap by presenting a species-specific key based on morphological features of the egg case. The key was developed primarily analysing fresh egg cases dissected from the oviduct, egg cases collected from the seabed or found dried lying on the seashore, after species confirmation by DNA analysis. Original data were integrated with information scrutinized from literature. In order to improve species identification, a protocol for the standardized acquisition of morpho-biometric and meristic features is also provided as a pre-requisite for the appropriate use of the identification key. The total width and length included the horns, when they are not broken, are the parameters that best explain the assignment of the egg case to a specific species.
Introduction
Cartilaginous fishes are characterized by a K-type reproductive strategy with slow population growth and low resilience to biomass reduction, due to natural causes or over exploitation (Ricklef Citation1979; Hoenig & Gruber Citation1990; Ramírez-Amaro et al. Citation2020). The reproductive modes of cartilaginous fishes are very complex and articulated in different biological phases such as birth, growth, maturation and reproduction, with two main strategies: oviparous with development of the embryo external to the mother’s body and viviparous with development of the embryo internal to the mother’s body (sensu Hamlett Citation2005).
Oviparity, allows the production of one or more egg cases. In fact, the females of some species are able to retain up to 10 capsules in the oviducts before being deposited (Hamlett Citation2005). The embryo develops inside the egg case, and after consuming all of the yolk, emerges from the capsule with the same body shape as an adult (Hamlett Citation2005).
Some sharks (e.g. Scyliorhinidae, Heterodontidae, Orectolobidae), and all skates and chimaeras, are oviparous, producing a relatively thick capsule that protects the developing embryo from external factors while the embryo develops over several months to years, depending on the species (Hamlett Citation2005; Stevenson et al. Citation2007; Ebert & Stehmann Citation2013).
In the Mediterranean, even if the presence of some species is doubtful, there are at least 22 oviparous chondrichthyans (Serena Citation2005; FAO Citation2018a, Citation2018b). These include: four sharks with two species each in the genera Scyliorhinus and Galeus; 16 skates species: three Dipturus, one Rostroraja, four Leucoraja and eight Raja genera; one belonging to the genus Chimaera, one to the genus Hydrolagus. A further egg case belonging to the resurrected species Scyliorhinus duhamelii (Garman 1913) (Soares & De Carvalho Citation2019) has not been considered, since the species must be definitively confirmed for the Mediterranean (Serena et al. Citation2020).
Production of egg cases occurs throughout the year, with peaks in certain seasons. In the Mediterranean these peaks mainly occur in spring-summer (Serena et al. Citation2010). The deposition of egg cases is carried out in specific areas (egg case nursery areas) (Hoff Citation2016), towards which the cartilaginous fish migrate for hatching and where the females tend to concentrate in large groups (Springer Citation1967; Serena Citation2011). Being that spawning is strictly connected to ecological conditions, females usually utilize the same places every year (Castro Citation1993; Hoff Citation2009).
The taxonomic use of egg cases has been well documented and many authors have described the morphology of the egg cases by providing useful working tools (e.g. Clark Citation1922, Citation1926; Springer Citation1939; Ishiyama Citation1958; Cox Citation1963; Hitz Citation1964; Templeman Citation1982; Koob & Summers Citation1996; Howard Citation2002, Citation2017; Iglesias et al. Citation2002; Ebert Citation2005; Ebert et al. Citation2006, Citation2008; Treloar et al. Citation2006; Ebert & Davis Citation2007; Stevenson et al. Citation2007; Mabragaña et al. Citation2009, Citation2011; Concha et al. Citation2012; Ishihara et al. Citation2012; Maia et al. Citation2015; Bor Citation2016; Gordon et al. Citation2016; Porcu et al. Citation2017).
Due to the important role of Chondrichthyes related to biodiversity, conservation, and fishery ecology, here we develop a dichotomous key for taxonomic identification of egg cases based on their morphological, biometric and meristic features. This key is based on personal experiences combined with other descriptive keys of chondrichthyan egg cases (Iglesias et al. Citation2002; Treloar et al. Citation2006; Ebert & Davis Citation2007; Stevenson et al. Citation2007; Mabragaña et al. Citation2009; Ishihara et al. Citation2012; Gordon et al. Citation2016).
Material and methods
Data sources
Data and material collected over 35 years of Italian and International trawl surveys (GRUND and MEDITS respectively) conducted with the aim of evaluating fishery resources in the Mediterranean along with information gathered from research programs such as “Oceanografia e fondi Marini”, have provided numerous egg cases belonging to various species of sharks, skates and chimaeras (Bertrand et al. Citation2000; C.N.R Citation1979; Relini et al. Citation2010; Serena Citation2014; Follesa et al. Citation2019; Spedicato et al. Citation2019). Since it was not possible to collect egg cases for all chondrichthyan species living in the Mediterranean area, data were integrated from images provided by colleagues from outside the Mediterranean Sea.
Processing of the collected egg cases was carried out both on fresh (eggs extracted from female uteri) and on those beached or found on the sea bottom. For dry samples, it was needed a rehydration, which was achieved by immersing capsules in seawater for at least 30 minutes. Fresh or rehydrated capsules were stored in 70% alcohol. This allows species identification to be verified through genetic analysis of the egg cases or its contents afterwards. The mitochondrial Cytochrome Oxidase I (COI) gene, commonly used for DNA barcoding and widely applied in studies on Mediterranean elasmobranchs (Kousteni et al. Citation2015; Frodella et al. Citation2016; Cariani et al. Citation2017; Vella et al. Citation2017; Ferrari et al. Citation2018) also constitutes an efficient tool in the identification of their egg cases at species level (e.g. the case of Raja asterias [Catalano et al. Citation2021]), especially where the biological contents are better-preserved (Massi et al. Citation2018; Catalano Citation2020).
Morphology, biometry and meristics of the egg cases
In order to optimize the collection of information for each egg case, we utilized an updated scheme of morphological features of the egg cases for three chondrichthyan groups (sharks, skates and chimaeras). Using the features identified in the , a detailed guide to facilitate biometric or meristic measurements is given in order to make any future statistical analysis more reliable. Moreover, the egg case surface can show a furry surface texture constituted by rows of low rounded spines (Hoff Citation2009; Concha et al. Citation2010; Gordon et al. Citation2016). Due to the complexity of the analysis of these features, especially for beached egg cases, we have not included this feature in the identification key proposed here.
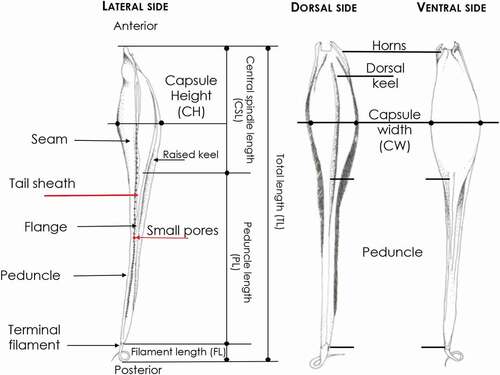
Figure 1. Morphology, terminology and biometry of the Mediterranean shark egg cases considered in the text.
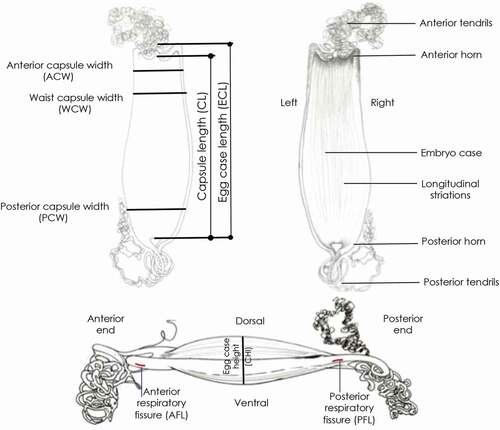
Figure 2. Morphology, terminology and biometry of the Mediterranean skate egg cases considered in the text (Partially from Stevenson et al. Citation2007).
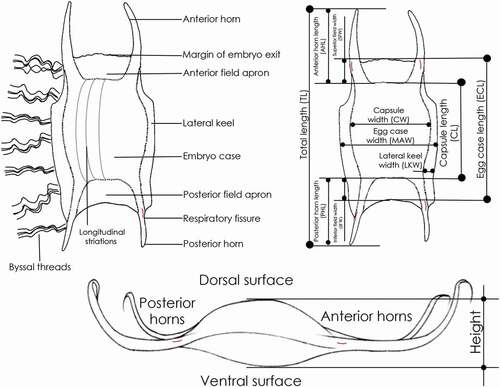
Figure 3. Morphology, terminology and biometry of the Mediterranean chimaera egg cases considered in the text.
Proposed identification key for the egg cases
In order to organize a useful identification key of the chondrichthyan egg cases, the detailed and diagnostic characters described below for the three groups, shown in , are utilized. The measurements are expressed in millimetres (mm) and detected using calliper. Another important diagnostic feature might is also be the structure of the capsule surface, the texture that characterizes it, the presence of filaments on the keel and on the horns, etc.
Sharks
It is possible to explain in detail the various components that describe the shape of the eggs belonging to the different species following the descriptions used by Ebert et al. (Citation2006), Flammang et al. (Citation2007) and Concha et al. (Citation2010) (). Therefore, the following measurements (in mm) of the egg cases were made: anterior border width (ABW), anterior capsule width (ACW), waist capsule width (WCW), posterior capsule width (PCW), posterior border width (PBW), anterior respiratory fissure length (AFL), egg case height (CHI), posterior respiratory fissure length (PFL), capsule length (CL) and egg case length (ECL).
Skates
It is possible to detail the morphological measurements of the egg case of skates by referring to Treloar et al. (Citation2006), Ebert & Davis (Citation2007) and Stevenson et al. (Citation2007) (), which suggest that some key measurements (mm) are to be taken, as used in this study: egg case length (ECL), anterior (proximal) horn length (AHL), posterior (distal) horn length (PHL), maximum egg case width (MAW), lateral keel width (LKW). In this study we suggest slightly different references: total length with horns (TL), making sure that the horns were not broken, capsule length without horns (CL), capsule width without keels (CW), superior field width (SFW) and inferior field width (IFW).
Chimaeras
Egg cases of Chimaeridae are not well documented, and the differences between capsules belonging to different species were not very evident. However, the number of respiratory pores on each lateral side of the egg case provides a good identification feature for this group ().
Also, for chimaera egg cases, as well as for sharks and skates, we suggest recording the main measurements (mm): total length (TL), central spindle length (CSL), peduncle length (PL), filament length (FL), capsule height (CH), capsule width (CW).
The dichotomous key proposed here, mainly takes into account the morphological features of the egg cases. In this sense, a protocol for the standardized acquisition of morpho-biometric and meristic features is provided as a pre-requisite for effective use of the identification key (). This protocol, with the inclusion of additional details, is provided according to Mancusi and Serena (Citation2017) who described the egg case morphology of the Mediterranean species and the analysis procedures used in the laboratory. In particular, with regard to the egg case of the new chimaera for the Mediterranean Hydrolagus mirabilis (Collett, 1904) (Hassan Citation2013; Farrag Citation2016), it seems to be extremely difficult to find it in the wild. Therefore, in order to provide a good dichotomous key, the features of the genus Hydrolagus have been utilized, while for the detailed description of morphological features the egg case of Chimaera monstrosa Linnaeus, 1758 was used.
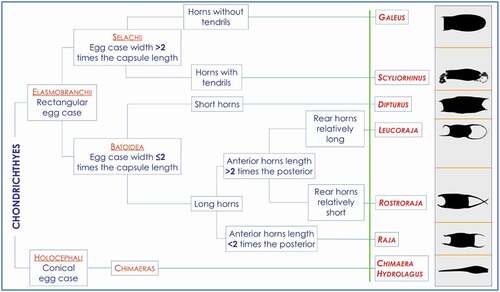
The use of taxonomy keys for determining the identification of egg cases to a particular species has been well documented and developed by Ishiyama (Citation1958), Hubbs & Ishiyama (Citation1968), Ishihara & Ishiyama (Citation1985), Ebert (Citation2005), Ebert & Davis (Citation2007) and Stevenson et al. (Citation2007). In this work, we try to adopt the same method to create a dichotomous key for identifying the egg cases of the species of sharks, skates and chimaeras that live in the Mediterranean basin. Furthermore, a simplified key for the correct identification at the genus level was also presented ().
Figure 4. Simplified key for the correct identification at the genus level. The picture it is not a cladogram.
Results
We were unable to collect the egg cases of all the Mediterranean species. Nevertheless, we were able to fill the gaps with assistance from colleagues who provided photographs of missing images from Atlantic Ocean specimens. In particular, these colleagues provided us with photographs of Leucoraja naevus (Müller & Henle, 1841), Leucoraja fullonica (Linnaeus, 1758) and Leucoraja circularis (Couch, 1838), the latter from the Clark (Citation1926) catalogue. The only egg case we were unable to obtain a sample from H. mirabilis, which was therefore left out of the key. Indeed, the research program allowed us to collect many egg cases of different chondrichthyans, taken directly from the female just before deposition. A total of 461 egg cases were collected and analyzed both from a morphological and statistical perspective (Mancusi & Serena Citation2017). In Annex I, for each individual species, the number of egg cases analysed are reported.
A preliminary multivariate analysis of biometrics of some chondrichthyan egg cases have suggested that their total width and length, in terms of absolute sizes, are the measures with the highest power for species identification. These two variables alone allow for a 96% separation of the correlation function of the species under study, obviously the horns must be unbroken (Mancusi & Serena Citation2017). Following the average value of the egg case width of some most representative species is shown ().
Table I. Main statistic of the egg-case width of nine skates: n = number of available specimens, avg = average width, sd = standard deviation, se = standard error, cl- and cl+ = 95% confidence limits, min and max = minimum and maximum observed values.
Morphology of the egg case
The egg case structure is typically oblong with a dorsal and a ventral side, the first usually convex correlated to the inner surface of the oviduct where it remains for a certain period.
Overall, egg case surface structure varies among species and is useful for identification at species level especially in the case of the skates. In fact, the egg case surface can show rows of low rounded spines creating a corduroy texture or rows of long multifid spines creating a furry surface texture (Hoff Citation2009; Concha et al. Citation2010; Gordon et al. Citation2016). These features, however, were not considered in the definition of the identification key proposed in this work.
Sharks
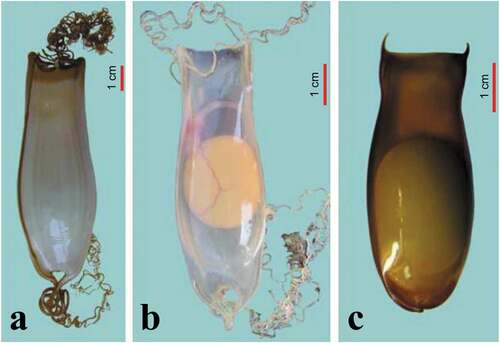
The egg cases, jar-shaped, are light yellow or dark brown when they are fresh and can be covered with 12–15 or more longitudinal striations that are partly evident and reinforce the structure. The egg case may be of differential thickness and can be relatively transparent, making the embryo visible with its yolk sac. The anterior edge of the egg case is straight, while the posterior edge is semi-circular with the horns tending to unite. The anterior and posterior ends of the egg case continue with small extensions called horns which in some species are considerably reduced or very long, with filaments curling up into tendrils, useful for anchoring the egg cases to a suitable substrate. At the ends of the egg cases, before the beginning of the horns, there are small openings on the dorsal and ventral side, respectively, useful for oxygenation (Ishiyama & Ishihara Citation1977; Flammang et al. Citation2007) ().
Figure 5. Three of the four egg cases from species of the two genera of Mediterranean sharks: a) Scyliorhinus stellaris; b) Scyliorhinus canicula; c) Galeus melastomus (by F. Serena).
Skates
The egg cases are usually rectangular shaped with a horny process at each corner, an anterior margin, through which the embryo, in most cases, comes out and a more or less flattened posterior margin. The dorsal and ventral surfaces are relatively convex. The longitudinal sides of the capsule are generally equipped with keels or fringed. The lateral keels in some species can extend anteriorly and posteriorly to the horns. The size of the horns varies from shortest, measuring just some millimetres, to very long, measuring several times the length of the capsule, in many cases with tapered horns that become filamentous, thread-like or flatten out towards the tips. In some species the egg cases may have, over their surfaces, fibrous bisso-like laminae, which allow them to adhere to the seabed. In any case, this does not preclude that deep currents can move them even if for short distances. The lateral margins of some capsules, depending on the species, may have fixing fibres. The oxygenation is aided by the fissures present at the ends, before the beginning of the horns, both on the dorsal and ventral sides (Ishiyama & Ishihara Citation1977; Flammang et al. Citation2007) ().
Figure 6. Egg cases of the four genera of Mediterranean skates: a) Dipturus; b) Leucoraja; c) Rostroraja; d) Raja. Photos by D. Massi (a); S. Iglesias (b); G. Morey (c); F. Serena (d).

Chimaeras
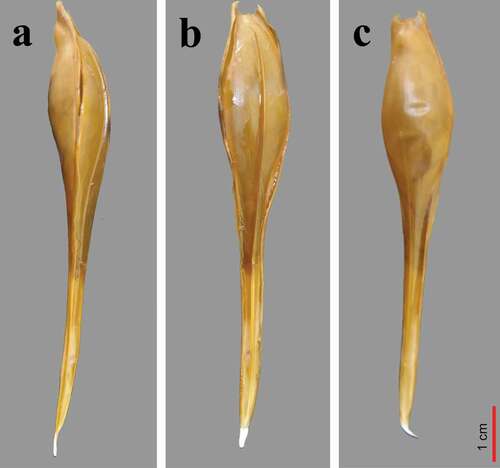
The egg cases are small, tapered and with little fringe. They have a tear drop-shape that thins in the back and becomes an elongated peduncle whose terminal filament often is inserted into the substrate. The dorsal surface has, in the middle part, a longitudinal keel. Along the edges of the central spindle and of the sheath or tail, there is a series of small pores which, after the deposition of the capsule, open to allow the oxygenation of the embryo (Dean Citation1906; Didier Citation1995) (). The dorsal side has an evident keel starts from the end of the peduncle to just before the opening placed in the proximal side of the egg case. The ventral side is devoid of keel and highlights the presence of thin rugae flowing towards the distal end of the peduncle. At the thinner end of the egg case, there is a short filament that usually gets fixed in the mud or on some substrate (Bini Citation1967; Armstrong Citation1996; Hamlett Citation2005).
Figure 7. Egg case of Chimaera monstrosa: a) Lateral side; b) Dorsal side; c) Ventral side. The chimaera egg case comes out of the genital opening with the dorsal side facing downwards. Then, the ventral side faces upwards in direct contact with the mother’s belly (by F. Serena).
Identification key for egg cases
Eight genera of Chondrichthyes occur in our study area divided into three distinct groups: sharks, skates and chimaeras. The main morphological features that allowed their distinction are summarized below: in sharks, there are egg cases completely devoid of tendrils (Galeus) unlike Scyliorhinus which instead has long ones on both ends of the egg case. The egg case of Dipturus has very short horns on both ends, unlike Leucoraja whose horns are always very long both at the anterior and posterior of the egg case. Rostroraja instead shows an intermediate situation with very long anterior horns and relatively short posterior ones. Raja is instead characterized by a regular, almost squared capsule whose anterior and posterior horn are almost equivalent but never very long. Finally, Chimaera egg case is a sort of cone with the upper part swollen and the lower part tapering ().
The identification key includes all of the known valid shark, skate and chimaera species in our study area (, Annex I). Only in two cases do we have non-specific egg cases regarding the species to which they belong, they are L. circularis and H. mirabilis. For the former, we have inserted in the key an egg case described by Clark (Citation1926) since it shows an undamaged egg case with complete horns. In the case of H. mirabilis, its egg cases were not available at all.
Table II. Identification keys to the chondrichthyans egg cases of the Mediterranean and Black Sea.
Discussion and conclusions
In order to better manage the chondrichthyan fish stocks, as well as to improve our knowledge of the conservation status of stocks and biodiversity, the recognition of certain zones as deposition, hatching and nursery areas is a priority. Being able to identify such nursery areas is one of the main requirements for the definition of sound actions required to conserve and enhance these Essential Fish Habitats (EFH) of certain species, intended as waters necessary for spawning, breeding, feeding or growth to maturity. The protection of such areas could be an excellent management measure in order to allow the renewal of elasmobranch populations (McMillan & Morse Citation1999; Serena et al. Citation2006). The observed presence of a large quantity of egg cases in a particular area, if repeated over time, is a way to determine whether an area is a spawning area that likely will subsequently concentrate there the newborn juveniles (Heupel et al. Citation2007; Hoff Citation2016; Rooper et al. Citation2019). Moreover, many skate species exhibit site fidelity, returning to the same nursery ground annually as reported for some species of Bathyraja spp. in the eastern Bering Sea (Hoff Citation2010) or for Raja asterias in the north Tyrrhenian Sea (Serena & Relini Citation2006). Therefore, the study of egg cases allows us not only to resolve taxonomic problems, but also to study the spatial distribution of habitat where the critical phases of species life cycle occur.
Thanks to their species-specific morphology, egg cases can be used to identify the species to which they belong. However, it is always difficult for the observer to attribute the parent species to the egg case, since little information on their morphology is often available, especially for some Mediterranean species. In these circumstances, we were forced to use egg cases of the same species collected in the Atlantic area (e.g. L. fullonica). In one case (e.g. L. circularis) even the original egg case of the individual studied is missing.
Indeed, regarding this last species very few images of fresh egg cases are available. However, some valid drawings or old images are usable from literature (Clark Citation1926; Mnasri et al. Citation2009). Despite this, we found it useful to produce a first dichotomous key for identifying the egg cases of the Mediterranean Chondrichthyes. A key that surely can be improved if we can collect and analyse more egg cases for each species, than those currently available.
Moreover, we are aware of the need to collect species-specific missing egg cases directly from the relevant pregnant females. Indeed, only in this way could we be completely sure of the real belonging of the egg cases to a certain species. In any case, genetic analysis provides useful help by directing choices in the best way. Distinguishing and assigning an egg case to a specific species represents a hard challenge. Nevertheless, considering two different methods simultaneously, such as genetic and morphologic approaches, this will help to achieve the goal of a better identification of the species to which the egg case belongs (Massi et al. Citation2018). For a genetic identification the mitochondrial COI gene can be used, while the main morphological features can be taken into account to compare the egg cases each other (Catalano Citation2020). In fact, the preliminary multivariate analysis of biometrics of the chondrichthyan egg cases suggested that the two main variables for species identification are total width and length. These variables even explain the 96% and certainly the width is the most reliable feature () (Mancusi & Serena Citation2017).
Finally, we must remember that the average values of the measurements mainly depend on the number of samples collected and analyzed, and/or the dimensions of the female. Indeed, the relationship between the total length of females and the total length of the egg cases has been demonstrated (Oddone et al. Citation2006).
Supplemental Material
Download PDF (25.8 MB)Acknowledgements
Our thanks go to all colleagues that provided us some pictures of the egg cases. In particular, Bárbara S. Pereira and Catarina Maia (IPMA - IP, Portugal), Gabriel Morey (CSIC/UIB, Spain), Peter H.F. Bor (Leiden, The Netherlands), Hajime Ishihara (W&I Ass. Japan), Francesca Gravino (Malta), Laura Castellani and Giovanni Raimondi (Genova and Livorno Aquarium, Italy), Dominique A. Didier (Millersville University, USA), Christian Capapé (Montpellier II University, France), and Alvaro J. Abella for their precious suggestions., Special thanks to Daniel Miller (Institute of Cancer Research, London, UK) for carefully reading the first version of the text in order to improve it from the English language point of view. We would like to thank to the anonymous referees for suggested us important amendments of some basic sentences for a better understanding of the text. Finally, major thanks go to Allison Wood for helping us to obtain a more understandable and better structured technically text.
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplementary material
Supplemental data for this article can be accessed here.
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
References
- Armstrong RH. 1996. Alaska’s fish. A guide to selected species. Alaska Northwest Books. pp. 94.
- Bertrand J, Gil De Sola L, Papakonstantinou C, Relini G, Souplet A. 2000. Contribution on the distribution of the Elasmobranchs in the Mediterranean Sea (from the MEDITS surveys). Biologia Marina Mediterranea 7:385–399.
- Bini G. 1967. Atlante dei pesci delle coste italiane. Vol. 1. Mondo Sommerso Editore. pp. 206.
- Bor P. 2016. Egg-capsules of sharks and skates. Available: HTTP://HOME.PLANET.NL/BOR00213/ENTER.HTML.
- C.N.R. 1979. Atti del Convegno Scientifico Nazionale Progetto Finalizzato Oceanografia e fondi marini. Vols. I, Il, III. Roma: Roma 5-6-7 Marzo 1979 - Consiglio Nazionale delle Ricerche. pp. 1–1493.
- Cariani A, Messinetti S, Ferrari A, Arculeo M, Bonello JJ, Bonnici L, Cannas R, Carbonara P, Cau A, Charilaou C, El Ouamari N, Fiorentino F, Follesa MC, Garofalo G, Golani D, Guarniero I, Hanner R, Hemida F, Kada O, Lo Brutto S, Mancusi C, Morey G, Schembri PJ, Serena F, Sion L, Stagioni M, Tursi A, Vrgoc N, Steinke D, Tinti F. 2017. Improving the conservation of Mediterranean Chondrichthyans: The ELASMOMED DNA Barcode Reference Library. PLoS One 12(1):e0170244. DOI: 10.1371/journal.pone.0170244.
- Castro JI1993. The shark nursery of Bulls Bay, South Carolina, with a review of the shark nurseries of the south-eastern coast of the United States. Environmental Biology of Fishes 38:37–48. DOI:10.1007/BF00842902.
- Catalano G.2020. Inferring the phylogeography and population structure of the Mediterranean endemic starry ray Raja asterias (Delaroche, 1809) with a special focus on egg cases geographical assignment. Thesis, Alma Mater Studiorum-University of Bologna. Scuola di Scienze – Campus di Ravenna Corso di Laurea Magistrale in Biologia Marina. Tesi di laurea in Struttura e Connettività delle Popolazioni Marine. Anno Accademico 2018/2019. pp. 86.
- Catalano G,Crobe V, Ferrari A, Baino R, Massi D, Titone A, Mancusi C, Serena F, Cannas R, Follesa MC, Hemida F, Manfredi C, Scarcella G, Sion L, Stagioni M, Tinti F, Cariani A. 2021. Strongly structured populations and reproductive habitat fragmentation increase the vulnerability of the Mediterranean starry ray Raja asterias(Elasmobranchii, Rajidae). Journal: Aquatic Conservation: Marine and Freshwater Ecosystems. Available: https://onlinelibrary.wiley.com/journal/10990755. In press.
- Clark RS. 1922. Rays and skates (Raiae). No. I - Egg capsules and young. Journal of the Marine Biological Association of the United Kingdom 12:577–643. DOI: 10.1017/S002531540000967X.
- Clark RS. 1926. Rays and skates. A revision of the European species. Fisheries, Scotland, Scientific Investigations 1:1–66.
- Concha F, Bustamante C, Oddone MC, Hernandez S, Lamilla J. 2010. Egg capsules of the dusky catshark Bythaelurus canescens (Carcharhiniformes, Scyliorhinidae) from the south-eastern Pacific Ocean. Journal of Fish Biology 77:963–971. DOI: 10.1111/j.1095-8649.2010.02736.x.
- Concha F, Oddone MC, Bustamante C, Morales N. 2012. Egg capsules of the yellownose skate Zearaja chilensis (Guichenot 1848) and the roughskin skate Dipturus trachyderma (Krefft and Stehmann 1974) (Rajiformes: Rajidae) from the south-eastern Pacific Ocean. Ichthyological Research 59:323–327. DOI:10.1007/s10228-012-0293-z.
- Cox KW. 1963. Egg cases of some elasmobranchs and a cyclostome from Californian waters. California Fish & Game 49:271–289.
- Dean B. 1906. Chimaeroid fishes and their development. Washington, DC: Carnegie Inst. Washington Publ. 32. pp. 156.
- Didier DA. 1995. Phylogenetic Systematics of Extant Chimaeroid Fishes (Holocephali, Chimaeroidei). Novitates. New York, 10024: American Museum of Natural History. Number 3119. pp. 86, 46 figures, 8 tables.
- Ebert DA. 2005. Reproductive biology of skates, Bathyraja (Ishiyama), along the eastern Bering Sea Continental Slope. Journal of Fish Biology 66:618–649. DOI:10.1111/j.0022-1112.2005.00628.x.
- Ebert DA, Compagno LJV, Cowley PD. 2006. Reproductive biology of catsharks (Chondrichthyes: Scyliorhinidae) from the west coast of southern Africa. ICES Journal of Marine Science 63:1053–1065. DOI:10.1016/j.icesjms.2006.04.016.
- Ebert DA, Compagno LJV, Cowley PD. 2008. Aspects on the reproductive biology of skates (Chondrichthyes: Rajiformes: Rajoidei) from southern African waters. ICES Journal of Marine Science 65:81–102. DOI:10.1093/icesjms/fsm169.
- Ebert DA, Davis CD. 2007. Description of skate egg cases (Chondrichthyes: Rajiformes: Rajoidei) from the eastern North Pacific. Zootaxa 1393:1–18.
- Ebert DA, Stehmann MFW. 2013. Sharks, batoids, and chimaeras of the North Atlantic. FAO Species Catalogue for Fishery Purposes. No. 7. Rome: FAO. pp. 523.
- FAO. 2018a. Species photographic plates. Mediterranean sharks, by Monica Barone, Fabrizio Serena and Mark Dimech. Rome, Italy.
- FAO. 2018b. Species photographic plates. Mediterranean skates, rays and chimaeras, by Monica Barone, Fabrizio Serena and Mark Dimech. Rome, Italy.
- Farrag MMS. 2016. Deep-sea ichthyofauna from Eastern Mediterranean Sea, Egypt: Update and new records. Egyptian Journal of Aquatic Research 42:479–489. DOI:10.1016/j.ejar.2016.12.005.
- Ferrari A, Tinti F, Bertucci Maresca V, Velonà A, Cannas R, Thasitis I, Oliveira Costa F, Follesa MC, Golani D, Hemida F, Helyar SJ, Mancusi C, Mulas A, Serena F, Sion L, Stagioni M, Cariani A. 2018. Natural history and molecular evolution of demersal Mediterranean sharks and skates inferred by comparative phylogeographic and demographic analyses. PeerJ 6:e5560. DOI: 10.7717/PEERJ.5560.
- Flammang BE, Ebert DA, Cailliet GM. 2007. Egg cases of the genus Apristurus (Chondrichthyes: Scyliorhinidae): Phylogenetic and ecological implications. Zoology 110:308–317. DOI:10.1016/j.zool.2007.03.001.
- Follesa MC, Marongiu MF, Zupa W, Bellodi A, Cau A, Cannas R, Colloca F, Djurovic M, Isajlovic I, Jadaud A, Manfredi C, Mulas A, Peristeraki P, Porcu C, Ramirez-Amaro S, Salmerón Jiménez F, Serena F, Sion L, Thasitis I, Cau A, Carbonara P. 2019. Spatial variability of Chondrichthyes in the northern Mediterranean. Scientia Marina 83S1. DOI: 10.3989/SCIMAR.04998.23A.
- Frodella N, Cannas R, Velonà A, Carbonara P, Farrell E, Fiorentino F, Follesa MC, Garofalo G, Hemida F, Mancusi C, Stagioni M, Ungaro N, Serena F, Tinti F, Cariani A. 2016. Population connectivity and phylogeography of the Mediterranean endemic skate Raja polystigma and evidence of its hybridization with the parapatric sibling. R.montagui. Marine Ecology Progress Series 554:99–113. DOI: 10.3354/MEPS11799.
- Gordon CA, Hood AR, Ellis JR. 2016. Descriptions and revised key to the egg- cases of the skates (Rajiformes: Rajidae) and catsharks (Carcharhiniformes: Scyliorhinidae) of the British Isles. Zootaxa 4150(3):255–280. DOI: 10.11646/ZOOTAXA.4150.3.2.
- Hamlett WC, editor. 2005. Reproductive biology and phylogeny of Chondrichthyes: Sharks, batoids and chimaeras. Enfield, New Hempshire 03748. United States of America: Scientific Publishers, INC, Post Office Box 699. pp. 562.
- Hassan M. 2013. Occurrence of large-eyed rabbitfish Hydrolagus mirabilis, Chimaeridae, in Syrian waters (Eastern Mediterranean). Marine Biodiversity Records 6, and 7. DOI: 10.1017/S175526721200111X.
- Heupel MR, Carlson KJ, Simpfendorfer CA. 2007. Shark nursery areas: Concepts, definition, characterization and assumptions. Marine Ecology Progress Series 337:287–297. DOI:10.3354/meps337287.
- Hitz CR. 1964. Observations on egg cases of the big skate (Raja binoculata (Girard, 1855)) found in Oregon coastal waters. Journal of the Fisheries Research Board of Canada 21:851–854. DOI:10.1139/f64-076.
- Hoenig JM, Gruber SH. 1990. Life-history patterns in the elasmobranchs: Implications for fisheries management. In: Elasmobranchs as living resources: Advances in the biology, ecology, systematics and the status of the fisheries. Edited by Pratt HL, Jr, SH Gruber and T Taniuchi. NOAA Technical Report NMFS 90:1–16.
- Hoff GR. 2009. Skate Bathyraja spp. egg predation in the eastern Bering Sea. Journal of Fish Biology 74:250–269. DOI:10.1111/j.1095-8649.2008.02137.x.
- Hoff GR. 2010. Identification of skate nursery habitat in the eastern Bering Sea. Marine Ecology Progress Series 403:243–254. DOI:10.3354/meps08424.
- Hoff GR. 2016. Identification of multiple nursery habitats of skates in the eastern Bering Sea. J Fish Biol 88:1746–1757.
- Howard MJ. 2002. Windows into elasmobranch embryonic development: A method to install a window on the egg case of the Big skate, Raja binoculata (Girard, 1855), before the completion of the first trimester. Drum and Croaker 33:26–30.
- Howard MJ. 2017. Chapter 44: Fecundity, egg case sizes, and neonate morphometrics of big skate Beringraja binoculata (Girard, 1855). In the Elasmobranch Husbandry Manual II: Recent advances in the care of sharks and their relatives. In Smith M, Warmolts D, Thoney D, Hueter R, Murray M, Ezcurra J, editors. Ohio Biological Survey, INC, Columbus. pp. 451–458, viii + 504.
- Hubbs C, Ishiyama R. 1968. Method for the taxonomic study and description of skates (Rajidae). Copeia 3:483–491.
- Iglésias SP, Du Buit M-H, Nakaya K. 2002. Egg capsules of deep-sea catsharks from eastern north Atlantic, with first descriptions of the capsule of Galeus murinus and Apristurus aphyodes (Chondrichthyes: Scyliorhinidae). Cybium 26:59–63.
- Ishihara H, Ishiyama R. 1985. Two new North Pacific skates (Rajidae) and a revised key to Bathyraja in the area. Japanese Journal of Ichthyology 32:143–179. DOI:10.1007/BF02938446.
- Ishihara H, Treloar M, Bor PHF, Senou H, Jeong CH. 2012. The Comparative Morphology of Skate Egg Capsules (Chondrichthyes: Elasmobranchii: Rajiformes). Bulletin of the Kanagawa Prefectural Museum (Natural Science) 41:17–33.
- Ishiyama R. 1958. Observations on the egg-capsules of the skates of the Family Rajidae, found in Japan and its adjacent waters. Bulletin of the Museum of Comparative Zoology Harvard College 118(1):1–24.
- Ishiyama R, Ishihara H. 1977. Five new species of skates in the genus Bathyraja from the western North Pacific, with reference to their interspecific relationships. Japanese Journal of Ichthyology 24(2):71–90.
- Koob TJ, Summers AP. 1996. On the hydrodynamic shape of little skate (Raja erinacea) egg capsules. Bulletin of the Mount Desert Island Biological Laboratory 35:108–111.
- Kousteni V, Kasapidis P, Kotoulas G, Megalofonou P. 2015. Strong population genetic structure and contrasting demographic histories for the small-spotted catshark (Scyliorhinus canicula) in the Mediterranean Sea. Heredity 114(3):333–343. DOI: 10.1038/hdy.2014.107.
- Mabragaña E, Figueroa DE, Scenna LB, Dìaz de Astarloa JM, Colonello JH, Delpiani G. 2011. Chondrichthyan egg cases from the southwest Atlantic. Journal of Fish Biology 79:1261–1290. DOI:10.1111/j.1095-8649.2011.03111.x.
- Mabragaña E, Figueroa DE, Scenna LB, Dìaz de Astarloa JM, Colonello JH, Massa AM. 2009. Clave de identificación de cápsulas de huevos de condrictios del mar argentino. INIDEP Informe Técnico 77:16.
- Maia C, Serra-Pereira B, Erzini K, Figueiredo I. 2015. How is the morphology of the oviductal gland and of the resulting egg capsule associated with the egg laying habitats of Rajidae species? Environmental Biology of Fishes 98:2037–2048. DOI: 10.1007/s10641-015-0425-1.
- Mancusi C, Serena F. 2017. Diagnosi morfologica delle capsule ovigere dei condroitti mediterranei. Metodiche di studio e chiavi di determinazione specifica. Seminario IAMC-CNR, Mazara del Vallo (TP). CNRSOLAR cod. 8512BC2017:75.
- Massi D, Cariani A, Mancusi C, Titone A, Ferrari A, Crobe V, Tinti F, Serena F. 2018. Biometry and molecular data of egg -cases of Raja asterias Delaroche, 1809 in the Strait of Sicily (Central Mediterranean). Biologia Marina Mediterranea 17(2):204.
- McMillan DG, Morse WW. 1999. Essential fish habitat source document: Spiny Dogfish, Squalus acanthias, Life History and Habitat Characteristics. NOAA Technical Memorandum NMFS-NE-150:1–19.
- Mnasri N, Boumaïza M, Capapé C. 2009. Morphological data, biological observations and occurrence of a rare skate, Leucoraja circularis (Chondrichthyes: Rajidae), off the northern coast of Tunisia (central Mediterranean). Pan-American Journal of Aquatic Sciences 4(1):70–78.
- Oddone MC, Mesa A, Ferreira de Amorim AF. 2006. The egg capsule of Rioraja agassizi (Müller & Henle) (Elasmobranchii, Rajidae), endemic to the SW Atlantic. Pan-American Journal of Aquatic Sciences 1(2):43–48.
- Porcu C, Marongiu MF, Bellodi A, Cannas R, Cau A, Melis R, Mulas A, Soldovilla G, Vacca L, Follesa MC. 2017. Morphological descriptions of the egg cases of skates (Rajidae) from the central-western Mediterranean, with notes on their distribution. Helgoland Marine Research 71:10. DOI: 10.1186/s10152-017-0490-2.
- Ramírez-Amaro S, Ordines F, Esteban A, Garcìa C, Guijarro B, Salmerón F, Terrasa B, Massutì E. 2020. The diversity of recent trends for chondrichthyans in the Mediterranean reflects fishing exploitation and a potential evolutionary pressure towards early maturation. Scientific Reports 10:547. DOI: 10.1038/S41598-019-56818-9.
- Relini G, Mannini A, De Ranieri S, Bitetto I, Follesa MC, Gancitano V, Manfredi C, Casciaro L, Sion L. 2010. Chondrichthyes caught during the medits surveys in Italian waters. Biologia Marina Mediterranea 17(1):186–204.
- Ricklef RE. 1979. Ecology. 2nd ed. New York: Chiron Press. pp. 966.
- Rooper CN, Hoff GR, Stevenson DE, Orr JW, Spies IB. 2019. Skate egg nursery habitat in the eastern Bering Sea: A predictive model. Marine Ecology Progress Series 609:163–178. DOI: 10.3354/meps12809.
- Serena F. 2005. Field identification guide to the sharks and rays of the Mediterranean and Black Sea. FAO Species Identification Guide for Fishery Purposes. Rome: FAO. pp. 97, 11 colour plates + egg cases.
- Serena F, Relini G. 2006. Use of Scientific Campaigns (Trawl Surveys) for the knowledge of the Sensitive Habitats. A review of the MEDITS, GRUND and APHIA data with special Attention to the Italian Seas. In: Basusta N, Keskin C, Serena F, Seret B, editors. The Proceedings of the Workshop on Mediterranean Cartilaginous Fish with Emphasis on Southern and Eastern Mediterranean. Vol. 23. Istanbul-Turkey: Turkish Marine Research Foundation. pp. 135–148.
- Serena F. 2011. Elasmobranchs. Chapter 2, Ecological aspects, Biodiversity: A great heritage. In: Cataudella S, Spagnolo M, editors. 2011 - The state of Italian marine fisheries and aquaculture. Rome (Italy): Ministero delle Politiche Agricole, Alimentari e Forestali (MiPAAF). pp. 156–158, 620.
- Serena F, editors. 2014. Lo status degli elasmobranchi dei mari italiani (Elasmostat). I Programma Nazionale triennale della pesca e dell’acquacoltura 2007-2009 (prorogato a tutto il 2012). Progetto di ricerca: “7 – Tematica A3”. Rapporto Finale 28 febbraio 2014:321. DOI: 10.13140/2.1.1721.4723.
- Serena F, Abella AJ, Barone M, Colloca F, Ferretti F, Fiorentino F, Bargnesi F, Moro S. 2020. Species diversity, taxonomy and distribution of Chondrichthyes in the Mediterranean and Black Sea. The European Zoological Journal 87(1):497–536. DOI: 10.1080/24750263.2020.1805518.
- Serena F, Abella JA, Baino R, Mancusi C, Voliani A, Relini G. 2006. Sensitive habitats for elasmobranch fishes in the Italian seas through the analysis of trawl surveys data. Report of the Mediterranean subgroup (SGMED 06-01) of the scientific, technical and economic committee for fisheries (STECF), sensitive and essential fish habitats in the Mediterranean Sea. Rome, 6–10 March 2006, pp. 358–378.
- Serena F, Mancusi C, Barone M. 2010. Field identification guide to the skates (Rajidae) of the Mediterranean Sea. Guidelines for data collection and analysis. Biologia Marina Mediterranea 17(2):204.
- Soares KDA, De Carvalho MR. 2019. The catshark genus Scyliorhinus (Chondrichthyes: Carcharhiniformes: Scyliorhinidae): Taxonomy, morphology and distribution. Zootaxa 4601(1):001–147. DOI: 10.11646/ZOOTAXA.4601.1.1.
- Spedicato MT, Massutí E, Mérigot B, Tserpes G, Jadaud A, Relini G. 2019. The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Scientia Marina 83S1:9–20. DOI: 10.3989/SCIMAR.04915.11X.
- Springer S. 1939. The egg case of the texas skate. Copeia 4:237. DOI:10.2307/1436897.
- Springer S. 1967. Social organization of shark populations. In GilbertPW, Mathewson,RF RallDPeditors, Sharks, Skates and Rays. Baltimore: Johns Hopkins Press. pp. 149–174.
- Stevenson DE, Orr JW, Hoff GR, McEachran JD. 2007. Field guide to sharks, skates, and ratfish of Alaska. Fairbanks, AK: Alaska Sea Grant. pp. 77.
- Templeman W. 1982. Development, occurrence and characteristics of egg capsules of the thorny skate, Raja radiata, in the northwest Atlantic. Journal of Northwest Atlantic Fishery Science 3:47–56. DOI:10.2960/J.v3.a4.
- Treloar MA, Laurenson LJB, Stevens JD. 2006. Descriptions of rajid egg cases from southeastern Australian waters. Zootaxa 1231:53–68. DOI:10.11646/zootaxa.1231.1.3.
- Vella A, Vella N, Schembri S. 2017. A molecular approach towards taxonomic identification of elasmobranch species from Maltese fisheries landings. Marine Genomics 36:17–23. DOI: 10.1016/J.MARGEN.2017.08.008.
Annex I.
List of the egg cases of each single species as identified in the dichotomous key (). In brackets, the number of egg cases analyzed in the period considered. Regarding Leucoraja circularis, it was preferred to use the old image of Clark (Citation1926) since it has shown unbroken horns. We were unable to find an original image of Hydrolagus mirabilis egg case. It seems that the egg capsule of this species has not yet been described.