
Abstract
The Gulf of Guinea is characterised by a complex hydrology and supports a high species diversity. A recent study has indicated that the area can be considered a refuge for rare taxa. Intensive sampling off the coast of Ghana (Gulf of Guinea) in 2012 on board the R/V F. Nansen resulted in the discovery of a large number of benthic invertebrates, including peracarid crustaceans. A preliminary investigation demonstrated a high diversity of the Cumacea, with 95% of the species being new to science. The most species-rich genus was Eocuma with seven species; six of them proved new to science and are described here. Additionally, the global distribution of Eocuma was analysed with the help of a phylogenetic tree based on morphological characters, Bayesian Binary Markov chain Monte Carlo biogeographic analysis and literature data. The radiation centres of the west African fauna are discussed.
Introduction
The genus Eocuma Marcusen, Citation1894 was erected to accommodate Eocuma hilgendrofi Marcusen, Citation1894 found off Japan. The genus was defined by a flattened carapace and the position of the first pereopods in ventral view, where the basal article comes to the midline (Marcusen Citation1894). The illustrations missing from the original description were later added by Zimmer (Citation1903), who noticed that the position of pereopods shifts during ontogenesis. However, because a similar pereopod position was observed in a few other genera (Calman Citation1904), this character was discarded as a diagnostic feature for Eocuma. As the lack of unique characters made the definition of Eocuma vague, Calman (Citation1904) transferred a few bodotriid species (i.e. Bodotria ferox (Fischer, Citation1872) and Cyclaspis sarsii (Kossmann, Citation1880)), with the carapace armed in lateral horns, to the genus. By that time, and after two new Eocuma species (E. affine and E. taprobanicum) had been discovered in the Gulf of Maanar (Indian Ocean), the genus contained five species (Fischer Citation1872; Kossmann Citation1880; Zimmer Citation1903; Calman Citation1904).
Somewhat later, Calman (Citation1907a, Citation1907b) described four species: one (E. longicorne) from the vicinity of Suez (Calman Citation1907a), two (E. latum and E. stelliferum) from the Gulf of Thailand (Calman Citation1907a), and one (E. dollfusi) from the Gulf of Biscay (Calman Citation1907b). Subsequently, Zimmer (Citation1914) and Fage (Citation1928, Citation1950) published descriptions of E. agrion from Australian waters and E. calmani and E. cadenati from off western Africa, respectively. Over the next few years, knowledge on the diversity and distribution of Eocuma was significantly expanded. New species were found off the Indian coasts (Kurian Citation1951, Citation1954) and off Japan (Gamô Citation1967). A survey of the South Africa fauna carried out by Day (Citation1978) resulted in discovering three new species (Day Citation1978). She supplemented the generic diagnosis of Eocuma by adding several characters, e.g. the pereonite 1 always covered by the carapace, the basis of the maxilliped 3 with a dorsodistal extension, the six-articled pereopod 2, and the one-articled endopod of the uropod (Day Citation1978). Following that new and restricted definition of Eocuma, seven species were described from the tropical Indian Ocean (Kurian & Radha Devi Citation1983; Mühlenhardt-Siegel Citation1996; Petrescu Citation1998, Citation2003; Patel et al. Citation2003; Corbera et al. Citation2005). In addition, Haye (Citation2007) synonymised Mossambicuma Day, Citation1978 with Eocuma, whereby the genus was complemented by M. elongatum Day, Citation1978 and M. victoriae Mühlenhardt-Siegel, Citation2003. Recently, Kim et al. (Citation2020) described a species (E. obriculatum) from the Korean waters.
At present, Eocuma is represented by 32 nominal species, which makes it the fourth most speciose bodotriid cumacean genus. However, despite the relatively long history of taxonomic studies on the genus, the knowledge of its diversity and distribution patterns is still far from complete.
The Gulf of Guinea is located off the west coast of equatorial Africa. It is characterised by a complex hydrology driven by several currents, i.e. the Guinea, Benguela and South Equatorial Counter Currents (Ukwe et al. Citation2006). In the northern part, environmental conditions are shaped by upwelling, while the southern part is strongly affected by nutrient input from land drainage and river runoff (mainly the Volta River) (Ukwe et al. Citation2006). This unique environmental setting is considered to support a high biodiversity in the region, recognised in the Marine Ecoregions of the World (MEOW) as a discrete ecoregion (Spalding et al. Citation2007).
Recent surveys of the Gulf of Guinea within the framework of the Oil for Development (OfD) Program (Annual Report Citation2019) under the aegis of the Food and Agriculture Organization of the United Nations (FAO) resulted in the discovery of a high biodiversity of benthic crustaceans, including peracarids (Jakiel et al. Citation2015; Stępień et al. Citation2021; Jóźwiak et al. Citation2022). For example, 95% of morphospecies in only one peracarid order, the Cumacea, collected off the Ghana coast proved new to science (Stępień et al. Citation2021). The genus Eocuma, represented by seven species, appeared to be the most diverse, and E. calmani Fage, Citation1928 was formally described.
This study is aimed at a taxonomic examination of Eocuma, an abundant and diverse cumacean genus identified in the benthic collection from the Gulf of Guinea. Six species new to science are described, and the worldwide distribution of Eocuma is analysed. Based on the current knowledge, we discuss the unique nature of the west African shallow-water fauna and its hypothetical dispersal routes.
Material and methods
Sampling
The material was collected with a Van Veen grab (0.1 m2) in October–November 2012 on board R/V Fridtjof Nansen. The sampling process was described by Pabis et al. (Citation2020). The samples were sieved through a 0.3 mm mesh size sieve and preserved in 4% formaldehyde solution. provides the location of the sampling sites.
Table I. Detailed information about samples with Eocuma species collected from the R/V Fridtjof Nansen.
Morphological studies and taxonomic description
Chemically sharpened tungsten needles were used for dissection of the individuals selected for detailed morphological analyses. The dissected appendages were placed on a microscopic slide in a drop of glycerine and closed with a coverslip. Drawings were prepared using a light microscope (Nikon Eclipse 50i) equipped with a camera lucida. Publication-quality illustrations were prepared using a digital tablet and Adobe Illustrator software (Coleman Citation2003). The morphological description follows Jakiel et al. (Citation2019), with “BL” standing for body length, “Nx” replacing “N times as long as” and “N L:W” replacing “N times as long as wide”. The type material was deposited at the University Museum of Bergen (Norway).
Phylogenetic analysis
A phylogenetic analysis was performed to investigate phylogenetic relationships between the Eocuma species. A matrix with 47 morphological characters and 33 Eocuma species including the newly described six was compiled. Thirty-two species were classified as an ingroup. Six species (E. affine Calman, Citation1904, E. agrion Zimmer, Citation1914; E. bacescui Petrescu, Citation2003; E. dimorphum Fage, Citation1928; E. hilgendorfi Marcusen, Citation1894; E. muradianae Petrescu, Citation1998) were excluded from the analysis because of their poor descriptions which included only the general appearance of the body and no details on either the pereopods or mouthparts. Both females and males were analysed; sexually dimorphic characters were considered separately for males and females. Based on the systematics and phylogeny of the family Bodotriidae (Haye Citation2007), Upselapsis caparti (Fage, Citation1951) was selected as an outgroup.
Four of the 47 characters were continuous and 43 were discrete. Discrete characters included 38 binary and five multistate traits. The characters were unordered and unweighted. The data matrix was prepared in Microsoft Excel and transformed, with the New Technology TNT (Goloboff et al. Citation2008), for use in the tree analysis.
Morphological characters
A heuristic search was performed in TNT v. 1.5 (Goloboff et al. Citation2008, Citation2016) using the traditional search algorithm appropriate for a smaller database (Goloboff et al. Citation2008), with 1000 replications and 10 trees held per replicate. The branch-swapping algorithm used was “tree bisection and reconnection” (TBR). The consensus tree was calculated using the strict consensus algorithm. The consistency index (CI), which is a way to measure homoplasy, was checked; it is calculated as a minimum number of changes divided by the number required on the tree, and ranged between 0 and 1 (CI = 1 means no homoplasy). The retention index (RI) shows how the tree fits to the character and how well synapomorphies explain the tree; it is calculated as (max steps − observed steps)/(max steps − min steps). RI ranges between 0 and 1; if RI = 1, the character fits perfectly. To measure the support of the clades obtained, the relative Bremer support (RBS) was calculated. RBS is based on the Bremer support, but it takes into account relative amounts of favourable and contradictory characters (Goloboff & Farris Citation2001). The RBS may be directly comparable with resampling techniques (Giribet Citation2003).
Biogeographical analysis
The distribution range of Eocuma was divided into six areas according to the realms proposed by Spalding et al. (Citation2007), i.e. (A) South Africa (SA), (B) Western Indo-Pacific (WIP), (C) Temperate North Pacific (TNP), (D) Tropical Atlantic (TA), (E) Temperate North Atlantic (TNA), and (F) Central Indo-Pacific (CIP). Should a species be recorded in two or more realms, only the type locality is considered (to avoid a possible misidentification). To analyse the possible ancestral range, the Bayesian Binary Markov chain Monte Carlo analysis (BBM) using the consensus phylogenetic tree was employed. BMM suggests possible ancestral ranges at each node and calculates probabilities at each ancestral range at the node (Joy et al. Citation2016). BMM was conducted with the Reconstruct Ancestral State in Phylogenies (RASP; Yu et al. Citation2015). The MCMC chains were run simultaneously for 50,000 generations. The state was sampled every 100 generations. The fixed Jukes–Cantor was used for BBM with the null root distribution. The maximum number of areas was six.
Results
Phylogenetic analysis
Heuristic searches resulted in four most parsimonious trees from which the consensus tree was developed () (length 152 steps, CI = 0.33, RI = 0.56). The RBS did not produce a significant value; the few branches that produced RBS > 0.5 are shown in .
Figure 1. The consensus tree developed for Eocuma Marcusen, Citation1894. Numbers above branches correspond to relative Bremer support; numbers below branches represent clade numbers.
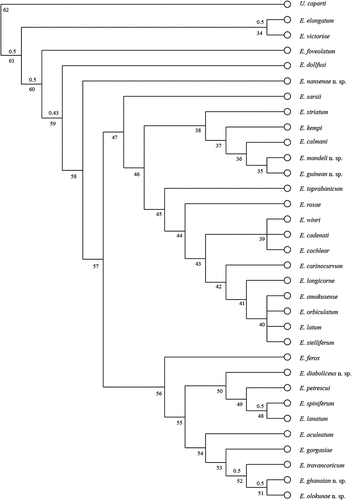
Clade 61 separates two species, E. victoriae and E. elongatum, from the remaining Eocuma species. The clade is supported by the following synapomorphies: maxilliped 3 ischium 3x merus, peropod 1 propodus longer than dactylus (at least 2x), the presence of relatively elongated siphon, and RBS equal to 0.5.
Clade 60 contains the other Eocuma species studied and was distinguished due to: carapace with 1.5 L:W or less, the presence of a projection on the basis of pereopod 1, and Maxilliped 3 ischium 2x merus or shorter. Three species (E. foveolatum, E. dollfusi and E. nansenae) failed to group with clade 60, but formed a cascade of branches (clades 60, 59 and 58). Eocuma foveolatum was distinguished due to the lack of horns and the proportion of articles of the second and third pereopods: pereopod 2 merus 1.2x carpus and pereopod 3 merus 0.8x carpus. Eocuma dollfusi was separated due to: pereopod 1 basis shorter than the length of ischium, merus, carpus, propodus and dactylus combined, and pereopod 1 dactylus shorter than propodus. Eocuma nansenae was separated due to a combination of the following characters: pseudorostrum situated on the horn level and a narrow ocular lobe.
The remaining Eocuma species formed clade 57, most species of which are characterised by the absence of a distinct medial carina. The clade is divided into two main lineages: clade 56 and 47. Clade 56 groups are E. aculeatum, E. diabolicus, E. ferox, E. gorgasiae, E. ghanaian, E. lanatum, E. olokunae, E. petrescui, E. spiniferum and E. travancoricum; their distinguishing characters include body not flattened (oval in cross section), carapace oval in dorsal view, and the presence of one pair of laterally directed horns.
Inside clade 56, E. ferox is distinct due to maxilliped 3 carpus which is 1.6x propodus; the remaining species form two sister clades: 50 and 54. Clade 54 groups E. aculeatum, E. gorgasiae, E. ghanaian, E. olokunae and E. travancoricum; these species are characterised by a rounded tip of horns which are relatively short (2 L:W or less). Clade 50 includes E. diabolicus, E. lanatum, E. petrescui and E. spiniferum, the species showing ocular lobes similar in length to or longer than half of the carapace as well as a lateral carina on it.
The second main lineage (clade 47) contains species (E. amakusense, E. cadenati, E. calmani, E. carinocurvum, E. cochlear, E. guinean, E. kempi, E. latum, E. longicorne, E. mandeli, E. orbiculatum, E. rosae, E. sarsii, E. stelliferum, E. striatum, E. taprobanicum and E. winri) with the carapace tapering towards the posterior and a distinct antennal notch. Although grouped with clade 47, E. sarsii is associated with a separate branch because of a relatively short uropod peduncle (0.25x rami), the presence of one pair of horns which are 2 L:W, and pseudorostrum situated at the level of the horns.
Clade 46 is subdivided into two sister groups: clades 38 and 45. The latter includes E. amakusense, E. cadenati, E. carinocurvum, E. cochlear, E. latum, E. longicorne, E. orbiculatum, E. rosae, E. stelliferum, E. taprobanicum and E. winri. Synapomorphy for the clade involves the first antennule article which is trapezoid in shape. Two species, E. rosae and E. taprobanicum, form the outermost branches within the clade. The former is separated due to the uropod peduncle being 0.4x rami, the latter showing maxilliped 3 carpus 2.5 times as long as wide and not meeting the pseudorostral plates. The remaining species are grouped inside clade 43 in which E. cadenati, E. cochlear and E. winri form a small unresolved group (clade 39), with articles (basis, ischium, merus) of pereopod3 being of a similar length. Clade 42 groups E. amakusense, E. carinocurvum, E. latum, E. longicorne, E. orbiculatum and E. stelliferum as distinct due to pleonites 5 and 6 being similar in length. In addition, E. amakusense, E. latum, E. orbiculatum and E. stelliferum form an unresolved clade 40.
Clade 38 groups E. calmani, E. guinean, E. kempi, E. mandeli and E. striatum, which form a cascade of branches and are clustered together because of their sharp horn tips.
Biogeographical analysis
Results of the BBM analysis show the genus to be dispersed in 19 areas (). The ancestral state is SA, with an 82% probability for clade 61, grouping E. victoriae and E. elongatum. The same ancestral state, with a probability of 88%, is proposed for clade 60 grouping the remaining Eocuma species. TNA is revealed as the ancestral state for clade 59 (E. dollfusi). A single dispersal, to the tropical part of the Atlantic (TA), is indicated for the next clade (58, with E. diaboliceus).
Figure 2. A graphical output from the BBM analysis (exported from RASP). Numbers in circles corresponds to the clade numbers from the phylogenetic consensus tree. A colour key to possible ancestral states at different nodes is given. SA – South Africa, WIP – Western Indo-Pacific, TNP – Temperate North Pacific, TA – Tropical Atlantic, TNA – Temperate North Atlantic, and CIP – Central Indo-Pacific.
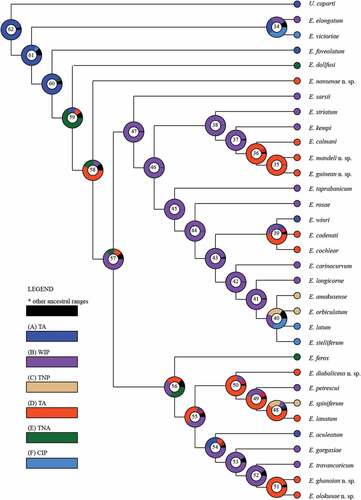
WIP is proposed as the ancestral state (with 63% probability) for clade 57 which divides Eocuma into two main lineages. Alternative ancestral states include TNP (13% probability) and TA (12%).
The ancestral state of the first main lineage (clade 56) remains unresolved, with three areas, TNP, WIP and TA, showing similar probabilities (32%, 29% and 26%, respectively). WIP is indicated as the ancestral state (98% probability) for the second main lineage (clade 47). Within clade 47, a single dispersion to TA (clades 36 and 39) as well as to CIP and TNP (clade 40) is observed.
Taxonomy
Genus Eocuma Marcusen, Citation1894
Diagnosis (after Kim et al. Citation2020)
Carapace oval or dorsally flattened, with or without lateral horn, with or without medial and/or dorso-lateral and/or lateral carinae. First pereonite invisible. Maxilliped 3 basis with dorsodistal prolongation. Pereopod 1 basis with or without dorsodistal prolongation. Pereopod 2 without ischium, with dactylus longer than propodus. Uropod peduncle shorter than pleotelson or rami. Uropod endopod one-articulated. Uropod exopod with two articles, first article shorter than second. Males with five pairs of pleopods.
Type species
Eocuma hilgendorfi Marcussen, Citation1894, by designation.
Species included
Eocuma aculeatum Day, Citation1978; E. affine Calman, Citation1904; E. agrion Zimmer, Citation1914; E. amakusense Gamô, Citation1967; E. bacescui Petrescu, Citation2003; E. cadenati Fage, Citation1950; E. calmani Fage, Citation1928; E. carinocurvum Corbera, Tirado and Martin, Citation2005; E. cochlear Le Loeuff and Intès, Citation1972; E. diaboliceus sp. nov.; E. dimorphum Fage, Citation1928; E. dollfusi Calman, 1907; E. elongatum (Day, Citation1978); E. ferox (Fischer, Citation1872); E. foveolatum Day, Citation1978; E. ghanaian sp. nov.; E. gorgasiae Mühlenhardt-Siegel, Citation1996; E. guinean sp. nov.; E. hilgendorfi Marcusen, Citation1894; E. kempi Kurian, Citation1954; E. lanatum Le Loeuff and Intès, Citation1972; E. latum Calman, 1907; E. longicorne Calman, 1907; E. mandeli sp. nov.; E. muradianae Petrescu, Citation1998; E. nansenae sp. nov.; E. olokunae sp. nov.; E. orbiculatum Kim, Lee and Kim, Citation2020; E. petrescui Patel, Haye and Kornfield, Citation2003; E. rosae Corbera and Galil, Citation2007; E. sarsii (Kossmann, Citation1880); E. spiniferum Gamô, Citation1976; E. stelliferum Calman, 1907; E. striatum Kurian and Radha Devi, Citation1983; E. taprobanicum Calman, Citation1904; E. travancoricum Kurian, Citation1951; E. winri Day, Citation1978; M. victoriae (Mühlenhardt-Siegel, Citation2003).
Eocuma calmani Fage, Citation1928
Material
One non-ovigerous female, ZMBN 149186 st.G1/100; one non-ovigerous female, one male, ZMBN 149187 st.G2/103; seven non-ovigerous females, ZMBN 149188 st.G3/100; two non-ovigerous females, one male, ZMBN 149189 st.G4/100; four non-ovigerous females, one male, ZMBN 149190 st.G6/103; 10 non-ovigerous females, three males, ZMBN 149191 st.G7/101; six non-ovigerous females, ZMBN 149192 st.G9/104.
Distribution
The Gulf of Guinea, off the coast of Ghana, Eastern Atlantic; depth about 100 m.
Eocuma diaboliceus sp. nov.
Registered under the ZooBank number urn:lsid:zoobank.org:act:256CC610-494E-4E9D-8F26-07EB41FC760A
Material
Holotype
Non-ovigerous female, ZMBN 149199 st. G6/51.
Paratype
One non-ovigerous female, dissected and drawn, ZMBN 149195 st. G1/50; two non-ovigerous females, one destroyed female, ZMBN 149195 st. G1/50; one non-ovigerous female, ZMBN 149196 st. G2/50; four non-ovigerous females, one brooding female, ZMBN 149197 st. G4/52; one non-ovigerous female, ZMBN 149198 st. G6/23; three non-ovigerous females, ZMBN 149199 st. G6/51.
Etymology
diaboliceus [Lat.] = devilish. The name refers to the long horns of the species.
Diagnosis
Carapace oval in dorsal view, with long (3.3 L:W) lateral horns directed almost 90° in relation to lateral margins. Medial carina weakly marked, lack of lateral carinae. Pleon 2x carapace. Pereopod-1 basis 0.6x remaining article combined, carpus 0.9x propodus, dactylus 0.4x propodus. Uropod exopod article2 with smooth margin (no crenulation).
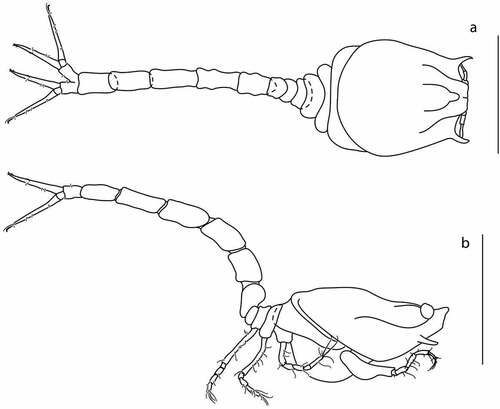
Description of female
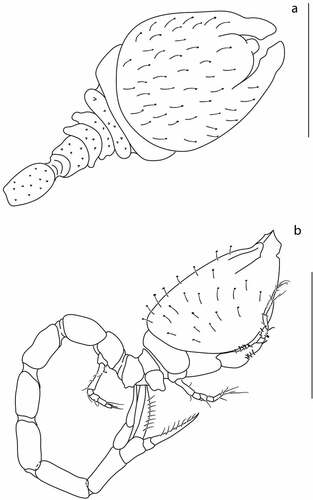
Body from holotype (ZMBN 149199), appendages from paratype (ZMBN 149195). BL = 2 mm. Ocular lobe oval. Carapace ()) 1.5 L:W, oval in dorsal view, with lateral horns (directed almost 90° in relation to lateral margin of carapace). Pseudorostral plates meeting in front of ocular lobe, triangular in dorsal view, with rounded apices. Antennal notch shallow. Medial carina weakly marked.
Figure 3. Eocuma diaboliceus sp. nov., non-ovigerous female, holotype (ZMBN 149199). (a) Body dorsal view; (b) body lateral view. Scale bar = 1 mm.
Pereon ()) 0.5x carapace, first pereonite invisible. Pleon 2x carapace.
Antennule ()) peduncle tri-articulated. Article 1 elongated, 3 L:W, 1.6x article 2, with two penicillate setae on lateral margin. Article 2 2.7 L:W, 0.6x article 3, with simple distal seta. Article 3 5 L:W, with a bunch of simple distal setae. Main flagellum two-articled, article 2 tipped with three aesthetascs. Accessory flagellum minute, tipped with two penicillate setae.
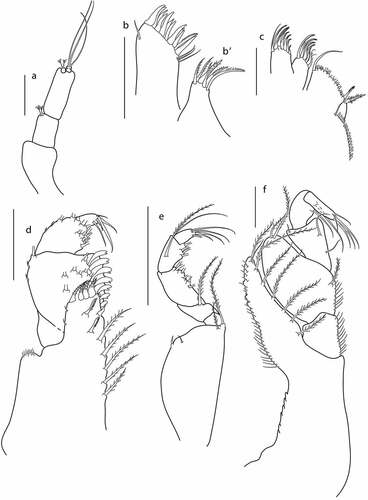
Figure 4. Eocuma diaboliceus sp. nov., non-ovigerous female, paratype (ZMBN 149195). a, Antennule; b, left mandible; c, right mandible; d, maxilla outer endite; d’, maxilla inner endite; e, maxilliped 1; f, maxilliped 2; g, maxilliped 3. Scale bars = 0.1 mm.
Antenna not observed.
Left mandible ()) with row of 10 plumose setae, incisor with four teeth, lacinia mobilis with three teeth.
Right mandible ()) with a row of 11 plumose setae, incisor with three teeth.
Maxilla outer endite ()) with 11 spines terminally, plumose subterminal seta on outer margin and bunch of simple subterminal setae on inner margin, and with simple setae along inner margin. Inner endite (’)) with four plumose terminal setae.
Maxillula not observed.
Maxilliped 1 ()) basis stout, with simple seta on medial surface, with bunch of setules on outer distal corner, and with three plumose setae on inner margin; endite with four plumose terminal setae (one with broad basis), with two plumose setae on medial surface and two hook setae on inner margin; merus 0.5x carpus, with no setae; carpus 1.4x propodus, with six flattened setae accompanied by simple seta along inner margin, with numerous setae on medial surface, and with plumose seta on outer distal corner; propodus 2.5x dactylus with four simple terminal setae and two subterminal setae; dactylus terminated with two simple setae.
Maxilliped 2 ()) basis as long as remaining articles combined, with two plumose setae on inner distal corner, and with short simple setae along outer margin; ischium short, with no setae; merus as long as carpus, with plumose setae on medial surface; carpus 0.8x propodus, with five plumose setae along inner margin; propodus 2.1x dactylus, with five plumose setae along inner margin; dactylus 0.4x propodus, with four simple terminal setae.
Maxilliped 3 ()) basis 0.6x remaining articles combined, with crenulation along inner margin in distal half, distal prolongation reaching articulation of ischium and merus, with a row of five plumose setae along inner margin, a row of six simple setae along outer margin, and with three simple distal setae; ischium 0.4x merus, with four plumose setae along inner margin; merus 1.4 L:W, as long as carpus, with four plumose setae along inner margin; carpus 2.0 L:W, as long as propodus, with no setae; propodus 2.1 L:W, as long as dactylus, with two plumose setae at medial surface; dactylus with four sharp teeth along inner margin, minute seta on outer margin in proximal half, and with five simple terminal setae. Exopod as long as basis.
Pereopod 1 ()) basis 0.6x remaining article combined, proximal part wider than terminal part, with a row of numerous setae on ventral margin, and a row of teeth along dorsal margin, and with plumose seta on distal corner; ischium 0.4x merus, with no setae; merus 0.6x carpus, with seven simple setae; carpus 0.9x propodus, with three simple setae at medial surface; propodus 2.1x dactylus, with 14 simple setae on whole surface; dactylus 0.4x propodus, with three simple terminal setae and three simple subterminal setae. Exopod equal to basis.
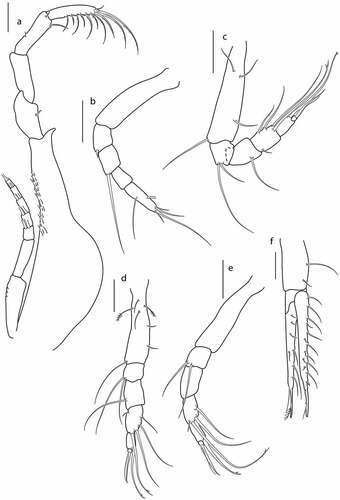
Figure 5. Eocuma diaboliceus sp. nov., non-ovigerous female, paratype (ZMBN 149195). a, Pereopod 1; b, pereopod 2; c, pereopod 3; d, pereopod 4; e, pereopod 5; f, uropod. Scale bars = 0.1 mm.
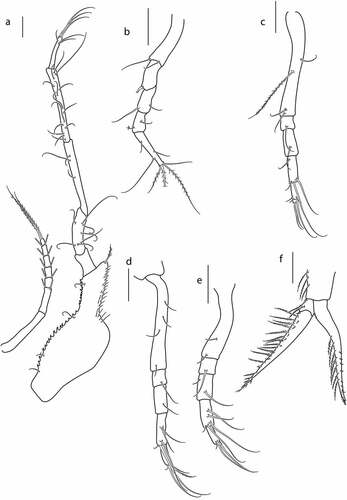
Pereopod 2 ()) basis 0.4x remaining articles combined, with two distal setae; ischium fused with basis; merus 1.2x carpus, with spine on distal corner; carpus 1.2x propodus, with simple setae; propodus 0.5x dactylus, with simple and penicillate setae; dactylus 3 L:W, with three simple setae laterally, and with four plumose terminal setae.
Pereopod 3 ()) basis 1.1x remaining articles combined, with simple seta and plumose seta on lateral margin, and with two simple distal setae; ischium 0.6x merus, with simple distal seta; merus as long as carpus, with simple setae at medial surface; carpus 1.5x propodus, with simple seta on lateral margin, and with two simple setae (one long, one short), and one bifurcated distal seta; propodus 3.4x dactylus, with strong bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Pereopod 4 ()) basis as long as remaining articles combined, with three simple setae in proximal half, and with simple seta subdistally; ischium 0.6x merus, with simple distal seta; merus 0.8x carpus, with simple seta on medial surface; carpus 1.5x propodus, with two simple setae on medial surface, and with two simple setae (one long, one short), and one bifurcated distal seta; propodus 3.5x dactylus, with bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Pereopod 5 ()) basis 0.7x remaining articles combined, with simple seta on medial surface, and simple distal seta; ischium 0.6x merus, with simple distal seta; merus 0.8x carpus, with two simple setae; carpus 1.6x propodus, with two simple setae on medial surface, and with two simple setae (one long, one short), and one bifurcated distal seta; propodus 3.5x dactylus, with bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Uropod ()) peduncle about 0.4x pleotelson, with three plumose setae along lateral margin. Endopod one-articulated, 3.2x peduncle, with row of seven plumose setae along inner margin, and row of seven short simple setae along outer margin, terminated with setulose seta. Exopod two-articulated, subequal to endopod; article 1 with two plumose setae on outer margin; article 2 with two rows of numerous setae on outer margin: first row of long plumose setae, second row of short simple setae, terminated with two stout setulose setae.
Distribution
Gulf of Guinea, off the coast of Ghana, Eastern Atlantic; depth 20–50 m.
Remarks
Long, forward-directed lateral horns characteristic for E. diaboliceus are also observed in E. aculeatum, E. gorgasiae, E. lanatum and E. travancoricum. However, the almost straight direction of the horns (right angle to lateral margins) in the new E. diaboliceus is unique among all the Eocuma with lateral horns. Moreover, E. diaboliceus is the only species (among those listed above) that has a serrated lateral margin of pereopod 1 basis. Moreover, the new species lacks ornamentation on carapace and pereonites, in contrast to E. aculeatum and E. travancoricum which have tubercles and setae on the tegumental surface. Much longer and slenderer horns differentiate E. diaboliceus from E. gorgasiae, and the smooth margin of uropod rami characteristic for the new species distinguishes it from E. lanatum with crenulation on uropod exopod.
Eocuma ghanaian sp. nov.
Registered under the ZooBank number urn:lsid:zoobank.org:act:F50E63F3-1B65-48B7-88E6-42BA9B613723
Material. Holotype
Brooding female, ZMBN 149193 st. G6/23.
Paratype
One non-ovigerous female, dissected and illustrated, ZMBN 149194 st. G6/23; one non-ovigerous female, one destroyed female and one juvenile male, ZMBN 149194 st. G6/23.
Etymology
Named after the type locality: the Ghanaian coast.
Diagnosis
Carapace oval in dorsal view, covered with minute setae, without medial and dorsolateral carinae, without horns. Pleon 1.7x carapace. Pereopod 1 basis 0.8x remaining articles combined, carpus similar to propodus, dactylus 0.6x propodus. Uropod exopod article 2 with crenulated margin.
Description of female
Body from holotype (ZMBN 149193), appendages from paratypes (ZMBN 149194). BL = 4.3 mm. Ocular lobe oval. Carapace ()) 1.3 L:W, oval in dorsal view, surface covered with minute setae, without lateral horns. Pseudorostral plates rounded, not meeting. Antennal notch tiny. Medial carina absent, dorsolateral carinae absent.
Figure 6. Eocuma ghanaian sp. nov., brooding female, holotype (ZMBN 149193). a, Body dorsal view; b, body lateral view. Scale bars = 1 mm.
Pereon ()) 0.6x carapace, first pereonite invisible, pereonites 2–5 covered with small tubercles. Pleon 1.7x carapace.
Antennule ()) peduncle tri-articulated. Article 1 widest, with parallel margins, and with two simple setae along lateral margin. Article 2 2.2 L:W, 0.7x article 3, with seta on inner and seta on outer margin. Article 3 3 L:W, with simple and penicillate distal setae. Main flagellum two-articulated, article 2 tipped with two long simple setae, one aesthetasc and one penicillate seta. Accessory flagellum minute, tipped with two penicillate and simple setae.
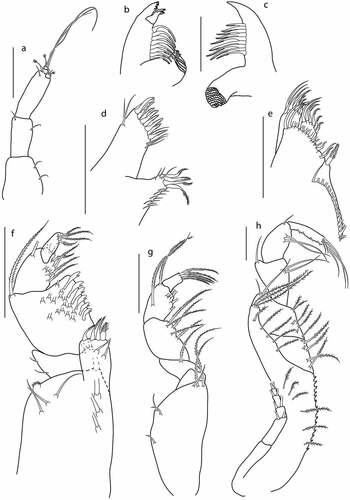
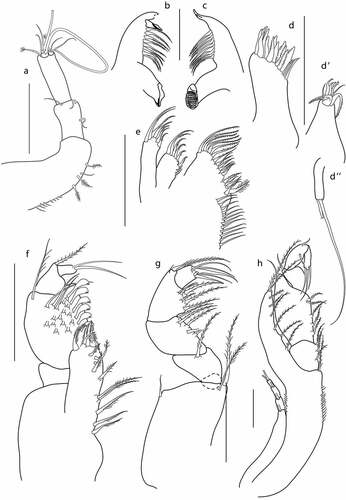
Figure 7. Eocuma ghanaian sp. nov., non-ovigerous female, paratype (ZMBN 149194). a, Antennule; b, left mandible; c, right mandible; d, maxilla; e, maxillula; f, maxilliped 1; g, maxilliped 2; h, maxilliped 3. Scale bars = 0.1 mm.
Antenna not observed.
Left mandible ()) with a row of eight plumose setae, incisor with four teeth, lacinia mobilis with three teeth.
Right mandible ()) with a row of nine plumose setae, incisor with blunt tooth.
Maxilla ()) outer endite with 11 spines terminally, two simple subterminal setae on outer margin, and three simple subterminal setae on inner margin. Inner endite with four stout, plumose terminal setae and numerous thin setae along lateral margin.
Maxillula ()) broad endite with row of 16 subterminal setae, and with a row of 16 simple setae and one plumose terminal seta. Outer endite with five plumose terminal setae and two simple subterminal setae. Inner endite with six plumose terminal setae.
Maxilliped1 ()) basis stout, with six simple setae on medial surface, a bunch of setules on outer distal corner, and with seta on inner distal corner; endite with four stout plumose terminal setae and with two setae on medial surface; merus 0.3x carpus, with two setae on inner distal corner and with two setae along outer margin; carpus 1.2x propodus, with six flattened setae along inner margin, with numerous setae on medial surface, and with long plumose setae on outer distal corner; propodus 1.5x dactylus, with nine plumose setae along inner margin, and with seta on outer distal corner; dactylus 0.6x propodus, with two terminal setae and with three subterminal plumose setae.
Maxilliped2 ()) basis 0.8x remaining articles combined, with one simple and two distal plumose setae and with three simple setae on outer margin; ischium short, with no setae; merus 0.4x carpus, with plumose distal setae; carpus 1.3x propodus, with five plumose setae along inner margin; propodus 2x dactylus, with six setae along inner margin, with plumose setae on medial surface, and with plumose seta on outer distal corner; dactylus with five plumose terminal setae.
Maxilliped3 ()) basis stout, 0.5x remaining articles combined, with row of teeth along inner margin, and with row of six plumose setae on medial surface, distal prolongation reaching articulation of ischium and merus, with a row of eight plumose setae along distal and inner margin; ischium subequal to merus, with six plumose setae along inner margin; merus 1.2 L:W, 0.8x carpus, with six plumose setae along inner margin and with two simple setae on outer distal corner; carpus 1.4 L:W, similar in length to propodus, with two simple setae at midlength of inner margin, and with simple seta on outer distal corner; propodus 1.5 L:W, 1.2x dactylus, with two simple setae at midlength of inner margin, and with simple seta on outer distal corner; dactylus 0.8x propodus, with three simple terminal setae and two subterminal setae and with row of four short setae along inner margin. Exopod shorter than basis.
Pereopod1 ()) basis 0.8x remaining articles combined, proximal part wider than distal, with nine plumose setae medially, and with plumose seta on distal corner; ischium 0.4x merus, with short simple seta laterally; merus 0.8x carpus, with five plumose setae laterally; carpus 0.8x propodus, with three simple setae laterally; propodus 1.5x dactylus, with three short simple setae laterally, and with three long setae distally; dactylus 0.6x propodus, with three simple terminal setae, and with three short simple subterminal setae. Exopod subequal to basis.
Figure 8. Eocuma ghanaian sp. nov., non-ovigerous female, paratype (ZMBN 149194). a, Pereopod 1; b, pereopod 2; c, pereopod 3; d, pereopod 4; e, pereopod 5; f, uropod. Scale bars = 0.1 mm.
Pereopod2 ()) basis 0.6x remaining articles combined, with two simple setae; ischium fused with basis; merus 1.2x carpus, with spine on distal corner; carpus subequal to propodus, with two simple distal setae; propodus 0.6x dactyls, with no setae; dactylus 3.2 L:W 1.7x propodus, with four terminal setae and two simple setae on lateral margins.
Pereopod3 ()) basis 0.5x remaining articles combined, with simple distal seta; ischium 0.6x merus, with simple distal seta; merus subequal to carpus, with simple setae laterally, carpus 1.5x propodus, with simple setae laterally, and with two setae distally: one bifurcate, and one simple; propodus 3x dactylus, with strong bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Pereopod4 ()) basis 0.8x remaining articles combined, with five simple setae and one penicillate seta laterally; ischium 0.7x merus, with simple setae subdistally; merus 0.8x carpus, with simple seta laterally; carpus 1.5x propodus, with penicillate seta laterally on proximal half, and with one simple seta and one bifurcated seta on distal corner; propodus 3.2x dactylus, with strong distal seta; dactylus terminated with one thin seta and one strong seta.
Pereopod5 ()) basis 0.5x remaining articles combined, with five simple setae and one penicillate seta laterally; ischium 0.7x merus, with simple setae subdistally; merus subequal to carpus, with two simple setae laterally; carpus subequal to propodus, with simple seta laterally on proximal half, and with two setae (one broken) on distal corner; propodus 3.3x dactylus, with strong bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Uropod ()) peduncle 0.3x pleotelson. Endopod one-articulated, 4.5x peduncle, with a row of five penicillate setae along outer margin, and with a row of eight setae along inner margin, terminated with stout, setulose seta. Exopod two-articulated, subequal to endopod, second article with three setae on outer margin, and crenulation on inner margin, terminated with two stout setulose setae.
Distribution
The Gulf of Guinea, off the coast of Ghana, Eastern Atlantic; depth about 20 m.
Remarks
Eocuma ghanaian sp. nov., E. elongatum, E. foveolatum, E. obriculatum and E. mandeli sp. nov. share the absence of lateral horns on the carapace. Four of these species, E. ghanaian, E. foveolatum, E. obriculatum and E. elongatum, show basis of pereopod1 shorter than total length of the remaining articles. Moreover, two species, E. ghanaian and E. foveolatum, show the carapace to be of a similar shape, tapering in the anterior part. However, E. ghanaian can be distinguished from the other four Eocuma by the lack of small holes on the surface of the body. The carapace is covered with minute setae and the pereonites are covered with small tubercles. Furthermore, the dorsolateral carinae, obvious in E. obriculatum and marked by a shallow indentation or medial depression in E. mandeli, E. elongatum and E. faveolatum, are apparently absent in E. ghanaian. Finally, uropod exopod article2, crenulated in E. ghanaian, is simple in other species, or only setae are present, as in E. obriculatum and E. mandeli, or setae with spines, as in E. faveolatum.
Eocuma guinean sp. nov.
Registered under the ZooBank number urn:lsid:zoobank.org:pub:A4A175E4-91A9-41DF-B8D0-01C39CBDFCB6
Material
Holotype
Non-ovigerous female, ZMBN 149215 st. G5/26.
Paratype
One non-ovigerous female, dissected and drawn, ZMBN 149214 st. G4/52; one non-ovigerous female and two juvenile males, ZMBN 149216 st. G6/23; one adult male, body illustrated, ZMBN 149217 st. G9/25; one non-ovigerous female, ZMBN 149217 st. G9/25.
Etymology
Named after type locality: the Gulf of Guinea.
Diagnosis
Carapace dorsoventrally flattened, with lateral carinae, and with two pairs of pointed horns, tip of anterolateral pair of horns at the level of pseudorostrum tip. Pleon 1.2x carapace. Pereopod 1 basis 0.5x remaining articles combined, carpus similar to propodus, dactylus 0.8x propodus. Uropod exopod article 2 with smooth margin (no crenulation).
Description of female
Body from holotype (ZMBN 149215), appendages from paratypes (ZMBN 149214). BL = 6.1 mm. Ocular lobe rounded. Carapace ()) 1.1 L:W, strongly flattened, with two pairs of horns, horns short (about as long as wide). Pseudorostral plates rounded. Antennal notch absent. Lateral carinae nearly reach the border of carapace.
Figure 9. Eocuma guinean sp. nov., non-ovigerous female, holotype (a, b: ZMBN 149215), and adult male, allotype (c, d: ZMBN 149217). a, Body dorsal view, b, Body lateral view, c, Body dorsal view, d, Body lateral view. Scale bar = 1 mm.
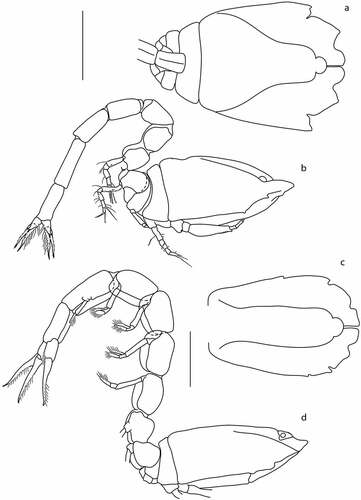
Pereon ()) 0.7 × carapace, first pereonite invisible. Pleon 1.2x carapace.
Antennule ()) peduncle tri-articulated. Article 1 trapezoidal, 3.1 L:W, 2.7x article 2, with two penicillate distal setae, and with numerous setae on lateral surface; article 2 0.6x article 3, with two penicillate distal setae; article 3 with short simple seta in proximal half, and with two penicillate setae and simple distal seta. Main flagellum two-articulated, article 2 tipped with aesthetasc and two simple setae. Accessory flagellum minute, tipped with two penicillate setae.
Figure 10. Eocuma guinean sp. nov., non-ovigerous female, paratype (ZMBN 149214). a, Antennule; b, left mandible; c, right mandible; d, maxilla outer endite; D’, maxilla inner endite; e, maxillula; f, maxilliped 1; g, maxilliped 2; h, maxilliped 3. Scale bars = 0.1 mm.
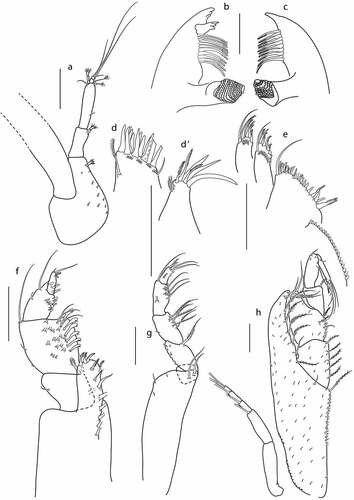
Antenna not observed.
Left mandible ()) with a row of 11 plumose setae, incisor with three teeth, lacinia mobilis with three teeth.
Right mandible ()) with a row of 10 plumose setae, incisor with blunt tooth.
Maxilla outer endite ()) with 12 spines terminally, four bunches of simple subterminal setae and plumose seta on inner margin. Inner endite (’)) with three plumose setae and three stout simple terminal setae, and four simple setae on outer margin.
Maxillula ()) broad endite with a row of 20 subterminal setae, and with a row of 21 simple setae and two plumose terminal setae. Outer endite with five plumose terminal setae, and two simple subterminal setae. Inner endite with four plumose and one simple terminal seta.
Maxilliped 1 ()) basis stout, endite with five plumose setae along distal margin, and with a row of seven setae along inner margin; merus 0.7x carpus, with two plumose setae on inner distal corner, and two simple setae on both lateral margins; carpus 1.3x propodus, with six flattened setae along inner margin, with numerous setae on medial surface, and with long plumose seta on outer distal corner; propodus 2x dactylus, with two rows of simple setae along inner margin, and two simple setae on inner margin; dactylus with two terminal setae and two subterminal short setae.
Maxilliped 2 ()) basis as long as remaining articles combined, with minute setae on outer distal corner, and with two long simple setae on inner distal corner; ischium short, with no setae; merus as long as carpus, with plumose seta on inner distal corner, and numerous short setae along outer margin; carpus as long as propodus, with five plumose setae along inner margin, and short seta on outer margin; propodus 1.9x dactylus, with two rows of eight setae along inner margin, with seta on medial surface, and with long, simple seta on outer distal corner; dactylus with four simple terminal setae, with seta at medial surface, and with minute setae along inner margin.
Maxilliped 3 ()) basis stout, 0.7x remaining articles combined, with long plumose seta on inner distal corner, and with simple distal seta, distal prolongation reaching articulation of merus and carpus, with a row of plumose setae along inner margin, whole surface of basis covered with minute setae; ischium as long as merus, with five plumose setae along inner margin, and with minute setae on whole surface; merus 1.3 L:W, 1.1x carpus, with three plumose setae along inner margin, and with minute setae on whole surface; carpus 1.3 L:W, as long as propodus, with plumose seta on inner distal corner, and simple seta on outer distal corner; propodus 2.3 L:W, as long as dactylus, with simple seta on inner margin, simple seta on outer distal corner, and with minute setae on medial surface distally; dactylus with two simple terminal setae, and two subterminal simple setae. Exopod shorter than basis.
Pereopod 1 ()) basis 0.5x remaining articles combined, with small teeth and simple setae on distal corner, and with minute setae on whole surface; ischium 0.3x merus, with no setae; merus 0.3x carpus, with small setae on medial surface; carpus as long as propodus, with six lateral setae and one distal seta; propodus 1.2x dactylus, with three short setae on medial surface, and with three long distal setae; dactylus with three simple setae laterally, with four setae (three simple, one plumose) terminally, and with two simple subterminal setae. Exopod shorter than basis.
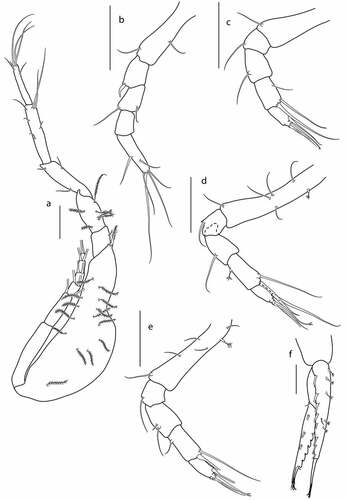
Figure 11. Eocuma guinean sp. nov., non-ovigerous female, paratype (ZMBN 149214). a, Pereopod 1; b, pereopod 2; c, pereopod 3; d, pereopod 4; e, pereopod 5; f, uropod. Scale bars = 0.1 mm.
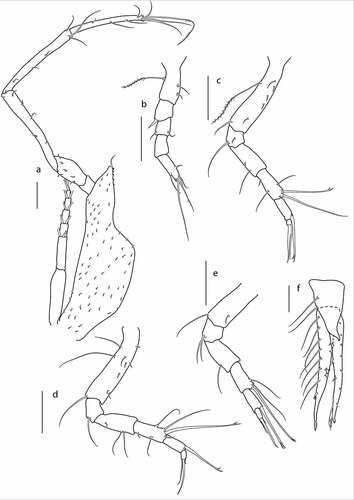
Pereopod 2 ()) basis 0.5x remining articles combined, with plumose setae and two simple lateral setae; ischium fused with basis; merus 1.2x carpus, with spine on distal corner; carpus 0.7x propodus, with two simple distal setae; propodus 0.7x dactylus, with no setae; dactylus 3.1 L:W, with two lateral setae laterally and four terminal setae.
Pereopod 3 ()) basis 0.8x remaining articles combined, with long plumose setae and five short lateral setae, and with simple setae on distal corner; ischium 0.4x merus, with short seta on medial surface, and with long, simple seta on distal corner; merus 0.8x carpus, with two simple lateral setae, carpus 1.6x propodus, with three simple lateral setae, and with two setae (one simple, one bifurcated) on distal corner; propodus 2.7x dactylus, with strong bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Pereopod 4 ()) basis 0.7x remaining articles combined, with two long simple setae, and seven short simple setae on lateral margin, and with long simple seta on distal corner; ischium 0.4x merus, with simple seta on distal corner; merus 1.2x carpus, with two lateral setae, and with two setae (one simple, one bifurcated) on distal corner; propodus 2.6x dactylus, with strong bifurcated distal seta; dactylus terminated with two thin setae and one strong seta.
Pereopod 5 ()) basis 0.5x remaining articles combined, with simple seta on medial surface, and with simple seta on distal corner; ischium 0.5x merus, with two simple setae on distal corner; merus 1.2x carpus, with simple seta laterally; carpus 1.3x propodus, with two simple lateral setae, and with bifurcated seta on distal corner; propodus 2.6x dactylus, with strong bifurcated distal seta; dactylus terminated with two thin setae and one strong seta.
Uropod ()) peduncle about 0.5x pleotelson, with two setae on lateral margin. Endopod one-articulated, 4.5x peduncle, with a row of seven setae along lateral margin and bifurcated spine on lateral margin in distal half, terminated with setulose seta. Exopod two-articulated, subequal to endopod, article2 with row of seven setae along lateral margin, terminated with two stout setulose setae.
Description of male
Body from allotype (ZMBN 149217). BL = 7.0 mm. Ocular lobe rounded. Carapace ()) 1.6 L:W, strongly flattened, with two pairs of horns. Pseudorostral plates rounded. Antennal notch absent. Lateral carinae nearly reach the border of carapace.
Pereon ()) 0.6x carapace, first pereonite invisible. Pleon 1.2x carapace, with five pairs of pleopods.
Distribution
The Gulf of Guinea, off the coast of Ghana, Eastern Atlantic; depth 20–50 m.
Remarks
Due to the dorsoventrally flattened body and the presence of two pairs of horns, one dorsolateral and one lateral, the new species shows similarities to E. amakusense, E. cadenati, E. carinocurvum, E. hilgendorfi, E. latum, E. stelliferum, E. striatum, E. taprobanicum and E. winri. However, the new species can be distinguished from them by a combination of the following features: carapace slightly narrowing from the anterior to the posterior part, proportion of pereopod1: basis 0.5x remaining articles combined, carpus similar to propodus and dactylus 0.8x propodus. Moreover, E. guinean can be distinguished by the appearance of antennule: trapezoid article1, article2 0.6x article3, and the presence of bifurcated spine on uropod rami ().
Table II. Diagnostic features distinguishing E. guinean sp. nov. from E. amacusensis, E. cadenati, E. carinocurvum, E. hilgendorfi, E. latum, E. steliferra, E. striata, E. trapabanica and E. winri. Features: 1 – carapace L:W; 2 – ratio of carapace width at the widest line to the width at the posterior part; 3 – antennule article 1 shape; 4 – antennule article 1 to article 3; 5 – ratio of pereopod 1 basis to the remaining articles; 6 – ratio of pereopod 1 dactylus to propodus; 7 – ratio of pereopod 1 merus to carpus; 8 – ratio bifurcated setae on uropod. *based on male.
Eocuma mandeli sp. nov.
Registered under the ZooBank number urn:lsid:zoobank.org:pub:A4A175E4-91A9-41DF-B8D0-01C39CBDFCB6
Material
Holotype
Non-ovigerous female, ZMBN 149212 st. GW 1/0.
Paratype
Adult male, body drawn, ZMBN 149213 st. GW 1/0; one non-ovigerous female, dissected and drawn, ZMBN 149213 st. GW 1/0; 12 non-ovigerous females, four broken females, four juvenile males, and one broken male, ZMBN 149213 st. GW 1/0.
Etymology
The species name is dedicated to Nelson Mandela, the revolutionary political leader and Nobel Peace Prize laureate from South Africa.
Diagnosis
Carapace oval in dorsal view, covered with shallow holes, without lateral horns, with protruding tips of the subrostral tooth. Medial carina weakly marked, dorsolateral and transverse carinae form shallow depression. Pleon 1.7x carapace. Pereopod 1 basis 1.2x remaining articles combined, carpus 1.1x propodus, dactylus 0.7x propodus. Uropod exopod article 2 with smooth margin (no crenulation).
Description of female
Body from holotype (ZMBN 149212), appendages from paratypes (ZMBN 149213). BL = 4.7 mm. Ocular lobe rounded. Carapace ()) 1.2 L:W, oval in dorsal view, surface covered with shallow holes, with no lateral horns, with protruding tips of the subrostral tooth. Pseudorostral plates straight, with crenulated margin. Antennal notch well developed. Medial carina weakly marked, transverse carina in anterior part.
Figure 12. Eocuma mandeli sp. nov., non-ovigerous female, holotype (a, b: ZMBN 149212), and adult male, allotype (c, d: ZMBN 149213). a, Body dorsal view; b, body lateral view; c, body dorsal view; d, body lateral view. Scale bars = 1 mm.
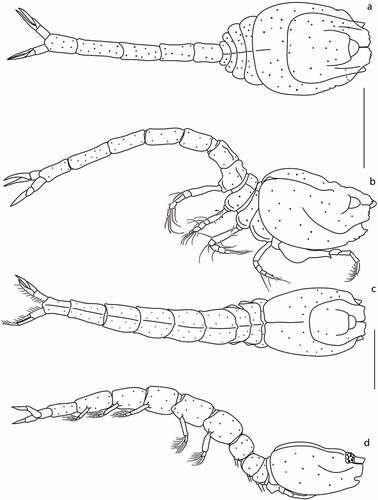
Pereon ()) 0.6x carapace, first pereonite invisible, pereonites 2–5 covered with shallow holes. Pleon 1.7x carapace.
Antennule ()) peduncle tri-articulated. Article 1 with parallel lateral margin, 2.2 L:W, 1.5x article 2, with no setae; article 2 0.6x article 3, with two penicillate setae on distal corner; article 3 similar in length to article 1. Main flagellum two-articulated, article 2 tipped with aesthetasc and two long setae. Accessory flagellum minute, tipped with two penicillate setae.
Figure 13. Eocuma mandeli sp. nov., non-ovigerous female, paratype (ZMBN 149213). a, Antennule; b, maxilla outer endite; b’, maxilla inner endite; c, maxillula; d, maxilliped 1; e, maxilliped 2; f, maxilliped 3. Scale bars = 0.1 mm.
Antenna not observed.
Left and right mandible not observed.
Maxilla ()) outer endite with 12 terminal spines (11 simple, 1 plumose) and with four simple subterminal setae: three on inner corner and one on outer corner. Inner endite (’)) with six plumose terminal setae.
Maxillula ()) broad endite with 17 subterminal setae, and with 16 simple setae and one plumose terminal seta. Outer endite with six plumose terminal setae, and with two simple subterminal setae. Inner endite with six plumose terminal setae.
Maxilliped 1 ()) basis stout, with a row of five plumose setae along inner margin, and with a bunch of setules on outer distal corner; endite with four plumose terminal setae (with thick basis), and with three plumose setae on medial surface; merus 0.4x carpus, with no setae; carpus 2x propodus, with seven flattened setae along lateral margin, with numerous setae on medial surface, and with seta on outer distal corner, and two short lateral setae on outer margin; propodus 2x dactylus, with 10 setae along inner and distal margin, and with two short setae and one long seta on outer margin laterally; dactylus with two simple terminal and three simple subterminal setae.
Maxilliped 2 ()) basis 0.8x remaining articles combined, with two plumose setae on inner distal corner, and with two short setae on outer margin; ischium 0.5x merus, with no setae; merus 0.4x carpus, with plumose seta on distal margin; carpus 1.2x propodus, with six setae along inner margin; propodus 1.6x dactylus, with four setae along inner margin, and with plumose seta on medial surface; dactylus 0.6x propodus, with two terminal setae, and with three subterminal setae.
Maxilliped 3 ()) basis 0.6x remaining articles combined, with crenulation along outer margin, distal prolongation reaching articulation of ischium and merus, with a row of eight plumose setae along inner margin, and with minute setae along outer margin; ischium 0.7x merus, with minute setae along inner margin; merus 1.4 L:W, 1.2x carpus, with plumose seta and a row of minute setae along inner margin, and with two plumose setae on outer distal corner; carpus 1.2 L:W, 1.4x propodus, with two simple setae on inner margin, and with short seta on outer distal corner; propodus 1.3 L:W, as long as dactylus, with two setae on inner margin; dactylus with three simple terminal setae, and with two simple subterminal setae.
Pereopod 1 ()) basis 1.2x remaining articles combined, with three rows of minute setae along outer margin; ischium subequal to merus, with short simple seta on lateral margin; carpus 1.1x propodus, with no setae; propodus 0.9x dactylus, with short simple setae on lateral margin, and with two distal setae; dactylus with a row of 10 plumose setae along lateral and terminal margin. Exopod shorter than basis.
Figure 14. Eocuma mandeli sp. nov., non-ovigerous female, paratype (ZMBN 149213). a, Pereopod 1; b pereopod 2; c, pereopod 3; d, pereopod 4; e, pereopod 5; f, uropod. Scale bars = 0.1 mm.
Pereopod 2 ()) basis 0.8x remaining articles combined, with no setae; ischium fused with basis, merus 0.9x carpus, with spine on distolateral corner; carpus 1.2x propodus, with long simple seta on distolateral corner; propodus 0.5x dactylus, with three simple terminal and one subterminal setae, and with two setae on lateral margin; dactylus 3.2 L:W, with two lateral setae and four setae (two short, two long) distally.
Pereopod 3 ()) basis 0.9x remaining articles combined, with two simple setae on proximal half and with two simple setae at distal half; ischium 0.7x merus, with two simple setae on distolateral corner; merus 1.3x carpus, with simple lateral seta; carpus 1.3x propodus, with two simple lateral setae, and one simple and one bifurcated distally; propodus 3.2x dactylus, with strong bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Pereopod 4 ()) 0.9x remaining articles combined, with four simple setae and two plumose setae on medial surface and laterally, and with long simple seta on distolateral corner; ischium 0.5x merus, with two simple setae on distolateral margin; merus 0.9x carpus, with two long setae and short simple setae on lateral margins, and with two setae distally: one simple and one bifurcated; propodus 3.2x dactylus, with strong bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Pereopod 5 ()) basis 0.9x remaining articles combined, with no setae; ischium 0.7x merus, with two simple setae on distolateral corner, merus 1.2x carpus, with no setae; carpus 1.2x propodus, with simple seta laterally, and with three distal setae: one short and simple, one long and simple, and one long and bifurcated; propodus 3.2x dactylus, with strong bifurcated distal seta; dactylus terminated with one thin seta and one strong seta.
Uropod ()) peduncle 0.4x pleotelson, with two setae on lateral margin. Endopod one-articulated, 2x peduncle, with row of 13 setae along lateral margin, with bifurcated spine on lateral margin in distal half, terminated with setulose setae. Exopod two-articulated, subequal to endopod, article 2 with seven setae along lateral margin, terminated with two stout setulose setae.
Description of male
Body and appendages from allotype (ZMBN 149213). BL = 5.3 mm. Ocular lobe rounded. Carapace ()) 1.3 L:W, oval in dorsal view, surface covered with shallow holes, with no lateral horns, with protruding tips of the subrostral tooth. Pseudorostral plates straight, with crenulated margin. Antennal notch well developed. Medial carina weakly marked, dorsolateral and transverse carinae form shallow depression in mid-lateral part.
Pereon ()) 0.7x carapace, first pereonite invisible, pereonites 2–5 covered with shallow holes. Pleon 1.6x carapace, with five pairs of pleonites.
Distribution
The Gulf of Guinea, off the coast of Ghana, Eastern Atlantic; depth about 25 m.
Remarks
The new species E. mandeli is similar to E. foveolatum, E. obriculatum and E. elongatum due to the lack of lateral horns and the presence of ornamentation, e.g. small and numerous holes on the surface of the body. However, the new species can be distinguished by:
The length and shape of the carapace in the female, 1.2 L:W and not tapering in anterior part; in E. foveolatum the carapace tapers anteriorly, in E. elongatum it is elongated (2 L:W), and in E. obriculatum it is 1.4 L:W;
The presence of the transverse carinae which are absent in E. obriculatum and E. foveolatum; E. elongatum has a transverse carina as in E. mandeli;
The maxilliped 3 merus is broad in E. mandeli and E. elongatum (1.5 L:W in both), but is elongated in E. foveolatum (2.7 L:W) and E. obriculatum (about 2 L:W);
The basis of pereopod 1 is 1.2x length of remaining articles combined in E. mandeli, but is shorter in E. foveolatum, E. obriculatum and E. elongatum (about 0.9-, 0.9- and 0.7-fold, respectively).
Eocuma nansenae sp. nov.
Registered under the ZooBank number urn:lsid:zoobank.org:pub:A4A175E4-91A9-41DF-B8D0-01C39CBDFCB6.
Material
Holotype
Brooding female, ZMBN 149204 st. G6/23.
Paratype
One non-ovigerous female, ZMBN 149201 st. G1/50; one female (poor condition), ZMBN 149202 st. G2/50; two non-ovigerous females, dissected, ZMBN 149203 st. G5/50; one non-ovigerous female, dissected and drawn, ZMBN 149204 st. G6/23; two non-ovigerous females, ZMBN 149205 st. G6/23; six non-ovigerous females, two juvenile males, one manca, ZMBN 149206 st. G6/51.
Etymology
The species is named after the R/V Fridtjof Nansen from which the samples were collected.
Diagnosis
Carapace oval in dorsal view, tapering towards the anterior, with surface smooth, and with lateral horns directed anteriorly, tips of horns reaching tip of pseudorostrum. Medial carina absent, dorsolateral carinae marked by a shallow indentation, only in anterior part. Pleon 1.4x carapace. Pereopod 1 basis 0.9x remaining articles combined, carpus 1.2x propodus, dactylus 0.6x propodus. Uropod exopod article 2 with smooth margin (no crenulation).
Description
Body from holotype (ZMBN 149204), appendage from paratype (ZMBN 149203). BL = 3.4 mm. Ocular lobe rounded. Carapace ()) 1.2 L:W, tapering towards the anterior, surface smooth, lateral horns directed anteriorly, horns short (about 1.5 L:W). Pseudorostral plates straight. Antennal notch missing. Medial carina absent, dorsolateral carinae marked by a shallow indentation, only in the anterior part.
Figure 15. Eocuma nansenae sp. nov., brooding female, holotype (ZMBN 149204). a, Body dorsal view; b, body lateral view. Scale bars = 1 mm.
Pereon ()) 0.5x carapace, first pereonite invisible. Pleon 1.4x carapace.
Antennule ()) peduncle tri-articulated. Article 1 with parallel lateral margin, 2.2 L:W, 2.2x article 2, with a lateral row of nine simple and three plumose setae; article 2 0.6x article 3, with simple lateral seta and with two simple distal seta; article 3 3 L:W, with penicillate distal seta. Main flagellum two-articled, article 2 with two aesthetascs and with two simple setae. Accessory flagellum minute, tipped with penicillate seta.
Figure 16. Eocuma nansenae sp. nov., non-ovigerous female, paratype (ZMBN 149203). a, Antennule; b, left mandible; c, right mandible; d, maxilla outer endite; d’, maxilla inner endite; d”, maxilla palp; e, maxillula; f, maxilliped 1; g, maxilliped 2; h, maxilliped 3. Scale bars = 0.1 mm.
Antenna not observed.
Left mandible ()) with a row of nine plumose setae, incisor with two teeth, lacinia mobilis with two teeth.
Right mandible ()) with a row of nine plumose setae, incisor with two teeth.
Maxilla outer endite ()) with 13 spines terminally, and five simple setae on outer subterminal margin. Inner endite (’)) with six simple setae and two strong plumose terminal setae. Palp (”)) with two long terminal setae.
Maxillula ()) broad endite with 12 subterminal setae, and with a row of 20 plumose terminal setae. Outer endite with five plumose terminal setae, and two simple setae on outer subterminal margin. Inner endite with six plumose terminal setae, and with simple short setae along inner margin.
Maxilliped 1 ()) basis stout; endite with five plumose strong terminal setae, and four plumose setae on medial surface, and with nine plumose setae along inner margin; merus 0.2x carpus, with no setae; carpus 2.6x propodus, with six flattened setae along inner margin, with numerous setae on medial surface, and with long plumose seta on outer distal corner; propodus 1.6x dactylus, with plumose seta on outer distal corner; dactylus with two simple terminal setae.
Maxilliped 2 ()) basis 0.7x remaining articles combined, with two plumose setae on inner distal corner, and with simple seta on outer distal corner; ischium short, with no setae; merus 0.8x carpus, with simple distal seta; carpus 0.8x propodus, with five plumose setae along inner margin; propodus 2.3x dactylus with six plumose setae along inner margin, and with long plumose seta and minute seta on outer distal corner; dactylus with two strong simple terminal setae, and with four simple subterminal setae.
Maxilliped 3 ()) basis stout, 0.7x remaining articles combined, with a row of short setae along lateral margins, and with plumose seta on inner distal corner, distal prolongation reaching articulation of ischium and merus, with six plumose setae along inner margin, and with two plumose setae on distal corner; ischium 0.5x merus, with three plumose setae along inner margin; merus 2 L:W, 2.8x carpus, with five plumose setae along inner margin, and with simple seta on outer distal corner; carpus 1.1 L:W, 0.5x propodus, with simple seta on outer distal corner; propodus 1.5 L:W, 1.4x dactylus, with two simple setae in distal half; dactylus with three simple terminal setae.
Pereopod 1 ()) basis 0.9x remaining articles combined, proximal part wider than distal, with no setae; ischium 0.4x merus, with short lateral seta; merus 0.8x carpus, with two simple lateral setae; carpus 1.2x propodus, with five simple lateral setae; propodus 1.4x dactylus, with two simple lateral setae, and with three distal setae; dactylus with three simple setae laterally, three terminal setae, and with simple subterminal seta. Exopod little longer than basis.
Figure 17. Eocuma nansenae sp. nov., non-ovigerous female, paratype (ZMBN 149203). a, Pereopod 1; b, pereopod 2; c, pereopod 3; d, pereopod 4; e, uropod. Scale bars = 0.1 mm.
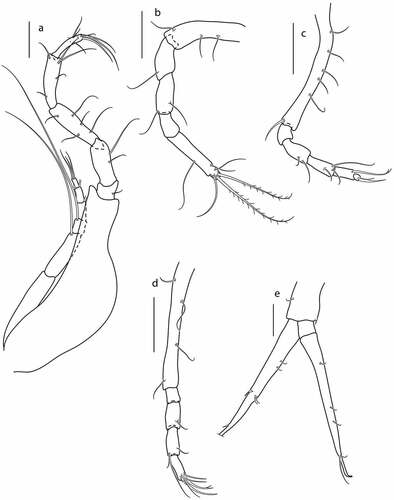
Pereopod 2 ()) basis 0.5x remaining articles combined, with three simple lateral setae; ischium fused with basis; merus subequal to carpus, with simple seta on distal corner; carpus 1.6x propodus with simple lateral seta, and simple seta on distal corner; propodus 0.3x dactylus, with no setae; dactylus 3.4 L:W, 2.7x propodus, with two simple setae laterally, two plumose setae terminally, and with two simple subterminal setae.
Pereopod 3 ()) basis 1.2x remaining articles combined, with a row of five simple setae along lateral margin, and with simple seta on distal corner; ischium 0.6x merus, with no setae; merus 1.1x carpus, with two simple setae laterally; carpus 1.5x propodus, with two simple setae distally, and seta on medial surface; propodus 4x dactylus, with bifurcated seta on distal corner; dactylus short, terminated with one thin seta and one strong seta.
Pereopod 4 ()) basis 1.3x remaining articles combined, with four simple setae on lateral margins; ischium 0.6x merus, with no setae; merus 0.9x carpus, with simple lateral seta; carpus as long as propodus, with simple lateral seta, and with two setae on distal corner; propodus 4x dactylus, with bifurcated seta and two simple setae on distal corner; dactylus short, terminated with one thin seta and one strong seta.
Pereopod 5 not observed.
Uropod ()) peduncle about 0.4x pleotelson, with short setae on lateral margin, and distal corner. Endopod one-articulated, with four simple setae on inner margin, and two spine on outer margin. Exopod two-articulated, subequal to endopod; second article with four simple setae along outer margin, terminated with two setulose setae.
Distribution
The Gulf of Guinea, off the coast of Ghana, Eastern Atlantic; depth 20–50 m.
Remarks
Eocuma nansenae, E. dollfusi, E. affine (only male known) and E. ferox share a similar distribution of the lateral horns and the shape of the carapace. However, the new species has a unique character in the form of the pseudorostrum tip which is located along the same line as the tip of the lateral horn. Other differences are:
The proportion of pereopod 1: in E. nansenae, dactylus is 0.6x propodus, while in E. dollfusi, dactylus is elongated (1.3x propodus), but it is short in E. affine and E. ferrox (0.8 and 0.7x propodus, respectively).
The length of the pleon which is 1.4x carapace in E. nansenae, but 1.7x carapace in E. dollfusi, 1.8x carapace in E. ferrrox (Bochert et al., Citation2011), () (E. affine: no information about female).
Eocuma olokunae sp. nov.
Registered under the ZooBank number urn:lsid:zoobank.org:pub:A4A175E4-91A9-41DF-B8D0-01C39CBDFCB6
Material
Holotype
brooding female, ZMBN 149208 st. G4/250.
Paratype
one non-ovigerous female, uncertain identification, ZMBN 149211 st. G1/50; one non-ovigerous female, dissected and drawn, ZMBN 149207 st. G1/250; one female in poor condition, ZMBN 149207 st. G1/250; one non-ovigerous female, ZMBN 149210 st. G6/51; one non-ovigerous female, ZMBN 149209 st. G6/250.
Etymology
Named after Olokun, the god of the ocean in the Yoruba religion practised in part of West Africa.
Diagnosis
Carapace oval in dorsal view, tapering in anterior direction, covered with small tubercles/spines, and with lateral horns, tip of which not reaching the tip of pseudorostrum. Medial carina weakly marked, dorsolateral carinae marked by shallow indentation. Pleon 2.5x carapace. Pereopod 1 basis 0.6x remaining articles combined, carpus 0.7x propodus, dactylus 0.4x propodus. Uropod exopod article 2 with crenulated margin.
Description
Body from holotype (ZMBN 149208), appendages from paratype (ZMBN 149207). BL = 3.3 mm. Ocular lobe oval. Carapace ()) 1.5 L:W, tapering in anterior direction, surface covered with small blunt spines/tubercles, lateral horns present, directed 45° in relation to lateral margin, horns elongated (about 3 L:W). Pseudorostral plates rounded. Antennal notch missing. Medial carina weakly marked, dorsolateral carinae marked only in anterior part, by shallow indentation.
Figure 18. Eocuma olokunae sp. nov., brooding female, holotype (ZMBN 149208). a, Body dorsal view; b, body lateral view. Scale bars = 1 mm.
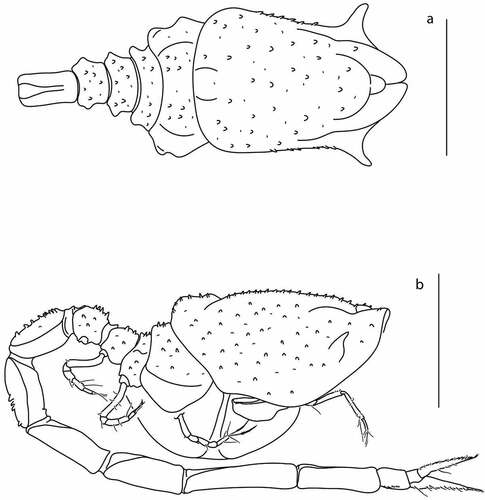
Pereon ()) 0.6x carapace, first pereonite invisible. Pereonites 2–5 covered with small blunt spines.
Pleon 2.5x carapace, first two pleonites covered with small blunt spines.
Antennule ()) peduncle tri-articled. Article 1 with parallel lateral margin, 2 L:W, 1.7x article 2, with no setae. Article 2 0.8x article 3, with two simple setae on subdistal corner. Article 3 3 L:W, with two penicillate setae on distal corner. Main flagellum two-articulated, article 2 with three terminal setae. Accessory flagellum one-articulated, with three penicillate terminal setae.
Figure 19. Eocuma olokunae sp. nov., non-ovigerous female, paratype (ZMBN 149207). a, Antennule; b, left mandible; c, right mandible; d, maxilla outer endite; d’, maxilla inner endite; e, maxillula; f, maxilliped 1; g, maxilliped 2; h, maxilliped 3. Scale bars = 0.1 mm.
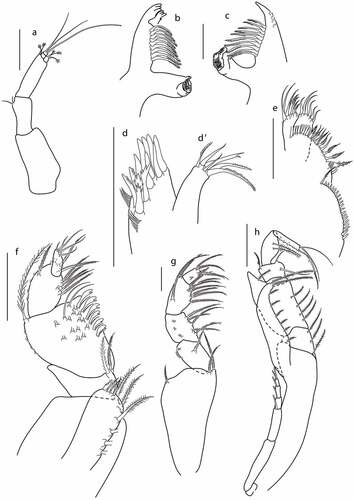
Antenna not observed.
Left mandible ()) with a row of nine plumose setae, incisor with three teeth, lacinia mobilis with four teeth.
Right mandible ()) with a row of 10 plumose setae, incisor with blunt tooth.
Maxilla ()) outer endite with seven simple spines, three trifurcated spines, and two plumose terminal spines, and with two rows of several subterminal setae. Inner endite (’)) with three plumose, two simple and one bifurcated terminal setae.
Maxillula ()) broad endite with 20 subterminal setae, and with row of 19 terminal setae. Outer endite with five plumose terminal setae, and two simple subterminal setae. Inner endite with six simple terminal setae.
Maxilliped 1 ()) basis stout, with no setae; endite with five plumose terminal setae, and two hook setae and a row of six plumose setae along inner margin; merus 0.2x carpus, with two plumose setae on inner distal corner; carpus 1.3x propodus, with six flattened setae accompanied by one thin seta along inner margin, with numerous setae on medial surface, and with plumose seta on outer distal corner; propodus 2x dactylus, with eight plumose setae along inner margin, and with plumose setae on outer margin; dactylus with three long and one short simple terminal setae.
Maxilliped 2 ()) basis 0.8x remaining articles combined, with two plumose setae on inner distal corner, and plumose seta on outer distal corner; merus 1.2x propodus, with plumose seta on distal surface, and numerous minute setae along outer margin; carpus 0.8x propodus, with five plumose setae along inner margin, propodus 2x dactylus, with seven plumose setae along inner margin, with plumose seta on outer distal corner, and with plumose seta on medial surface; dactylus with simple terminal seta, and five plumose subterminal setae.
Maxilliped 3 ()) basis stout, 0.9x remaining articles combined, distal prolongation reaching articulation of ischium and merus, with a row of six plumose setae along inner margin, and two plumose distal setae; ischium 0.3x merus, with three plumose setae; merus 2 L:W, 2.8x carpus, with four plumose setae on inner margin and plumose seta on subdistal corner; carpus 0.8 L:W, 0.6x propodus, with two setae on inner margin, and plumose seta on outer margin distally; propodus 1.3 L:W, 0.9x dactylus, with two plumose setae on inner margin; dactylus with one strong seta and three slender terminal setae.
Pereopod 1 ()) basis 0.6x remaining articles combined, proximal part wider than distal; with no setae; ischium 0.5x merus, with no setae; merus 0.6x carpus, with simple seta on inner margin; carpus 0.7x propodus, with two short setae laterally; propodus 2.5x dactylus, with two long setae, and four short setae laterally; dactylus 6.0 L:W, with two terminal and one subterminal seta, and four lateral setae in distal part. Exopod similar in length to basis.
Figure 20. Eocuma olokunae sp. nov., non-ovigerous female, paratype (ZMBN 149207). a, Pereopod 1; b, pereopod 2; c, pereopod 3; d, pereopod 4; e, pereopod 5; f, uropod. Scale bars = 0.1 mm.
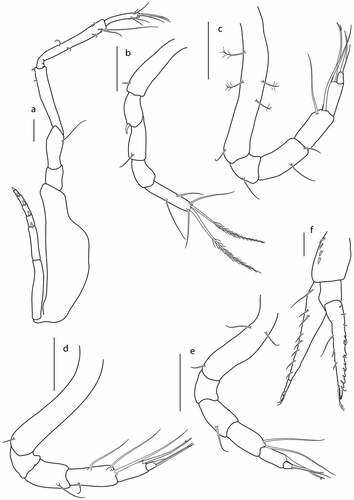
Pereopod 2 ()) basis 0.5x remaining articles combined, with short simple seta laterally; ischium fused with basis; merus 0.9x carpus, with spine on distal corner; carpus 1.3x propodus, with simple seta on lateral margin; propodus 0.6x dactylus, with no setae; dactylus 3.1 L:W, with two plumose terminal setae, two subterminal setae, and with two simple lateral setae.
Pereopod 3 ()) basis as long as remaining articles combined, with four penicillate setae laterally, and simple seta laterally in distal part; ischium 0.5x merus, with simple seta laterally; merus 1.2x carpus, with simple seta laterally, and simple distal seta; carpus 1.3x propodus, with one distal seta, and one seta subdistally; propodus 3x dactylus, with bifurcated seta on distal corner; dactylus short, terminated with one thin and one strong seta.
Pereopod 4 ()) basis 0.8x remaining articles combined, with simple lateral seta in distal part; ischium 0.6x merus, with no setae; merus 0.8x carpus, with simple lateral seta; carpus 1.5x propodus, with one distal, one subdistal and two simple setae on lateral margins; propodus 3.5x dactylus, with bifurcated seta on distal corner; dactylus short, terminated with one thin and one strong seta.
Pereopod 5 ()) basis 0.7x remaining articles combined, with two simple lateral setae in proximal part, and with simple lateral seta in distal part; ischium 0.7x merus, with no setae; merus as long as carpus, with simple lateral setae; carpus 1.4x propodus, with one seta on distal corner, and one subdistal seta; propodus 3.2x dactylus, with bifurcated seta on distal corner; dactylus short, terminated with one slender and one strong seta.
Uropod ()) peduncle 0.4x pleotelson, with a row of four setae along lateral margin. Endopod one-articulated, with a row of eight setae along outer margin, terminated with setulose seta. Exopod two-articulated, article 2 with a row of seven setae and crenulation on outer margin, with three setae on inner margin, and with two fine terminal setae.
Distribution
The Gulf of Guinea, off the coast of Ghana, Eastern Atlantic; depth 50–250 m.
Remarks
The presence of lateral horns and their direction renders E. olokunae similar to E. lanatum and E. aculeatum. The new species shares crenulation on the uropod exopod with E. lanatum, the presence of tubercles on the carapace being shared with E. aculeatum.
Eocuma olokunae can be distinguished from E. lanatum by the position of the horns. In the new species, the tip of the horns does not reach the tip of the rostrum, as is the case in E. lanatum. Moreover, the proportion of antennule article 3 is different: 3 L:W in E. olokunae sp. nov. and 7 L:W in E. lanatum (Bochert and Zettler Citation2011), (). Another small difference observed concerns the proportion of carpus to propodus in pereopod 1 which is 0.7x propodus in the new species and 0.5x in E. lanaum.
E. olokunae is distinguished from E. aculeatum by the absence of long minute setae that form a dense cover on the carapace of E. aculeatum. In addition, the proportion of pereopod 1 in both species is different: in the new species, carpus is shorter than propodus (0.7x), and propodus is 2.5x dactylus. In E. aculeatum, carpus is 1.3x propodus and 1.1x dactylus. Moreover, a strong spine on distal corner of pereopod 2 merus allows us to distinguish E. olokunae sp. nov. from E. aculaetum, with its unarmed pereopod 2 merus.
Discussion
Diversity
So far, the genus Eocuma has been known to include seven species from the tropical coasts of Africa (Jones Citation1957; Bochert & Zettler Citation2011). Jones (Citation1956) summarised the distribution of the shallow-water Cumacea and suggested tropical African coasts to support one of the poorest recognised fauna, compared to other tropical areas. However, Menegotto and Rangel (Citation2018) refer to a severe lack of funding for science and an inadequate scientific infrastructure as the real causes of the inadequate knowledge of the African marine fauna. Thus, the current state of knowledge is more of an artefact that needs to be readdressed, especially since the area is intensively exploited commercially for its oil and gas resources (Friedlander et al. Citation2014; Pabis et al. Citation2020).
Recent studies on peracarid crustaceans in the Gulf of Guinea reveal it is a repository of rare species sensitive to environmental factors (Jakiel et al. Citation2015; Buhl-Mortensen et al. Citation2017; Sobczyk et al. Citation2021; Jóźwiak et al. Citation2022). The high diversity and the presence of rare species allow us to regard the marine fauna of West Africa as unique and poorly known. For this reason, waters off the Atlantic coast of West Africa should be granted an effective environmental protection. Moreover, considering the general underestimation of marine diversity (Appeltans et al. Citation2012), it is expected that many more new peracarid species will be discovered, provided the earlier-unexplored areas are properly sampled and protocols appropriate for the collection and processing of macrobenthos samples are adhered to (Brandt et al. Citation2016; Frutos et al. Citation2022; Stefanni et al. Citation2022).
Phylogenetic analysis
The analysis presented here is the first attempt to explore the relationship between the Eocuma species and their possible routes of dispersal; it might be treated as a background for future studies. Nevertheless, the analysis carries a burden of many uncertainties. The high level of character polymorphism in the Cumacea (Haye Citation2007) in general, and in the genus Eocuma in particular, hinders recognition of clear patterns in its relationships. A similar problem is encountered in many other crustacean taxa. The high plasticity, sexual polymorphism and ontogenetic variation present obstacles to resolving the systematic position if the analysis is based on morphological characters only (Larsen & Wilson Citation2002). A more advanced analysis, including molecular assays, is needed here (Frutos et al. Citation2022).
All four trees developed here, and thus the consensus tree, revealed similar patterns, i.e. three sister groups within the genus Eocuma. The first group includes E. elongatum and E. victoriae, which originally were assigned to the genus Mossambicuma genera and later synonymised with Eocuma (Haye, Citation2007). Although the clade with these two species is separated, this separation is not supported by strong statistical evidence.
Other Eocuma species were grouped into one clade, with three species separated as ancestral. The most outlying species, E. fovellatum, lacks horns and its carpus and pereopod 3 propodus are of an almost equal length, as in Upselapsis which is regarded as the ancestral taxon. Eocuma dollfusi was separated because its pereopod 1 basis is shorter than the length of the ischium, merus, carpus, propodus and dactylus combined, and the dactylus is shorter than the propodus; the separation of E. nansenae is due to the pseudorostrum being situated at the level of the horns, and because of a narrow ocular lobe. Although these characters are highlighted as those separating E. fovellatum, E. dollfusi and E. nansenae sp. nov. from other Eocuma species, they can still be found in some Eocuma species, e.g. there is a lack of horns in E. ghanaian, or the pseudorostrum is at the level of horns in E. sarsii. All this confirms the relatively high level of polymorphism within Eocuma rather than proving that E. fovellatum, E. dollfusi and E. nansenae represent a distinct lineage.
The remaining Eocuma species were divided into two main lineages, the first containing species with a body of regular shape (oval in cross section) and the other including species with a flattened body, tapered posteriorly, with two pairs of horns. Although the division was present in all the four trees developed, the number of synapomorphic characters was too low to provide a strong statistic support. Moreover, some characters were not constant within the group, e.g. all of clade 56 species except E. ghanaian sp. nov. showing horns; all of clade 47 species showing horns; or most of clade 47 species having the carapace tapering towards the posterior, except for E. mandeli sp. nov. with the body oval in dorsal view.
Biogeography of Eocuma
Eocuma is a shallow-water taxon, occurring usually not deeper than 100 m (Day Citation1978; Corbera et al. Citation2005; Bochert & Zettler Citation2011), distributed in tropical and temperate zones. Seven new Eocuma species were collected on the Ghanaian shelf at depths of 23–100 m, but E. olokunae was also found deeper (250 m). So far, only E. aculeatum was found at a similar depth in the Mozambique Channel (Day Citation1978). The genus is clearly a warm-water taxon. The southernmost locations of Eocuma species are off South Africa (Day Citation1978); the northermost location was off the Kamchatka Peninsula in the Pacific (Marcusen Citation1894; Gamô Citation1967; Kim et al. Citation2020) and in the Atlantic’s Bay of Biscay (Fischer Citation1872; Calman 1907).
Together with the seven previously described species ( and ), the TA region supports 14 Eocuma species, which makes it the second most diverse region for the genus, after the Indian Ocean (WIP). The latter, together with the Red Sea and the Persian Gulf, hosts 15 Eocuma species. In contrast, the Central Indo-Pacific (CIP) and the Tropical South Africa (TSA) support five and three species, respectively (); four Eocuma species are known from both the north-west Pacific (TNP) and the Mediterranean Sea, two being known from the Bay of Biscay (TNA). Although relatively well studied, the Australian and New Zealand cumaceans (Petrescu Citation2004, Citation2006; Gerken, Citation2013, Citation2014, Citation2016, Citation2018) include as few as two Eocuma species.
Table III. Number of species shared between ecoregions. SA – South Africa, WIP – Western Indo-Pacific, TNP – Temperate North Pacific, TA – Tropical Atlantic, TNA – Temperate North Atlantic, and CIP – Central Indo-Pacific.
Figure 21. Worldwide distribution of Eocuma, including the newly described species from the Gulf of Guinea. Based on data in the literature: Fischer (Citation1872); Kossmann (Citation1880); Marcusen (Citation1894); Calman (Citation1904, Citation1907a, Citation1907b); Zimmer (Citation1914); Fage (Citation1928, Citation1950); Kurian (Citation1951, Citation1954); Gamô (Citation1967, Citation1976); Le Loeuff and Intès (Citation1972); Day (Citation1978); Kurian and Radha Devi (Citation1983); Mühlenhardt-Siegel (Citation1996); Petrescu (Citation1998, Citation2003); Patel et al. (Citation2003); Corbera et al. (Citation2005); Corbera and Galil (Citation2007); Bochert and Zettler (Citation2011); Kim et al. (Citation2020); Watling and Gerken (Citation2022).
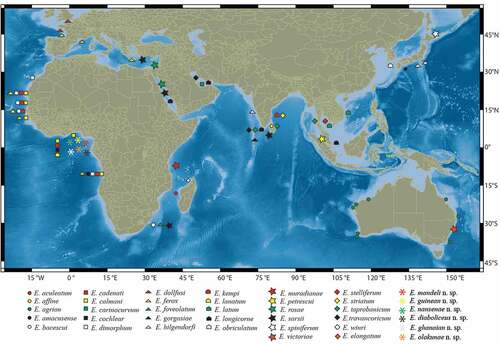
Results of our analysis show the fauna occurring off West Africa (TA) to be diverse, unique and clearly different from the faunas of other Atlantic and non-Atlantic locations. Only one species recorded off the African coast (E. ferrox) was found in the eastern and central Mediterranean Sea.
Briggs and Bowen (Citation2013) reported four potential sources of Eocuma species for the African fauna: (1) the Tethys Sea, (2) the Western Atlantic and Eastern Pacific before the Isthmus of Panama developed, (3) radiation in the Atlantic, and (4) the Indo-Pacific region via southern Africa. Given that Eocuma is a truly warm-water taxon, and that the fauna of south-east Africa and the Indian Ocean is clearly different from that of the western African coasts, the hypothesis of an Indian Ocean origin of the African fauna seems unlikely. Furthermore, the currents along the southern African coasts (i.e. the warm Agulhas Current and the cold Benguela Current) are not likely to facilitate migration of warm-water fauna from the Indian Ocean to the Atlantic. In the case of Eocuma species, the south-east African fauna is so different that the hypothesis of the Indian Ocean as the origin of the Atlantic fauna is impossible to defend. Although such a pattern of species dispersal has been reported for at least some animal groups (Floeter et al. Citation2008; Teske et al. Citation2014), the lack of fauna common to the Atlantic and non-Atlantic regions is evident and rules out such dispersal directions.
It seems much more likely that the TA fauna is a descendant of the western Atlantic–eastern Pacific, the dispersal happening about 2.8 Ma ago (O’Dear et al. Citation2016). This hypothesis, however, seems untenable as well, as no species of the genus Eocuma has been found in the western Atlantic fauna, and the family Bodotriidae is poorly represented in this part of the world in general (Fischer Citation1872; Fage Citation1928, Citation1950; Le Loeuff & Intès Citation1972). In the fourth possible scenario (Briggs & Bowen, Citation2013), the tropical Atlantic fauna could have evolved in situ. Considering the multidirectionality of the dispersal (e.g. from and to the place of origin) and that fauna at the site of origin may have become extinct, and further assuming that the place of origin supports the highest diversity, erroneous conclusions may be drawn (Briggs, Citation2000, Citation2003; Mironov Citation2006). At present, only one Eocuma species (E. ferox) has been reported from the western part of the Mediterranean Sea, and it is present along the western African coast. Although current distribution-based inferences regarding the place of origin are subject to some bias, it cannot be ruled out that the Mediterranean Sea, separated from the Tethys Sea by the Alpine orogeny prior to the Miocene, may have been the origin of the West African littoral fauna until the formation of the Strait of Gibraltar. Furthermore, the ancient Tethys Sea is regarded as a centre of radiation for the family Bodotriidae (Haye Citation2007). During the Triassic-Jurrasic mass extinction, the Tethys Sea became a centre of radiation for many taxa, including the Cumacea, which were later transported to the newly opening Atlantic and Indo-Pacific regions.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Appeltans W, Ahyong ST, Anderson G, Angel MV, Artois T, Bailly N, Bamber R, Barber A, Bartsch I, Berta A, Błażewicz-Paszkowycz M, Costello MJ. 2012. The magnitude of global marine species diversity. Current Biology 22:2189–2202. DOI: 10.1016/j.cub.2012.09.036.
- Bochert R, Zettler ML. 2011. Cumacea from the continental shelf of Angola and Namibia with descriptions of new species. Zootaxa 2978(1):1–33. DOI: 10.11646/zootaxa.2978.1.1.
- Brandt A, Gutt J, Hildebrandt M, Pawłowski J, Schwendner J, Soltwedel T, Thomsen L. 2016. Cutting the umbilical: New technological perspectives in benthic deep-sea research. Journal of Marine Science and Engineering 4(2):36. DOI: 10.3390/jmse4020036.
- Briggs JC. 2000. Centrifugal speciation and centres of origin. Journal of Biogeography 27:1183–1188. DOI: 10.1046/j.1365-2699.2000.00459.x.
- Briggs JC. 2003. The biogeographic and tectonic history of India. Journal of Biogeography 30:381–388. DOI: 10.1046/j.1365-2699.2003.00809.x.
- Briggs JC, Bowen BW. 2013. Marine shelf habitat: Biogeography and evolution. Journal of Biogeography 40(6):1023–1035. DOI: 10.1111/jbi.12082.
- Buhl-Mortensen L, Serigstad B, Buhl-Mortensen P, Olsen MN, Ostrowski M, Błażewicz-Paszkowycz M et al. 2017. First observations of the structure and megafaunal community of a large Lophelia reef on the Ghanaian shelf (the Gulf of Guinea). Deep Sea Research Part II Topical Studies in Oceanography 137:148–156. DOI: 10.1016/J.DSR2.2016.06.007.
- Calman WT. 1904. Report on the Cumacea collected by professor Herdman, at Ceylon, in 1902. The Royal Society, Supplementary Report 7:159–180.
- Calman WT. 1907a. Crustacea. 2–Cumacea. National Antarctic expedition. Natural History 2:1–6.
- Calman WT. 1907b. Sur quelques Cumacés de côtes de France. Bulletin du Muséum d’Histoire Naturelle 2:116. Available: https://www.biodiversitylibrary.org/item/27187#page/7/mode/1up.
- Coleman CO. 2003. “Digital inking”: How to make perfect line drawings on computers. Organism Diversity and Evolution 3(4):33–34. DOI: 10.1078/1439-6092-00081.
- Corbera J, Galil BS. 2007. Colonisation of the eastern Mediterranean by Red Sea cumaceans, with the description of a new species. Scientia Marina 71 (1): 29–36. https://doi.org/10.3989/scimar.2007.71n129 .
- Corbera J, Tirado P, Martin D. 2005. Cumaceans (Crustacea: Peracarida) from the Persian Gulf. Zootaxa 1087:1–31. DOI: 10.11646/zootaxa.1087.1.1.
- Day J. 1978. South African Cumacea, Part 2: Family Bodotriidae, subfamily Bodotriinae. Annals of the South African Museum 75(7):159–290.
- Fage L. 1928. Cumacés de la côte atlantique du Maroc. Bulletin of the Museum d’Histoire Naturelle in Paris 8(7–9):173–181.
- Fage L. 1950. Sur un nouveau Cumacé de la côte occidentale d’Afrique Eocuma cadenati nov. sp. Bulletin of the Museum of Histoire Naturelle in Paris 22(4):450–451.
- Fage L. 1951. Cumacés. Volume 3 (1). In: Expédition Océanographique Belge dans les Eaux Côtières Africaines de l´Atlantique Sud (1948–1949). Résultats Scientifiques. Vol. 3. Bruxelles: Institut Royal des Sciences Naturelles de Belgique. pp. 1–9.
- Fischer P. 1872. Pages 47, 110. In: de Folin L, Perier L, editors. Les Fonds de la mer, étude internationale sur les particularités nouvelles des régions sous-marines, commencée et dirigée. Vol. 2 (1). Paris: Savy. pp. 47, 110.
- Floeter SR, Rocha LA, Robertson DR, Joyeux JC, Smith‐Vaniz WF, Wirtz P, … Bernardi G. 2008. Atlantic reef fish biogeography and evolution. Journal of Biogeography 35(1):22–47. DOI: 10.1111/j.1365-2699.2007.01790.x.
- Friedlander AM, Ballesteros E, Fay M, Sala E. 2014. Marine communities on oil platforms in Gabon, West Africa: High biodiversity oases in a low biodiversity environment. PLoS One 9(8):e103709. DOI: 10.1371/journal.pone.0103709.
- Frutos I, Kaiser S, Pułaski Ł, Studzian M, Błażewicz M. 2022. Challenges and advances in the taxonomy of deep-sea Peracarida: From traditional to modern methods. Frontiers in Marine Science 9:799191. DOI: 10.3389/fmars.2022.799191.
- Gamô S. 1976. Eocuma spinifera sp. nov., a new cumacean Crustacea from Japan. Bulletin of the National Science Museum 2 (2): 93–102.
- Gamô S. 1967. Studies on the Cumacea (Crustacea, Malacostraca) of Japan. Part 1. Seto Marine Biological Laboratory 15:133–163.
- Gerken S. 2013. New Zealand Bodotriidae (Crustacea: Cumacea). Zootaxa 3630(1):1–38. DOI: 10.11646/zootaxa.3630.1.1.
- Gerken S. 2014. Eleven new species and a new genus of Diastylidae Crustacea: Cumacea) from Australia and one new species from Canada. Records of the Australian Museum 66(1):1–62. DOI: 10.3853/j.2201-4349.66.2014.1601.
- Gerken S. 2016. Leuconidae (Crustacea: Cumacea) from the collections of the Museum Victoria, Australia. Zootaxa 4067(3):251–292. DOI: 10.11646/zootaxa.4067.3.
- Giribet G. 2003. Stability in phylogenetic formulations and its relationship to nodal support. Systematic Biology 52(4):554–564. DOI: 10.1080/10635150390223730.
- Goloboff PA, Catalano SA. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32:221–238. DOI: 10.1111/cla.12160.
- Goloboff PA, Farris JS. 2001. Methods for quick consensus estimation. Cladistics 17:S26–S34. DOI: 10.1111/j.1096-0031.2001.tb00102.x.
- Goloboff PA, Farris JS, Nixon KC. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24:774–786. DOI: 10.1111/j.1096-0031.2008.00217.x.
- Goloboff PA, Mattoni CI, Quinteros AS. 2006. Continuous characters analyzed as such. Cladistics 22:589–601. DOI: 10.1111/j.1096-0031.2006.00122.x.
- Haye PA. 2007. Systematics of the genera of Bodotriidae (Crustacea: Cumacea). Zoological Journal of the Linnean Society 151:1–58. DOI: 10.1111/j.1096-3642.2007.00322.x.
- Jakiel A, Palero F, Błażewicz M. 2019. Deep ocean seascape and pseudotanaidae (crustacea: tanaidacea) diversity at the clarion-clipperton fracture Zone. Scientific Reports 9(1):1–49. DOI: 10.1038/s41598-019-51434-z.
- Jakiel A, Stępień A, Jóźwiak P, Serigstad B, Błażewicz-Paszkowycz M. 2015. First record of Tanaidacea (Crustacea) from a deep-sea coral reef in the Gulf of Guinea. Zootaxa 3995:203–228. PMID: 26250314. DOI: 10.11646/zootaxa.3995.1.18.
- Jones NS. 1956. Cumacea from the west coast of Africa. Atlantide Report 4:183–212. Available: https://eurekamag.com/research/022/307/022307287.php.
- Jones NS. 1957. Cumacea, Key to families and references. Conseil International Pour l’Exploration de la Mer 71:1–3.
- Joy JB, Liang RH, McCloskey RM, Nguyen T, Poon AFY. 2016. Ancestral reconstruction. PLoS Computional Biology 12(7):e1004763. DOI: 10.1371/journal.pcbi.1004763.
- Jóźwiak P, Pabis K, Sobczyk R, Serigstad B. 2022. A paradise for rare species: Tanaidacean Fauna of the West African Continental margin. Frontiers in Marine Science 9. DOI: 10.3389/fmars.2022.779134.
- Kim SH, Lee CM, Kim YH. 2020. Eocuma orbiculatum sp. nov. (Crustacea, Cumacea, Bodotriidae) from Korean waters. ZooKeys 910:79–91. DOI: 10.3897/zookeys.910.47143.
- Kossmann R. 1880. Malacostraca. Zoologische Ergebnisse einer ausgeführten Reise in die Küstengebiete des Rothem Meeres 2:88–90.
- Kurian CV. 1951. The Cumacea of Travancore. Bulletin Centre Research Institute Universitary Travancore 2:77–118.
- Kurian CV. 1954. Notes on Cumacea (Sympoda) in the zoological survey of India. Records of the Indian Museum 52:275–312.
- Kurian CV, Radha Devi A. 1983. A new species of Cumacea (Crustacea, Peracarida) from Vizhinjom, Kerala, India, and John PA, editor. Selected papers on Crustacea. The Aquarium, Trivandrum. pp. 149–153.
- Larsen K, Wilson GDF. 2002. Tanaidacean phylogeny, the first step: The superfamily Paratanaidoidea. Journal of Zoological Systematics and Evolutionary Research 40(4):205–222. DOI: 10.1046/j.1439-0469.2002.00193.x.
- Le Loeuff P, Intès A. 1972. Les Cumacés du plateau continental de Côte dIvoire. Cahiers ORSTOM, Série Océanographie 10(1):19–46.
- Marcusen J. 1894. Ueber ein neues Cumaceengenus Eocuma Family Cumadae, aus Japan. Sitzungsberichte der Gesellschaft naturforschender Freunde zu Berlin 1894:170–171.
- Menegotto A, Rangel TF. 2018. Mapping knowledge gaps in marine diversity reveals a latitudinal gradient of missing species richness. Nature Communications 9(1):1–6. DOI: 10.1038/s41467-018-07217-7.
- Mironov AN. 2006. Centers of marine fauna redistribution. Zoologicheskii Zhurnal 85:32–47. DOI: 10.1134/S0013873806100034.
- Mühlenhardt-Siegel U. 1996. Cumacea (Crustacea) from the Red Sea and the Maldives (Indian Ocean) in the collection of the Zoological Museum, Hamburg, with the description of seven new species and a new genus. Beaufortia 46(7):105–134. Available: ark:/13960/t8md7mf71.
- Mühlenhardt-Siegel U. 2003. Shallow-water cumacean Crustacea from Australia and Lombok (Indonesia): Families Bodotriidae and Leuconidae. Records of the South Australian Museum (Adelaide) 36(1):21–25. Available from: https://biostor.org/reference/114079 .
- O’Dea A, Lessios HA, Coates AG, Eytan RI, Restrepo-Moreno SA, Cione AL, Collins LS, De Queiroz A, Farris DW, Norris RD, Stallard RF, Woodburne MO, Aguilera O, Aubry MP, Berggren WA, Budd AF, Cozzuol MA, Coppard SE, Duque-Caro H, Finnegan S, Gasparini GM, Grossman EL, Johnson KG, Keigwin LD, Knowlton N, Leigh EG, Leonard-Pingel JS, Marko PB, Pyenson ND, Rachello-Dolmen PG, Soibelzon E, Soibelzon L, Todd JA, Vermeij GJ, Jackson JBC. 2016. Formation of the Isthmus of Panama. Science Advances 2:1–11. DOI: 10.1126/sciadv.1600883.
- The Oil for Development (OfD) Programme, Annual reports. 2019. Published in June 2020. Available: https://www.norad.no/en/toolspublications/publications/2020/the-oil-for-development-programme-2019/.
- Pabis K, Sobczyk R, Siciński J, Ensrud T, Serigstadt B. 2020. Natural and anthropogenic factors influencing abundance of the benthic macrofauna along the shelf and slope of the Gulf of Guinea, a large marine ecosystem off West Africa. Oceanologia 62:83–100. DOI: 10.1016/j.oceano.2019.08.003.
- Patel CK, Haye PA, Kornfield I. 2003. Eocuma petrescui, a new species of bodotriid cumacean (Crustacea; Peracarida) from Malaysia. Proc Biol Soc Wash 116:978–985. Available: (PDF) Eocuma petrescui, a new species of bodotriid cumacean (Crustacea: Peracarida) from Malaysia (researchgate.net)
- Petrescu I. 1998. Cumaceans (Crustacea: Cumacea) collected by the expedition of “Grigore Antipa” Museum of Natural History from the coasts of Tanzania (1973–1974). Part I. Family Bodotriidae. Travaux du Museum National d’Histoire Naturelle “Grigore Antipa” 40:227–310.
- Petrescu I. 2003. First record of Cumacea (Crustacea: Peracarida) from Madagascar. Travaux du Muséum d’Histoire Naturelle Grigore Antipa 65:95–115.
- Petrescu I. 2004. The first mention of the genus Apocuma Jones, 1973 (Crustacea: Cumacea: Bodotriidae) from Australian waters. Travaux du Muséum d’Histoire Naturelle Grigore Antipa 66:45–54.
- Petrescu I. 2006. Nannastacidae (Crustacea: Cumacea) from eastern Bass Strait, the south-eastern Australian slope, and Antarctica in the collections of Museum Victoria. Memoirs of Museum Victoria 63(2):129–173. DOI: 10.24199/j.mmv.2006.63.14.
- Petrescu I. 2018. On the family Nannastacidae (Crustacea, Cumacea) from the Australian Museum collection. Records of the Australian Museum 1(70):1–111. DOI: 10.3853/j.2201-4349.70.2018.1645.
- Sobczyk R, Czortek P, Serigstad B, Pabis K. 2021. Modelling of polychaete functional diversity: Large marine ecosystem response to multiple natural factors and human impacts on the West African continental margin. Science of the Total Environment 792:148075. DOI: 10.1016/j.scitotenv.2021.148075.
- Spalding MD, Fox HE, Allen GR, Davidson N, Ferdaña ZA, Finlayson MAX, Halpern BS, Jorge MA, Lombana AL, Lourie SA, Martin KD, Robertson J. 2007. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 57(7):573–583. DOI: 10.1641/B570707.
- Stefanni S, Mirimin L, Stankovic D, Chatzievangelou D, Bongiorni L, Marini S, Modica MV, Manea E, Bonofiglio F, Río JD, Cukrov N, Aguzzi J. 2022. Framing cutting-edge integrative deep-sea biodiversity monitoring via environmental DNA and optoacoustic augmented infrastructures. Frontiers in Marine Science 8:797140–797141. DOI: 10.3389/fmars.2021.797140.
- Stȩpień A, Pabis K, Sobczyk R, Serigstad B. 2021. Extremely low abundance of Cumacean communities along the shelf and slope of the Gulf of Guinea (West Africa). Frontiers in Marine Science 8. DOI: 10.3389/fmars.2021.703547.
- Teske PR, Papadopoulos I, Barker NP, McQuaid CD, Beheregaray LB. 2014. Mitonuclear discordance in genetic structure across the Atlantic/Indian Ocean biogeographical transition zone. Journal of Biogeography 41(2):392–401. DOI: 10.1111/jbi.12201.
- Ukwe CN, Ibe CA, Sherman K. 2006. A sixteen-country mobilization for sustainable 734 fisheries in the Guinea current large marine ecosystem. Ocean & Coastal Management 49(7):385–412. DOI: 10.1016/j.ocecoaman.2006.04.006.
- Watling L, Gerken S. 2022. World Cumacea database. Available: https://www.marinespecies.org/cumacea. Accessed Mar 2022 30.
- Yu Y, Harris AJ, Blair C, He X. 2015. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Molecular Phylogenetics and Evolution 87:46–49. DOI: 10.1016/j.ocecoaman.2006.04.006.
- Zimmer C. 1903. Die Cumaceen des Museums für Naturkunde in Berlin. Zoologische Jahrbücher, Abtheilung für Systematik, Geographie und Biologie der Thiere 18(6):664–694.
- Zimmer C. 1914. Cumacea. Die Fauna Südwest-Australiens 5:176–195.