
Abstract
The development of road networks has given rise to man-made wildlife habitats. Birds of prey use roadsides and highway infrastructure as attractive foraging sites. In order to measure reactions to human disturbance, the flight initiation distance (FID) of Common Buzzards (Buteo buteo) foraging on expressways and in open farmland was investigated. The GLM model showed that FID was affected by habitat types, distance from the nearest building and distance from the nearest tree. Common Buzzards have a shorter FID on expressway than in farmland. The distance from the nearest building and the distance from the nearest tree were positively correlated with FID. Our results confirmed the behavioural plasticity of these birds and may provide a theoretical basis for measures aimed at reducing the risk of collisions with moving road vehicles.
Introduction
The human transformation of wildlife habitats is a widespread and ongoing phenomenon in the modern world (Sanderson et al. Citation2002; Steffen et al. Citation2015; Elhacham et al. Citation2020). One of the most attractive scientific issues that researchers are addressing is the impact of roads on wildlife (Pagany Citation2020). This is due to the development of road networks (especially two-lane expressways and controlled-access highways) expressed in terms of their increased density, area occupied and vehicle traffic. Roads generally have a negative impact on birds, primarily contributing to increased mortality as a result of collisions with vehicles (Fahrig & Rytwinski Citation2009; Benítez-López et al. Citation2010). However, some species of birds can adapt to the disturbed environments near roads, especially to the high level of noise pollution caused by the road traffic. These birds are usually consumers of invertebrates, which provide good foraging in the neighbourhood of roads (Wiącek et al. Citation2015). Birds of prey may also use roadsides and the highway infrastructure as attractive feeding habitats. Especially during poor weather in winter, substantial numbers of buzzards (Kitowski Citation2000; Wuczyński Citation2003), kestrels (Mülner Citation2000; Krasoń & Michalczuk Citation2019) and black kites (Meunier et al. Citation2000) can be seen along roads. The conditions in this type of habitat are favourable for the birds: the thinner snow cover, which makes food more accessible (Sonerud Citation1986; Dobler et al. Citation1991), more opportunities to obtain food in the form of roadkill (e.g. Orłowski & Nowak Citation2006; Borkovcová et al. Citation2012; Garrah et al. Citation2015), and small mammals living on roadsides (Meunier et al. Citation1999; Ruiz-Capillas et al. Citation2013). The road infrastructure also provides plenty of perching sites (Meunier et al. Citation2000; Wuczyński Citation2003; Krasoń & Michalczuk Citation2019), and as birds frequently change them, they can scan a larger area and consequently enhance their hunting success (Kitowski Citation2000).
Flight initiation distance (FID) is a simple behavioural measure of birds’ reactions to the proximity of humans or other predators (Stankowich & Blumstein Citation2005; Cooper & Wilson Citation2007). The decision to initiate escape in response to a disruptive stimulus is determined by a compromise between costs and benefits, where the perceived risk (cost) of an imminent threat becomes greater than the benefits of maintaining regular activity (Cooper & Blumstein Citation2015). FID offers a clear-cut, easily measurable change in animal behaviour (Beale Citation2007) and is likely to vary in cost-benefit assessments across individuals. The different responses to disturbances are due not only to biological differences among individuals, but also to the properties of the disturbance itself and the environment in which the interaction takes place. For example, the FID of a particular species relative to a human is shorter in urban than in rural areas (Møller Citation2008b; Bjørvik et al. Citation2015; Samia et al. Citation2015, Citation2017; Morelli et al. Citation2022) – this may be mediated by bird feeders (Møller et al. Citation2015) and cold winters (Møller et al. Citation2013). In some species, FID in response to vehicles has also been shown to be shorter than in response to pedestrians (McLeod et al. Citation2013), and the speed of vehicles is negatively correlated with the flight distance of birds (Legagneux & Ducatez Citation2013).
FID is closely correlated with synanthropization, while the proximity of humans as well as the various types of environmental disturbances they cause shorten the escape distance (Møller Citation2008b; McLeod et al. Citation2013; Bjørvik et al. Citation2015; Samia et al. Citation2015, Citation2017; Morelli et al. Citation2022). The aim of our research was to determine factors affecting the FID of wintering (non-breeding) Common Buzzards Buteo buteo (henceforth: Buzzard(s)) in the vicinity of expressways and in open farmland. This is the most common nesting bird of prey in Poland (Chylarecki et al. Citation2018), and its breeding season of a single brood begins in April (Cramp & Simmons Citation1980). During the wintering season, the Buzzard is the dominant species in the raptor assemblage in east-central Poland (Kasprzykowski & Rzępała Citation2002). It is a sturdily built, medium-sized to large diurnal bird of prey, with a total length of about 51–57 cm and a wingspan of 113–128 cm. Adults typically weigh 800–1200 g, while females are heavier than males by 20–30% on average (Cramp & Simmons Citation1980). In winter, Common Buzzards feed on small mammals, mainly Common Voles Microtus arvalis (Kowalski & Rzępała Citation1997).
Our hypothesis is that the FID of Buzzards depends on the habitat type. We anticipate that birds foraging on expressways will have a shorter FID than birds in farmland and that the distance from the hunting site to the nearest building and nearest tree will govern the initiation of escape. We also take into account the starting distance (SD), which is the initial distance between the bird’s position and that of the observer (Blumstein Citation2010). We believe that comparative research on the FID for this raptor could provide an answer regarding the plasticity of Buzzard behaviour in response to anthropogenic environmental disturbances. It will also be the basis for planning measures aimed at reducing the risk of vehicle collisions with birds of prey, which could have potentially catastrophic consequences for road users.
Material and methods
Study area
The research was carried out in north-eastern Poland, in two types of habitat characterized by different levels of human impact. The former areas were situated in the immediate vicinity of expressway S8 between the towns of Wyszków and Zambrów, a distance of about 60 km. The studied section of the expressway was a two-lane dual carriageway. Between the carriageways, each 10.5 m wide, there was a central reservation 12 m in width. The expressway was bounded along each side by a 2.5 m tall wire mesh fence supported on metal posts inserted every 3 m. There was also a single-lane service road running parallel to the expressway, along which the observer moved. In 2021, an average of 26 439 vehicles moved along the surveyed section of the expressway per day (GDDKiA Citation2021). The COVID-19 pandemic presumably had only a limited impact on the volume of motor vehicle traffic on the studied section of the S8. Indirect evidence for this is the average increase of 27% in vehicle traffic on international roads in Poland in 2021 compared to 2015 (GDDKiA Citation2021).
Along the S8, Buzzards perched on the wire-netting fence posts, also on road signs and lamp-posts, which were most often located near the eight road junctions. The latter areas were located in the open agricultural landscape up to about 15 km from that expressway, and covered a total area of about 280 km2. The study area is a mosaic of meadows, pastures and arable land with isolated trees and shrubs. The agricultural landscape here lies almost entirely within the Biała Forest, a fragmented forest complex dominated by pine monocultures.
Data collection
Observations in the two habitat types were conducted from early November to early March in the winter seasons of 2020/2021 and 2021/2022. This period is best suited to recording wintering birds because the number of buzzards increases in the second half of October as a result of their autumn migration, whereas the spring migration, taking place between late February and early March, is less pronounced (Tomiałojć & Stawarczyk Citation2003). Buzzards were observed at regular, approximately weekly intervals on each habitat type. In 2020/2021, FID was tested in 139 birds (80 by the roadside and 59 on farmland), and in 2021/2022 in 132 birds (76 by the roadside and 56 on farmland). FIDs were always recorded during the same part of the day, i.e. from 08:00 to 14:00 hrs. Buzzards were not individually marked, so multiple observations of the same individual were unavoidable. Nevertheless, all the records were treated equally, i.e. each recorded individual was counted separately (see also Peck et al. Citation2014; Rothery et al. Citation2017; Golawski & Sytykiewcz Citation2021). In addition, individuals from eastern European populations may appear during the winter in north-eastern Poland, especially when the weather conditions in their nominal wintering grounds deteriorate (Kasprzykowski & Rzępała Citation2002; Wuczyński Citation2003; Baltag et al. Citation2018). Waves of such individuals arriving from the east, which mix with birds wintering locally, may also reinforce the independence of observations.
FID was measured in that randomly selected buzzards were approached using a modified version of the technique developed by Blumstein (Citation2006). During the approach, the observer counted paces (of equal length, about 0.8 m) at a steady speed, approximately 1 pace/second. The distances measured in paces were converted to metres. The distance from the observer to the bird when it first took flight was recorded as the FID, while the starting distance SD was the distance between the point from where the observer started walking towards the bird and its position. If the bird was perching above ground, the height of its perch was also recorded to the nearest metre. Birds on the ground were assigned a height of 0. Together with FID, the following parameters were also recorded: date, time, place of observation (including the coordinates of the point from which the bird took off) and habitat type. The distance from the hunting site to the nearest building and the nearest tree was determined from geographical coordinates using QGIS software tools. FID was calculated as the square root of the sum of the squared horizontal distance and the squared height (Blumstein Citation2006).
Statistical analyses
A general linear model (GLM) with gamma distribution and log link function was used to find the effect of habitat types, SD, distance from the nearest building and distance from the nearest tree on Buzzard FID (). The models were constructed using the glm function in the lme4 package for R (Bates et al. Citation2015). The two habitat types – road and farmland – were treated as categorical factors. SD, the distance from the nearest building and the distance from the nearest tree were treated as continuous predictors and log-transformed. FID was the continuous dependent variable. An information-theoretic approach (AIC) was used to compare the competing models (Burnham & Anderson Citation2002) and to analyse model support using the AIC value, which corrects for small sample sizes and evaluates the strength of evidence for each model using normalized weights (AICwt). The models selected with the smallest AIC as being the best of all the models were compared; only the models within a ∆AIC of < 5.00 were taken into consideration. All data were analysed in the R environment (R Core Team Citation2023). The values reported are the mean ± Cl. Only those results with a probability of α ≤ 0.05 were assumed to be statistically significant.
Table I. Factors for analysing effects on Common Buzzard flight initiation distance.
Results
A total of 271 FIDs were recorded for Buzzards in their foraging areas. Most often, the birds initiated flight at FIDs between 100 and 200 m (); the mean FID was 203.30 m (SD = 107.55 m, min = 44 m, max = 528 m). Models containing four predictors were created on the basis of Akaike’s information criteria (). The best model of the 16 a priori models with the highest ranking received 57% support (sum of AICcwt, ) and included all the factors analysed: habitat types, SD, distance from the nearest building and distance from the nearest tree. The GLM model showed that three parameters had a significant influence on FID (). Buzzards took flight from a shorter distance on the expressway than in farmland (). The difference in the mean distance between these categories was 219.7 m, a figure less than half of that on sites situated close to the expressway. Distance from the nearest building and distance from the nearest tree were positively correlated with FID. SD did not predict Buzzard escape responses to an approaching person ().
Figure 1. Data distribution of flight initiation distance (m) of Common Buzzards Buteo buteo.
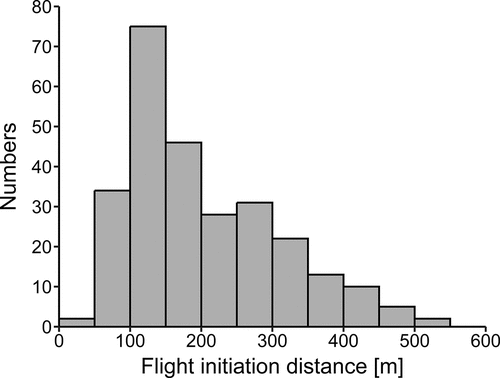
Figure 2. Mean number (dots), 95% confidence limits (vertical lines) and kernel distribution (violins) of recorded flight initiation distances in two types of habitats.
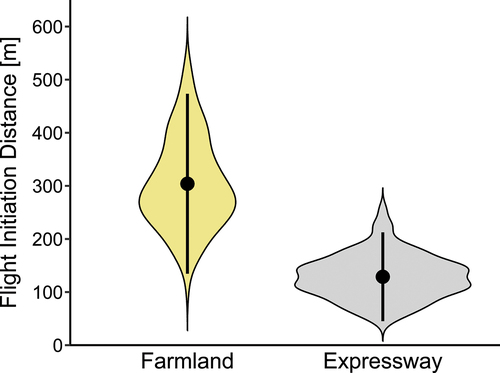
Table II. Results of models describing the influence of different factors on Common Buzzard flight initiation distance. Degrees of freedom (df), model log-likelihood (LL), corrected AIC criterion (AIC), difference between the model and the best model in the data set (Δ AIC), and weight for the model (AICwt) are shown.
Table III. Estimates of GLM model coefficients for the best model of factors affecting FID.
Discussion
Our study showed that the mean FID was at least several times greater than that of other accipitrids with a body mass comparable to that of Buzzards during the non-breeding season (Livezey et al. Citation2016). However, it is hard to draw unequivocal conclusions from this fact, because most of the studies that we compared ours to were based on a much smaller research sample. In the case of six Australian accipitrid species, the samples consisted of only a dozen or so birds, and for four of these species only single results were obtained (Weston et al. Citation2012). The same applies to Europe, where the mean FID for Buzzards was more than four and a half times shorter, a result based on a sample more than 30 times smaller than ours (Møller Citation2008a). Larger research samples were obtained in the case of Rough-legged Buzzard Buteo lagopus in North America, and here the mean FID was the closest to our results (Holmes et al. Citation1993). Another factor affecting the results of other researchers could have been the magnitude of environmental disturbances, e.g. the density or presence of passers-by (Weston et al. Citation2012; McGiffin et al. Citation2013), or even the possibility of a bird managing to learn to recognize particular persons (Levey et al. Citation2009).
We also found that the FID of Buzzards in localities close to expressways was less than half the figure for farmland. The explanation for this difference should be sought in the nature of the environment immediately adjoining busy roads. This environment is highly disturbed by human activities: intensive vehicle traffic, which is a source of noise, and the drastically transformed landscape with wide stretches of tarred road, embankments and drainage ditches, all enclosed on either side by a wire-netting fence. Such features of this environment can indirectly lead to a reduction in FID, because the birds perceive them as barriers to the free movements of pedestrians, who are then potentially less of a threat (Weston et al. Citation2012). Habitats associated with road infrastructure are also attractive foraging areas, where the ready availability of lookout posts means that energy can be saved during hunting, an aspect particularly important for maintaining the energy balance in winter (Meunier et al. Citation2000). The economic theory of escape postulates that by delaying their escape flight after having perceived a potential threat, birds can devote more of their limited energy resources to other key behaviours, such as foraging (Ydenberg & Dill Citation1986; Cooper & Frederick Citation2007). The reduced FID therefore reflects a compromise on the part of the bird between the advantages of remaining in situ in the form of time devoted to foraging and saving energy, and the costs of the risk of predation and death (Cooper & Blumstein Citation2015). So if another lookout post is available nearby, a Buzzard will delay taking off on seeing a pedestrian approaching, thus saving the maximum amount of energy. The modification of escape behaviours in Buzzards may also be due to the heavy traffic on the road and the consequent high level of noise. Where an environment is subject to strong human pressure, e.g. in urban areas, where conditions may be similar to those described above, birds generally have shorter average FIDs (Møller Citation2008b; Bjørvik et al. Citation2015; Samia et al. Citation2015, Citation2017; Morelli et al. Citation2022), especially during frosty winters (Møller et al. Citation2013).
This study demonstrated a positive link between Buzzard FID and the distance to the nearest building. This may be evidence of individual habituation and greater tolerance among Buzzards of the presence of people, i.e. phenotypic plasticity (Blumstein Citation2016), owing to the heightened activities of people around such artificial components of the landscape like barns, houses and farm buildings. Similar results were obtained in other bird species, where FIDs on perceiving a human were shorter in anthropogenically more disturbed areas (Møller Citation2008b; McGiffin et al. Citation2013; Bjørvik et al. Citation2015; Samia et al. Citation2015, Citation2017; Tryjanowski et al. Citation2020; Morelli et al. Citation2022). Changes in escape behaviour may be justified by the possibility of strong selection against timid individuals (Carrete et al. Citation2016) or the recruitment of less sensitive individuals in more disturbed environments (Weston et al. Citation2012; Carrete & Tella Citation2013). In addition, this kind of environmental disturbance may be a source of readily available food and encourage the birds to approach buildings, particularly when snowy winters make finding food difficult (Sonerud Citation1986; Dobler et al. Citation1991). In such instances, FID in many other bird species may also fall when a human being comes into sight (Møller et al. Citation2013, Citation2015). That such a situation may occur among Buzzards is indicated by their opportunistic manner of feeding (Oro et al. Citation2021) and the fact that in winter, their diet may contain domestic fowl or other synanthropic bird species (Goszczyński et al. Citation2005). At such times, one observes an apparent increase in the Buzzards’ tolerance towards disturbances, resulting from the limited availability of food in places distant from human habitations (Owens Citation1977).
The second positive association, between FID and distance to the nearest tree, can be interpreted in the case of Buzzards with respect to trees as sites providing protection and trees as lookout posts. The link between the escape distance and the distance to the nearest shelter has been widely described in the literature for a variety of animals, including fish, reptiles and mammals. As in our study, escape among various species of vertebrates was initiated the sooner, the greater the distance to a shelter (Dill & Houtman Citation1989; Dill Citation1990; Bonenfant & Kramer Citation1996; Cooper Citation1997, Citation2007; Cooper & Wilson Citation2007). The same relationship has been found in birds. In support of the predictions of the optimal escape theory (Ydenberg & Dill Citation1986; Cooper & Frederick Citation2007), Morelli et al. (Citation2022) demonstrated that the FID of birds in urban greenery increases with distance to the nearest accessible shelter. This suggests that birds also base their decisions to escape on the relative time needed to reach the nearest shelter, which in turn affects the assessment of the risk of danger from a potential predator. Hence, Buzzards may be more afraid of a pedestrian when they are farther away from the nearest tree. On the other hand, the shorter FID of Buzzards when a tree (lookout post) is closer to hand, can be interpreted as the consequence of saving energy and taking full advantage of the time available for foraging (Ydenberg & Dill Citation1986; Cooper & Frederick Citation2007). Maintaining the proper energy balance is absolutely crucial in winter. The lack of food or its limited availability in combination with inclement winter weather can lead to substantial energy losses in animals and may be the cause of their mortality (Sonerud Citation1986; Schmidt-Nielsen Citation1997; Mülner Citation2000; Blix Citation2016). Under such conditions, Buzzards, being able to fly to a nearby “safe haven”, i.e. another tree, probably postpone the decision to escape.
In the Buzzards that we studied, SD did not affect the FID. However, most other studies do not corroborate this finding (Blumstein Citation2003, Citation2010; Mikula et al. Citation2018, Citation2022; Morelli et al. Citation2022). Nonetheless, when analysing the behavioural decisions of birds, it is very important to take their context into consideration (Beale Citation2007). Therefore, the lack of a correlation between SD and FID in the Buzzards that we studied may be due to factors that hamper continuous monitoring by the observer approaching them and thus modify their escape behaviour (Blumstein Citation2010; Samia et al. Citation2017). Not to be overlooked is also the fact that some of the Buzzards in the population we studied are recruited from eastern Europe (Kasprzykowski & Rzępała Citation2002; Wuczyński Citation2003; Baltag et al. Citation2018), where they will have been inhabiting areas less densely populated by humans and less disturbed by human activities, which could have a bearing on their escape behaviours.
Conclusion
The present study has shown that the readiness of Common Buzzards to take a risk is mediated by the place where they happen to be (expressway vs. farmland), and also by the distances to the nearest building and to the nearest tree. Our results confirmed the behavioural plasticity of these birds and may provide a theoretical basis for measures aimed at reducing the risk of collisions with moving road vehicles.
Ethical standards
The authors declare that this publication complies with the current laws of the Ethical Committee.
Acknowledgments
We thank Peter Senn for the English language editing and Przemysław Obłoza for help in statistical analyses.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
Dataset supporting reported results can be found at the Mendeley Data Repository: https://doi.org/10.17632/k7jwk6pgpc.1
Additional information
Funding
References
- Baltag EŞ, Petrencu L, Bolboacă LE, Sfîcă L. 2018. Common Buzzards Buteo buteo wintering in Eastern Romania: Habitat use and climatic factors affecting their abundance. Acta Ornithologica 53(1):1–12. DOI: 10.3161/00016454AO2018.53.1.001.
- Bates D, Mächler M, Bolker B, Walker S. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1):1–48. DOI: 10.18637/jss.v067.i01.
- Beale CM. 2007. The behavioral ecology of disturbance responses. International Journal of Comparative Psychology 20(2): 111–120. DOI: 10.46867/ijcp.2007.20.02.13.
- Benítez-López A, Alkemade R, Verweij PA. 2010. The impacts of roads and other infrastructure on mammal and bird populations: A meta-analysis. Biological Conservation 143(6): 1307–1316. DOI: 10.1016/j.biocon.2010.02.009.
- Bjørvik LM, Dale S, Hermansen GH, Munishi PK, Moe SR. 2015. Bird flight initiation distances in relation to distance from human settlements in a Tanzanian floodplain habitat. Journal of Ornithology 156(1):239–246. DOI: 10.1007/s10336-014-1121-1.
- Blix AS. 2016. Adaptations to polar life in mammals and birds. Journal of Experimental Biology 219(8): 1093–1105. DOI: 10.1242/jeb.120477.
- Blumstein DT. 2003. Flight initiation distance in birds is dependent on intruder starting distance. The Journal of Wildlife Management 67(4):852–857. DOI: 10.2307/3802692.
- Blumstein DT. 2006. Developing an evolutionary ecology of fear: How life history and natural history traits affect disturbance tolerance in birds. Animal Behaviour 71(2):389–399. DOI: 10.1016/j.anbehav.2005.05.010.
- Blumstein DT. 2010. Flush early and avoid the rush: A general rule of anti-predator behavior? Behavioural Ecology 21(3):440–442. DOI: 10.1093/beheco/arq030.
- Blumstein DT. 2016. Habituation and sensitization: New thoughts about old ideas. Animal Behaviour 120:255–262. DOI: 10.1016/j.anbehav.2016.05.012.
- Bonenfant M, Kramer DL. 1996. The influence of distance to burrow on flight initiation distance in the woodchuck, Marmota monax. Behavioral Ecology 7(3):299–303. DOI: 10.1093/beheco/7.3.299.
- Borkovcová M, Mrtka J, Winkler J. 2012. Factors affecting mortality of vertebrates on the roads in the Czech Republic. Transportation Research Part D: Transport & Environment 17(1):66–72. DOI: 10.1016/j.trd.2011.09.011.
- Burnham KP, Anderson DR. 2002. Model selection and multimodel inference: A practical information-theoretic approach. New York: Springer-Verlag.
- Carrete M, Martínez-Padilla J, Rodríguez-Martínez S, Rebolo-Ifrán N, Palma A, Tella JL. 2016. Heritability of fear of humans in urban and rural populations of a bird species. Scientific Reports 6(1): 31060. DOI: 10.1038/srep31060.
- Carrete M, Tella JL. 2013. High individual consistency in fear of humans throughout the adult lifespan of rural and urban burrowing owls. Scientific Reports 3(1): 3524. DOI: 10.1038/srep03524.
- Chylarecki P, Chodkiewicz T, Neubauer G, Sikora A, Meissner W, Woźniak B, Wylegała P, Ławicki Ł, Marchowski D, Betleja J, Bzoma S, Cenian Z, Górski A, Korniluk M, Moczarska J, Ochocińska D, Rubacha S, Wieloch M, Zielińska M, Zieliński P, Kuczyński L. 2018. Trendy Liczebności Ptaków w Polsce. Warszawa: Główny Inspektorat Ochrony Środowiska. In Polish.
- Cooper Jr. WE. 1997. Escape by a refuging prey, the broad-headed skink (Eumeces laticeps). Canadian Journal of Zoology 75(6): 943–947. DOI: 10.1139/z97-113.
- Cooper Jr. WE. 2007. Escape and its relationship to pursuit-deterrent signalling in the Cuban curly-tailed lizard Leiocephalus carinatus. Herpetologica 63(2):144–150. DOI: 10.1655/0018-0831(2007)63[144:EAIRTP]2.0.CO;2.
- Cooper Jr WE, Blumstein DT, eds. 2015. Escaping from predators: An integrative view of escape decisions. Cambridge: Cambridge University Press.
- Cooper Jr WE, Frederick WG. 2007. Optimal time to emerge from refuge. Biological Journal of the Linnean Society 91(3):375–382. DOI: 10.1111/j.1095-8312.2007.00802.x.
- Cooper Jr WE, Wilson DS. 2007. Beyond optimal escape theory: Microhabitats as well as predation risk affect escape and refuge use by the phrynosomatid lizard Sceloporus virgatus. Behaviour 144(10): 1235–1254. DOI: 10.1163/156853907781890940.
- Cramp S, Simmons KEL, eds. 1980. Handbook of the birds of Europe, Middle East and North Africa: The birds of the Western Palearctic. Vol. 2. Oxford: Oxford University Press.
- Dill LM. 1990. Distance-to-cover and the escape decisions of an African cichlid fish, Melanochromis chipokae. Environmental Biology of Fishes 27(2):147–152. DOI: 10.1007/BF00001944.
- Dill LM, Houtman R. 1989. The influence of distance to refuge on flight initiation distance in the gray squirrel (Sciurus carolinensis). Canadian Journal of Zoology 67(1):233–235. DOI: 10.1139/z89-033.
- Dobler G, Schneider R, Schweis A. 1991. Influx of Rough-legged Buzzards (Buteo lagopus) into south-western Germany (Baden-Wurttemberg) in the winter 1986/87. Vogelwarte 36:1–18. In German with English summary.
- Elhacham E, Ben-Uri L, Grozovski J, Bar-On YM, Milo R. 2020. Global human-made mass exceeds all living biomass. Nature 588(7838):442–444. DOI: 10.1038/s41586-020-3010-5.
- Fahrig L, Rytwinski T. 2009. Effects of roads on animal abundance: An empirical review and synthesis. Ecology and Society 14(1):21. http://www.ecologyandsociety.org/vol14/iss1/art21/.
- Garrah E, Danby RK, Eberhardt E, Cunnington GM, Mitchell S. 2015. Hot spots and hot times: Wildlife road mortality in a regional conservation corridor. Environmental Management 56(4):874–889. DOI: 10.1007/s00267-015-0566-1.
- Generalna Dyrekcja Dróg Krajowych i Autostrad. 2021. Synteza wyników GPR 2020/21 na zamiejskiej sieci dróg krajowych. Available: https://www.gov.pl/web/gddkia/generalny-pomiar-ruchu-20202021. Accessed Feb 2024 10.
- Golawski A, Sytykiewcz H. 2021. How urban and rural birds respond to the colour of bird feeders? Journal of Ornithology 162(4):1193–1198. DOI: 10.1007/s10336-021-01907-8.
- Goszczyński J, Gryz J, Krauze D. 2005. Fluctuations of a Common Buzzard Buteo buteo population in Central Poland. Acta Ornithologica 40(1):75–78. DOI: 10.3161/068.040.0102.
- Holmes TL, Knight RL, Stegall L, Craig GR. 1993. Responses of wintering grassland raptors to human disturbance. Wildlife Society Bulletin 21(4):461–468. https://www.jstor.org/stable/3783420.
- Kasprzykowski Z, Rzępała M. 2002. Numbers and habitat preferences of raptors Falconiformes wintering in central-eastern Poland. Notatki Ornitologiczne 43:73–82. In Polish with English summary.
- Kitowski I. 2000. Numbers and behaviour of the Common Buzzard Buteo buteo in the post-breeding season and early spring in the agricultural landscape of the central Lublin region. Notatki Ornitologiczne 41:265–272. In Polish with English summary.
- Kowalski M, Rzępała M. 1997. Diet of the Common Buzzard Buteo buteo and Rough-legged Buzzard Buteo lagopus in the autumn-winter season. Notatki Ornitologiczne 38:121–130. In Polish with English summary.
- Krasoń K, Michalczuk J. 2019. Numbers, hunting behaviour and feeding preferences of birds of prey (Accipitriformes and Falconiformes) wintering in agricultural landscape in the Kotlina Sandomierska. Ornis Polonica 60:269–284. In Polish with English summary.
- Legagneux P, Ducatez S. 2013. European birds adjust their flight initiation distance to road speed limits. Biology Letters 9(5): 20130417. DOI: 10.1098/rsbl.2013.0417.
- Levey DJ, Londoño GA, Ungvari-Martin J, Hiersoux MR, Jankowski JE, Poulsen JR, Stracey CM, Robinson SK. 2009. Urban mockingbirds quickly learn to identify individual humans. Proceedings of the National Academy of Sciences USA 106(22): 8959–8962. DOI: 10.1073/pnas.0811422106.
- Livezey KB, Fernandez-Juricic E, Blumstein DT. 2016. Database of bird flight initiation distances to assist in estimating effects from human disturbance and delineating buffer areas. Journal of Fish and Wildlife Management 7(1):181–191. DOI: 10.3996/082015-JFWM-078.
- McGiffin A, Lill A, Beckman J, Johnstone CP. 2013. Tolerance of human approaches by Common Mynas along an urban-rural gradient. Emu-Austral Ornithology 113(2):154–160. DOI: 10.1071/MU12107.
- McLeod EM, Guay PJ, Taysom AJ, Robinson RW, Weston MA, Sueur C. 2013. Buses, cars, bicycles and walkers: The influence of the type of human transport on the flight responses of waterbirds. PLOS ONE 8(12):e82008. DOI: 10.1371/journal.pone.0082008.
- Meunier FD, Corbin J, Verheyden C, Jouventin P. 1999. Effects of landscape type and extensive management on use of motorway roadsides by small mammals. Canadian Journal of Zoology 77(1): 108–117. DOI: 10.1139/z98-203.
- Meunier FD, Verheyden C, Jouventin P. 2000. Use of roadsides by diurnal raptors in agricultural landscapes. Biological Conservation 92(3): 291–298. DOI: 10.1016/S0006-3207(99)00094-4.
- Mikula P, Díaz M, Albrecht T, Jokimäki J, Kaisanlahti-Jokimäki M-L, Kroitero G, Møller AP, Tryjanowski P, Yosef R, Hromada M. 2018. Adjusting risk-taking to the annual cycle of long-distance migratory birds. Scientific Reports 8(1):13989. DOI: 10.1038/s41598-018-32252-1.
- Mikula P, Kwieciński Z, Kaługa I, Tryjanowski P. 2022. The Crimean population of the lesser grey shrike (Lanius minor) has low behavioural flexibility in its response to approaching humans. Journal of Vertebrate Biology 71(22038):22038. DOI: 10.25225/jvb.22038.
- Møller AP. 2008a. Flight distance and blood parasites in birds. Behavioral Ecology 19(6):1305–1313. DOI: 10.1093/beheco/arn074.
- Møller AP. 2008b. Flight distance of urban birds, predation, and selection for urban life. Behavioral Ecology and Sociobiology 63(1): 63–75. DOI: 10.1007/s00265-008-0636-y.
- Møller AP, Grim T, Ibáñez-Álamo JD, Markó G, Tryjanowski P. 2013. Change in flight initiation distance between urban and rural habitats following a cold winter. Behavioral Ecology 24(5): 1211–1217. DOI: 10.1093/beheco/art054.
- Møller AP, Tryjanowski P, Díaz M, Kwieciński Z, Indykiewicz P, Mitrus C, Goławski A, Polakowski M. 2015. Urban habitats and feeders both contribute to flight initiation distance reduction in birds. Behavioral Ecology 26(3): 861–865. DOI: 10.1093/beheco/arv024.
- Morelli F, Mikula P, Blumstein DT, Díaz M, Markó G, Jokimäki J, Kaisanlahti-Jokimäki M-L, Floigl K, Abou Zeid F, Siretckaia A, Benedetti Y. 2022. Flight initiation distance and refuge in urban birds. Science of the Total Environment 842:156939. DOI: 10.1016/j.scitotenv.2022.156939.
- Mülner B. 2000. Population numbers, habitat preferences and perch use of wintering Common Buzzards (Buteo buteo) and Common Kestrel (Falco tinnunculus) in the upper Mur river valley (Styria). Egretta 43:20–36. In German with English summary.
- Orłowski G, Nowak L. 2006. Factors influencing mammal roadkills in the agricultural landscape of south-western Poland. Polish Journal of Ecology 54(2):283–294.
- Oro D, Sanz-Aguilar A, Carbonell F, Grajera J, Torre I. 2021. Multi-species prey dynamics influence local survival in resident and wintering generalist predators. Oecologia 197(2): 437–446. DOI: 10.1007/s00442-021-05042-2.
- Owens NW. 1977. Responses of wintering brent geese to human disturbance. Wildfowl 28:5–14. https://wildfowl.wwt.org.uk/index.php/wildfowl/article/view/540/540.
- Pagany R. 2020. Wildlife-vehicle collisions-Influencing factors, data collection and research methods. Biological Conservation 251:108758. DOI: 10.1016/j.biocon.2020.108758.
- Peck HL, Pringle HE, Marshall HH, Owens IPF, Lord AM. 2014. Experimental evidence of impacts of an invasive parakeet on foraging behavior of native birds. Behavioral Ecology 25(3):582–590. DOI: 10.1093/beheco/aru025.
- R Core Team. 2023. R: A Language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Available: https://www.R-project.org/. Accessed Apr 2023 22.
- Rothery L, Scott GW, Morrell LJ, Dyer AG. 2017. Colour preferences of UK garden birds at supplementary seed feeders. PLOS ONE 12(2):e0172422. DOI: 10.1371/journal.pone.0172422.
- Ruiz-Capillas P, Mata C, Malo JE. 2013. Road verges are refuges for small mammal populations in extensively managed Mediterranean landscapes. Biological Conservation 158:223–229. DOI: 10.1016/j.biocon.2012.09.025.
- Samia DSM, Blumstein DT, Diaz M, Grim T, Ibáñez-Álamo JD, Jokimäki J, Tätte K, Marko G, Tryjanowski P, Møller AP. 2017. Rural-urban differences in escape behavior of European birds across a latitudinal gradient. Frontiers in Ecology and Evolution 5:66. DOI: 10.3389/fevo.2017.00066.
- Samia DSM, Nakagawa S, Nomura F, Rangel TF, Blumstein DT. 2015. Increased tolerance to humans among disturbed wildlife. Nature Communications 6(1):8877. DOI: 10.1038/ncomms9877.
- Sanderson EW, Jaiteh M, Levy MA, Redford KH, Wannebo AV, Woolmer G. 2002. The human footprint and the last of the wild: The human footprint is a global map of human influence on the land surface, which suggests that human beings are stewards of nature, whether we like it or not. BioScience 52(10):891–904. DOI: 10.1641/0006-3568(2002)052[0891:THFATL]2.0.CO;2.
- Schmidt-Nielsen K. 1997. Animal physiology: Adaptation and environment. Fifth ed. Cambridge: Cambridge University Press.
- Sonerud GA. 1986. Effect of snow cover on seasonal changes in diet, habitat, and regional distribution of raptors that prey on small mammals in boreal zones of Fennoscandia. Holarctic Ecology 9(1):33–47. http://www.jstor.org/stable/3682109?origin=JSTOR-pdf.
- Stankowich T, Blumstein DT. 2005. Fear in animals: A meta-analysis and review of risk assessment. Proceedings of the Royal Society of London B, Biological Sciences 272(1581):2627–2634. DOI: 10.1098/rspb.2005.3251.
- Steffen W, Broadgate W, Deutsch L, Gaffney O, Ludwig C. 2015. The trajectory of the Anthropocene: The great acceleration. The Anthropocene Review 2(1): 81–98. DOI: 10.1177/2053019614564785.
- Tomiałojć L, Stawarczyk T. 2003. The avifauna of Poland. Distribution, numbers and trends. Wrocław: PTPP “proNatura”. In Polish with English summaries.
- Tryjanowski P, Kosicki JZ, Hromada M, Mikula P. 2020. The emergence of tolerance of human disturbance in neotropical birds. Journal of Tropical Ecology 36(1): 1–5. DOI: 10.1017/S0266467419000282.
- Weston MA, McLeod EM, Blumstein DT, Guay P-J. 2012. A review of flight-initiation distances and their application to managing disturbance to Australian birds. Emu-Austral Ornithology 112(4): 269–286. DOI: 10.1071/MU12026.
- Wiącek J, Polak M, Kucharczyk M, Bohatkiewicz J. 2015. The influence of road traffic on birds during autumn period: Implications for planning and management of road network. Landscape and Urban Planning 134:76–82. DOI: 10.1016/j.landurbplan.2014.10.016.
- Wuczyński A. 2003. Abundance of Common Buzzard (Buteo buteo) in the Central European wintering ground in relation to the weather conditions and food supply. Buteo 13:11–20.
- Ydenberg RC, Dill LM. 1986. The economics of fleeing from predators. Advances in the Study of Behavior 16:229–249. DOI: 10.1016/S0065-3454(08)60192-8.