
Abstract
Acerentulus panamensis sp. nov. from the tropical forest of the Gigante Peninsula of Barro Colorado Nature Monument (9°06’N, 79°54’W – Panama) is described. The new species belongs to the cunhai group being characterized by a short sensillum a not reaching the base of seta γ3 and by sensillum b shorter than c. Acerentulus panamensis sp. nov. is close to A. gerezianus da Cunha, 1952, A. ladeiroi Da Cunha, 1950, A. omoi Imadaté, 1988 and A. catalanus Condé, 1951. It differs from the first species by the absence of seta Pc on sternite VII, by the chaetotaxy of tergites II–V (6/16 in the new species and 8/14 in A. gerezianus), by the longer foretarsal sensillum e and by shorter sensilla f and a’. Finally, in Acerentulus panamensis sp. nov. sensillum a’ is at the same level of t1 closer to the base of tarsus than in A. gerezianus. In A. ladeiroi, the chaetotaxy of tergites II–V is 8/14 (as in A. gerezianus). Moreover, A. ladeiroi is distinguishable from the new species by shorter foretarsal sensilla e and t2, by a longer sensillum f, and by a longer sensillum a’ placed distally to t1. The new species differs from A. omoi by the absence of seta Pc on sternite VII, by the porotaxy of sternites I–III, and by the longer foretarsal sensillum e and shorter sensillum b. Acerentulus panamensis sp. nov. differs from A. catalanus by the chaetotaxy of tergites II–V, by the shorter sensilla a, b and f, and by the number of teeth of comb VIII (11–12 vs 7–8).
https://zoobank.org/urn:lsid:zoobank.org:pub:19B1F711-95B9-47DA-8157-98327ADEF104
Introduction
To date, nearly 850 species of Protura, belonging to three orders, seven families and 77 genera are known (Galli Citation2022) described from all continents but Antarctica (Galli & Rellini Citation2020). In Central and South America, this taxon of tiny soil arthropods has been the subject of few specific studies in a limited number of sites (see Szeptycki Citation2007; Palacios-Vargas & Figueroa Citation2014; Galli et al. Citation2021; Román de la Fuente Citation2024). Overall, from this area 59 species belonging to 15 genera have been recorded (). Three genera are endemic of the Neotropical region, Amazonentulus Yin, 1989, Andinentulus Tuxen, Citation1984 and Brasilidia Nosek, Citation1973 (Galli & Rellini Citation2020; Galli Citation2022), while a species not identified with certainty, but very similar to Amphientulus ambiguus (Tuxen, Citation1967) should be added to the species listed in (Tuxen Citation1984).
Table I. List of protura species from the Neotropical region.
The cosmopolitan genus Acerentulus Berlese, 1908 (Acerentomidae, Berberentulinae) is characterized by tufted maxillary and labial palps, canal of maxillary glands with simple dilation and a rather long proximal part, foretarsal sensillum t1 claviform, t3 willow leaf-shaped, a’ usually broad, abdominal appendages II–III unisegmented each with three setae (a long subapical, a shorter lateral apical, and the median apical the shortest), seta P3 on tergites II–V/VI placed anteriorly to the row of the other P-setae, sternites I–VII with three anterior setae and VIII with 4/2 setae, and striate band on abdominal segment VIII well developed (Galli et al. Citation2018). This genus includes 49 species arranged into three groups based on the length of foretarsal sensilla a, b and c: confinis, cunhai and traegardhi (Nosek Citation1973). In particular, in the Neotropical region only two species of this genus were known to date: Acerentulus christensoni (Ewing, 1940) 60 from Mexico and Acerentulus nemoralis Najt and Vidal Sarmiento, 1970 from Argentina (Shrubovych et al. Citation2014a), both belonging to the cunhai group. The species belonging to the cunhai group are characterized by a short sensillum a not reaching the base of seta γ3 and by sensillum b shorter than or subequal to c (Shrubovych et al. Citation2014b).
In this paper, a new species of proturan belonging to genus Acerentulus cunhai group is described based on material collected during research about the response of the soil arthropod community to litter manipulation in the tropical rainforest of Panama (Ashford et al. Citation2013), thus bringing to three the number of neotropical species of this genus.
Material and methods
Specimens were examined and identified at species and life-stage levels using an interference contrast microscope (Leica DM LB2 – Wetzlar, Germany), a Leica DFC 295 camera (Heerbrugg, Switzerland) and Leica Application Suite Vers. 3.8 (Heerbrugg, Switzerland).
These tools were also used to take diagnostic measurements of each specimen (see Nosek Citation1973). In the morphological description, the terminology adopted by Szeptycki (Citation1991), Rusek et al. (Citation2012), and Shrubovych (Citation2014) was used. Finally, ratios between measurements were indicated with acronyms as follows:
PR = pseudoculus ratio = length of the head (without rostrum)/length of pseudoculus
CF = head length/hind part of maxillary gland length
TR = tarsus ratio = length of foretarsus/length of claw
EU = empodium/unguis ratio = length of empodium/length of foretarsal claw
BS = base/sommet ratio = distance of foretarsal sensillum t1 from the base of tarsus/distance of t1 from the apex of tarsus.
Description
Acerentulus panamensis sp. nov
–8; .
Figure 5. Acerentulus panamensis sp. nov. (a) head in dorsal view; (b) pseudoculus; (c) maxillary glands; (d) maxillary palp; (e) labial palp (sensillum is marked); (f) foretarsus, dorsal view; (g) foretarsus, ventral view. Scale bars: (a, f, g) 50 µm, (b‒e) 20 µm. Figures (f, g): holotype. Figures (a‒e): paratype. Abbreviations of setae names: l = lateral, sd = subdorsal. Abbreviations of pore names: fp = frontal pore.
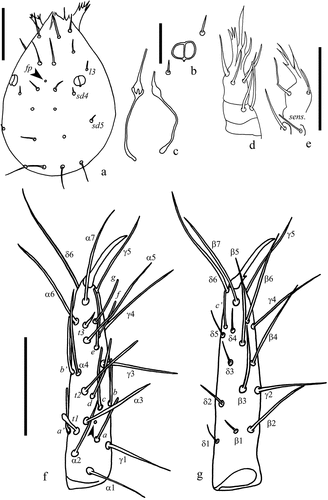
Figure 6. Acerentulus panamensis sp. nov. (a) pro- and mesonotum with detail (magnif. 5×) of seta P1a; (b) metanotum; (c) abdominal appendage II; (d) comb on tergite VIII. Scale bars: (a, b) 50 µm, (c, d) 20 µm. Figures (a‒d): holotype. Abbreviations of setae names: A = anterior, a = accessory, M = median, P = posterior. Abbreviations of pore names: sl = sublateral.
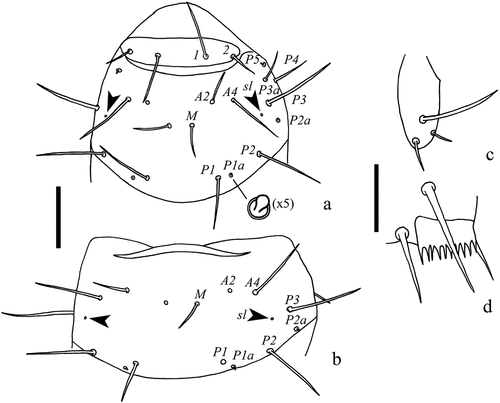
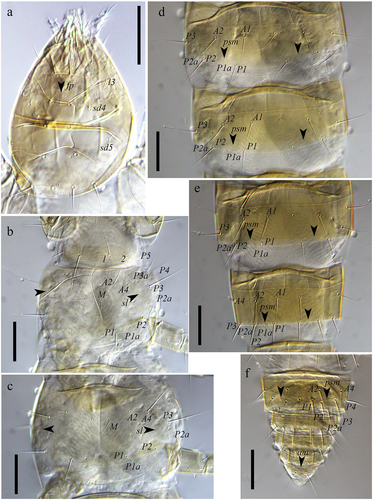
Figure 7. Acerentulus panamensis sp. nov. (a) tergite V; (b) tergite VI; (c) tergite VII; (d) tergites VIII‒XII; (e) thorax I‒II, ventral view; (f) thorax III, ventral view; (g) abdomen V, ventral view; (h) sternite VI of female. Arrows indicate pores. Scale bar: 50μm. Figures (a‒g): holotype. Figure (h): paratype. Abbreviations of setae names: A = anterior, a = accessory, c = central, M = median, P = posterior. Abbreviations of pore names: al = anterolateral, am = anteromedial, psm = posterosubmedial, sc = sternal central, spsm = sternal posterosubmedial.
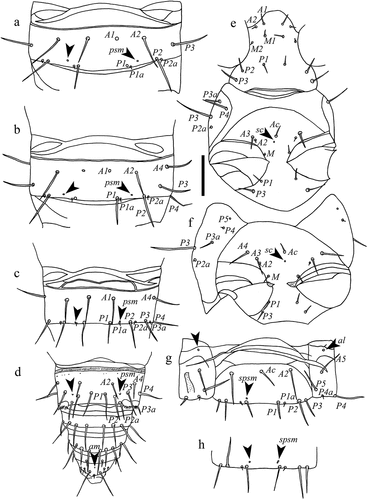
Figure 8. Acerentulus panamensis sp. nov. (holotype) (a) abdomen VI‒VII, ventral view; (b) abdomen VIII–XII, ventral view. Arrows indicate pores. Scale bar: 50 µm. Abbreviations of setae names: A = anterior, a = accessory, c = central, P = posterior. Abbreviations of pore names: al = anterolateral, sal = sternal anterolateral, spm = sternal posteromedial, spsm = sternal posterosubmedial.
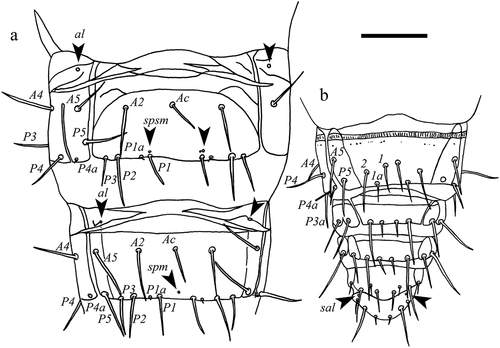
Table II. Acerentulus panamensis sp. nov. chaetotaxy. Setae nomenclature (abbreviations): A = anterior, a = accessory, c = central, M = median, P = posterior.
Table III. Acerentulus panamensis sp. nov. porotaxy. Pores nomenclature (abbreviations): al = anterolateral, am = anteromedial, psl = posterosublateral, psm = posterosubmedial, sal = sternal anterolateral, sc = sternal central, sl = sublateral, spm = sternal posteromedial, spsm = sternal posterosubmedial.
Material examined
Holotype: one male from the Gigante Peninsula of Barro Colorado Nature Monument (9°06’N, 79°54’W), Republic of Panama, lowland tropical forest (), August 2010, coll. Ashford et al. Paratype: one female, same data as the holotype. Type material () is preserved in the Museum of Department of Earth, Environment and Life Sciences, Genoa University, section Zoology (registration numbers IZUG-16116 and IZUG-16117).
Figure 1. Sampling area. (a) the Gigante peninsula of Barro Colorado nature monument, Panama; (b, c) lowland tropical forest (courtesy of Dumas Gálvez).

Figure 2. Acerentulus panamensis sp. nov. (interference contrast microscope) (a) holotype; (b) paratype.

Description
Measurements and ratios refer to both specimens: those relating to the paratype are reported in brackets. When only one value is reported, unless otherwise indicated, it means that the measurement/ratio was the same for both specimens.
Body length: 1412 (1450) μm.
Head 145 (147) μm long in dorsal view. Setae sd4, sd5 and l3 short and thin; seta d6 absent. Only frontal pore fp present (). Pseudoculus broader than long, 9 × 11(9 × 12) μm, longitudinally divided (); PR = 16. Maxillary gland with rounded calyx, long and slender posterior filament and simple posterior dilation (); proximal part 31 (38) μm long, CF = 4.7 (3.9). Maxillary palps with two slender sensilla, subequal in length (). Labial palps fully developed with apical tuft of setae and a slender sensillum ().
Figure 3. Acerentulus panamensis sp. nov. (interference contrast microscope) (a) head, dorsal view; (b) pro- and mesonotum; (c) metanotum; (d) tergites IV‒V; (e) tergites VI‒VII; (f) tergites VIII‒XII. Arrows indicate pores. Scale bars 50 µm. Figures (b‒f): holotype. Figure (a): paratype. Abbreviations of setae names: A = anterior, a = accessory, l = lateral, M = median, P = posterior, sd = subdorsal. Abbreviations of pore names: am = anteromedial, fp = frontal pore, psm = posterosubmedial, sl = sublateral.
Foretarsus length 106 (103) μm; claw without inner tooth or outer flap, long 31 (35) μm; TR = 3.4 (2.9); empodium length 7 μm (measurable only in the holotype), EU = 0.23; S-shaped seta longer than claw, 37 μm (measurable only in the holotype). Sensillum t1 claviform, placed 30 (29) μm from the base of tarsus (BS = 0.39); t2 thin, t3 shaped like a willow leaf. All other sensilla slender. Sensillum a slightly passing the base of d; b shorter than c (24 vs 32 μm); d long, reaching the base of f; e long, reaching the base of claw; f short, not reaching the base of claw, a bit nearer to e than to g; g longer than f, passing the base of claw. Sensillum a’ placed at the same level of t1, short, not reaching base of b’; b’ slightly passing base of c’; c’ short, not reaching base of claw. Ventral seta β1 shorter than interior seta δ4; δ4 situated proximally to c’ (). Foretarsal pore present near sensillum c (). Middle tarsus length 49 μm; claw length 20 μm. Hind tarsus length 54 (55) μm; claw length 23 (21) μm.
Chaetotaxy given in (see also and ). Length ratio of setae 1:2 on pronotum as 2.1:1 (1.9:1); ratio P1:P2 on mesonotum as 1:1.1. Posterior accessory setae on nota as short gemmate microchetae (). Abdominal tergites I–V each with three pairs of anterior setae (A1, A2, A5). All posterior accessory setae short (4‒5 μm), thin, and setiform. Seta P3a absent on tergites I–VI. Seta P3 anterior to the row of the other P-setae on tergites II–VI. Posterior margin of pleurite VIII smooth ()). Chaetal variability: in the holotype, P5 is asymmetrically missing on tergite II.
Figure 4. Acerentulus panamensis sp. nov. (interference contrast microscope) (a) prosternum; (b) mesosternum; (c) metasternum; (d) abdomen IV‒V, ventral view; (e) abdomen VI‒VII, ventral view; (f) abdomen VIII–XII, ventral view; (g, h) detail of spsm pores on female and male sternite VI; (i) comb on abdominal tergite VIII. Arrows indicate pores. Scale bars: (a‒f) 50 µm, (g‒i) 20 µm. Figures (a‒f, h, i): holotype. Figure (g): paratype. Abbreviations of setae names: A = anterior, a = accessory, c = central, M = median, P = posterior. Abbreviations of pore names: al = anterolateral, psl = posterosublateral, psm = posterosubmedial, sc = sternal central, spsm = sternal posterosubmedial.

Porotaxy given in . On meso- and metanotum sl pores present ( and ). Psm pores present on abdominal tergites I‒VIII; pores psm on tergites I‒VII flanked by two tiny teeth; al on tergites II‒VII; psl on VI and VII. Meso- and metasternum with sc pore () and ). Abdominal sternites I‒IV without pores. Sternite V with one asymmetrical spsm pore near the base of P1 (). Sternite VI with composed spsm pores (1‒2) symmetrically placed anterior to P1 (2 + 2 in the holotype male, 1 + 2 in the paratype female) (). Sternite VII with a spm pore near its hind margin, between P1 setae ().
Connecting lines in the anterior part of tergites and sternites IV‒VI present. Abdominal appendages II and III typical of genus Acerentulus (). Granular latero-dorsal area on tergite V posteriorly opened (). Striate band on abdominal segment VIII well developed, with distinct striae and with an irregular posterior row of barely visible granules ()); comb with 11–12 rather regular teeth ( and ). Male and female genitalia visible only at the level of acrostyles but too diaphanous in the remaining part for a reliable description. Female acrostyles long and pointed.
Ethymology
The name panamensis refers to the country where type specimens have been collected.
Diagnosis
Acerentulus panamensis sp. nov. belongs to cunhai species group (Nosek Citation1973). The new species is close to A. gerezianus da Cunha, 1952 from South Europe (and doubtfully Azores), A. ladeiroi da Cunha, 1950 from the Iberian Peninsula and Madeira (doubtfully Bosnia), A. omoi Imadaté, 1988 from Japan and A. catalanus Condé, 1951 from South Europe in having tergites I–III with seta P1a, tergite VI without seta P3a, and tergite VII with eight anterior setae. The first two species share with A. panamensis sp. nov. also the presence of a foretarsal sensillum b shorter than c. However, in A. gerezianus, there are 9 posterior setae on sternite VII, including Pc (which is absent in the new species). Moreover, in the European species chaetotaxy of tergites II–V is 8/14 while in the new species is 6/16. In addition, A. gerezianus has shorter foretarsal sensillum e and longer sensilla f and a’. Finally, in Acerentulus panamensis sp. nov. sensillum a’ is at the same level of t1 closer to the base of tarsus than in A. gerezianus. In A. ladeiroi the chaetotaxy of tergites II–V is 8/14 (as in A. gerezianus), differing from that of A. panamensis where it is 6/16. Moreover, A. ladeiroi is distinguishable from the new species by shorter foretarsal sensilla e and t2, by a longer sensillum f, and by a longer sensillum a’ reaching b’ and placed distally to t1.
In A. omoi, there are nine posterior setae on sternite VII, including Pc (which is absent in the new species) and there is a single spm or spsm pore on sternites I–III (while in A. panamensis pores are absent on these sternites). Moreover, the Japanese species has a longer sensillum b (nearly as long as c), and a shorter sensillum e, slightly passing the base of t3. On the other hand, A. catalanus has 8/14 setae on tergites II–V (vs 6/16 in the new species), a longer sensillum a passing the base of d, sensilla b and c subequal in length, a longer sensillum f reaching the base of claw, and comb VIII with only 7–8 teeth (vs 11–12 of A. panamensis).
It is possible that the presence on sternite VI of 2 + 2 spsm pores in the male specimen and 1 + 2 in the female of the new species is due to a sexual dimorphism of porotaxy already known in other Acerentulus species (see Szeptycki Citation1993).
Discussion
Protura are poorly known, and the fauna of many countries or large parts of them is still unstudied due to the very low number of specialists (see Galli & Rellini Citation2020; Galli Citation2022). In particular, in Central and South America, research on proturans was limited to a few localities and the total number of species amounts to only 59 (see also Shrubovych et al. Citation2014a). In the present paper, the first record of Protura from Panama is reported with the description of a new species of Acerentomidae. The fact that the only two specimens currently available for the entire country belong to a new species should be considered symptomatic of the incomplete level of knowledge of the fauna of this area.
Acerentulus panamensis sp. nov. brings to 60 the number of species known in the Neotropical region. Furthermore, genus Acerentulus rises to 50 species described worldwide, three of which, all belonging to the cunhai group, are reported from Central and South America. However, taking into account the low number of sites studied within the entire biogeographical region and also the growing evidence about the presence of species complexes among Protura (see Caterino & Recuero Citation2023), it seems very likely that much of the biodiversity of this group in the area remains undiscovered. This paper, therefore, represents a further small contribution in the direction of filling this gap.
Acknowledgments
We wish to thank Professor Dumas Gálvez (Universidad de Panamá) for kindly making available photos of the area where specimens were collected. The laboratory activity in Genoa University was supported by Genoa University Research funds (FRA Fondi per la Ricerca di Ateneo 2022). We would also like to thank Dr Osami Nakamura and the other anonymous reviewers for their valuable advice.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Ashford OS, Foster WA, Turner BL, Sayer EJ, Sutcliffe L, Tanner EVJ. 2013. Litter manipulation and the soil arthropod community in a lowland tropical rainforest. Soil Biology & Biochemistry 62:5–12. DOI: 10.1016/j.soilbio.2013.03.001.
- Caterino M, Recuero E. 2023. Molecular diversity of protura in southern high Appalachian leaf litter. Biodiversity Data Journal 11:e113342. DOI: 10.3897/BDJ.11.e113342.
- Galli L. 2022. Updates to Szeptycki’s check-list of the protura of the World. Biogeographia – The Journal of Integrative Biogeography 37(2):a020. DOI: 10.21426/B637258115.
- Galli L, Janžekovič F, Kozel P, Novak T. 2021. Protura (Arthropoda: Hexapoda) in Slovenian caves. International Journal of Speleology 50(1):65‒74. DOI: 10.5038/1827-806X.50.1.2380.
- Galli L, Rellini I. 2020. The geographic distribution of protura (Arthropoda: Hexapoda): A review. Biogeographia – The Journal of Integrative Biogeography 35:51–69. DOI: 10.21426/B635048595.
- Galli L, Shrubovych J, Bu Y, Zinni M. 2018. Genera of the protura of the World: Diagnosis, distribution, and key. ZooKeys 772:1–45. DOI: 10.3897/zookeys.772.24410.
- Nosek J. 1973. The European protura, their taxonomy, ecology and distribution with keys for determination. Muséum d’Histoire Naturelle, Genève. pp. 345.
- Palacios-Vargas JG, Figueroa D. 2014. Biodiversidad de Protura (Hexapoda: Entognatha) en México. Revista Mexicana de Biodiversidad 85:S232–S235. DOI: 10.7550/rmb.32714.
- Román de la Fuente T. 2024. Notas sobre los Protura (Arthropoda: Hexapoda) de Chile: síntesis basada en la literatura científica. Revista chilena de Entomología 50(1):19–26. DOI: 10.35249/rche.50.1.24.03.
- Rusek J, Shrubovych J, Szeptycki A. 2012. Head porotaxy and chaetotaxy of order Acerentomata (Protura). Zootaxa 3262(1):54–61. DOI: 10.11646/zootaxa.3262.1.5.
- Shrubovych J. 2014. Identification and character analysis of the Acerentomidae (Protura) of the northeastern palearctic (Protura: Acerentomidae). Zootaxa 3755(2):136–164. DOI: 10.11646/zootaxa.3755.2.2.
- Shrubovych J, Schneider C, D’Haese C. 2014a. Revision of genus Andinentulus (Protura Acerentomidae: Berberentulinae), with a key to South and Central American Acerentomidae species. Annals of the Entomological Society of America 107(3):567‒574. DOI: 10.1603/AN13151.
- Shrubovych J, Schneider C, D’Haese C. 2014b. Two new species of Acerentulus berlese, 1908 (Protura: Acerentomata: Acerentomidae) with its barcode sequence and a key to the cunhai group. Annales de la Société entomologique de France 50(2):129‒140. DOI: 10.1080/00379271.2014.934036.
- Szeptycki A. 1991. Polish protura V. Genus Acerentulus Berlese, 1908 (Acerentomidae). Acta Zoologica Cracoviensia 34:1–64.
- Szeptycki A. 1993. Three new protura from Western caucasus. Acta Zoologica Cracoviensia 36:29–43.
- Szeptycki A. 2007. Catalogue of the word Protura. Acta Zoologica Cracoviensia 50B(1):1–210.
- Tuxen SL. 1984. Brasilidia and a new genus and species of protura (insecta) from Argentina. Together with an improved generic key to Acerentomidae. Revue d’Écologue et de Biologie du Sol 21(2):283–295.