
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
The Northern Periphery and Arctic region faces unique economic and food security challenges that may be partly answered by commercial seaweed production. Alaria esculenta and Palmaria palmata are two seaweeds commonly found in the region and suitable for cultivation and processing for food and other commercial products. The drying kinetics for both species were obtained, and the Page and Weibull models best described the data. A drying air temperature increase from 40 to 70°C decreased drying time by 62.4% and 61.7% for A. esculenta and P. palmata, respectively. Desorption isotherms were obtained between 25 and 70°C and showed Brunauer Category III shapes, with water activity increasing with temperature for a fixed moisture content. Net heats of desorption were obtained, with drying to an equilibrium moisture content of 0.01 kgwater kgd.b.–1 requiring 18.1 and 3.94 kJ mol–1 K–1 for A. esculenta and P. palmata, respectively.
Introduction
The Northern Periphery and Arctic (NPA) region, although geographically diverse, shares several challenges including low accessibility, low economic diversity and high potential impact of climate change (Northern Periphery and Arctic Programme, Citation2016). Population growth and climate change have the potential to exacerbate these issues and raise food security concerns. Additionally, growing awareness of the carbon footprint of foods raises calls for food produced locally, efficiently and sustainably. The NPA Programme aims to overcome such challenges by transnational project cooperation involving nine partner countries in the region.
Seaweed production may form part of the solution to these challenges. Seaweed cultivation has no land area demand, and a processing facility needs only a small land footprint; there is thus no competition with agriculture for viable grazing or arable land, which is scarce in many parts of the NPA region (Jacobson, Citation2016; Kintisch, Citation2016). Seaweed cultivation and processing offers an alternative or complementary livelihood to fishing using many of the same skills and not requiring major retraining.
SW-GROW is a multidisciplinary project funded by the NPA that aims to innovate and communicate research and best practice to bolster the seaweed industry in the region. The SW-GROW programme identified two seaweed species as of special interest for food cultivation: Alaria esculenta and Palmaria palmata. A. esculenta (winged kelp, dabberlocks or lair) is a brown seaweed found at low tide and in the sublittoral zone across the NPA region (Springer, Lütz, Lütz-Meindl, Wendt, & Bischof, Citation2017). Its regional limits are determined by the 16°C summer water temperature isotherm (Lüning, Citation1990); with warming seawater, its southern range is expected to contract (Mieszkowska et al., Citation2006); however, its abundance across the British Isles has not changed significantly between 1974 and 2010 (Yesson, Bush, Davies, Maggs, & Brodie, Citation2015). It is relatively tolerant of salinity variation (Fredersdorf, Müller, Becker, Wiencke, & Bischof, Citation2009). It has been used as a food for humans (Chapman, Stévant, & Larssen, Citation2015) and land (Seterlund, Hoie, Sannan, & Raastad, Citation1968) and sea (Mai, Mercer, & Donlon, Citation1994) dwelling herbivores, and contains viable quantities of sugars, proteins and minerals (Schiener, Black, Stanley, & Green, Citation2015). It can also be used for nutrient sequestration in aquaculture (Reid et al., Citation2013) and in the production of biocoal and biomethane by hydrothermal carbonization (Smith & Ross, Citation2016). To date various seeded rope methods have been investigated for commercial cultivation of A. esculenta (Kerrison et al., Citation2020).
P. palmata (dulse, dillisk, söl) is a red seaweed found in the littoral and sublittoral zones across the NPA region (Werner & Dring, Citation2011); growth rate deterioration begins between 14°C and 18°C summer temperature (Morgan & Simpson, Citation1981), and salinity. It is amongst the most commonly used seaweeds for human consumption – in the UK and Ireland, Norway, the Faroes, Iceland and New England – due partly to its attractive taste profile, and is relatively high in amino acid and lipid content (Mouritsen et al., Citation2013). Additionally, the long-chain lipids in P. palmata are more readily digestible to humans than those found in other seaweeds (Lopes et al., Citation2019; van Ginneken, Helsper, de Visser, van Keulen, & Brandenburg, Citation2011) and can be further enhanced by heat treatment (Maehre, Edvinsen, Eilertsen, & Elvevoll, Citation2016). Alongside food use, P. palmata supplements have may have health benefits including hypertriglyceridaemia reduction (Takase et al., Citation2020) and hypertension and Type II diabetes control (Harnedy & FitzGerald, Citation2013).
Both seaweed species can be eaten fresh, but, because fresh seaweed is highly perishable, dehydration is normally required for commercial distribution and in order to preserve the seaweed for future consumption. Drying is an energy-intensive step in seaweed processing, and the objectives are to maximise nutritional characteristics whilst preventing microbial growth, at the minimum energy cost. Microbial growth is promoted with increasing water activity but cannot occur where aw < 0.6 (Fontana, Citation2020). Al-Muhtaseb, McMinn, and Magee (Citation2002) provide a succinct review of water activity isotherm modelling. Historically, open air drying was common but convective air drying methods are common today; principally because they are relatively simple to design and effectively preserve nutrients in the dried seaweed (Badmus, Taggart, & Boyd, Citation2019; Uribe et al., Citation2019). Erbay and Icier (Citation2010) and Onwude et al. (Citation2016) present thorough reviews and classification of convective drying models. This paper provides the drying kinetics, desorption isotherms and isosteric desorption heats of A. esculenta and P. palmata. Additionally, it enables seaweed producers to design and calibrate convection drying systems in which convection is either natural or the air velocity is low. We expect to report the effects of air velocity, temperature and humidity in forced convective drying systems in a future paper.
Materials and methods
Sample preparation
Both seaweed species were wild harvested: A. esculenta from the coastline stretching from 58°15ʹ02”N 6°08ʹ23”W to 58°14ʹ02”N 6°09ʹ23”W, on the Eye Peninsula, Isle of Lewis, UK; and P. palmata from 57°49ʹ07”N 6°49ʹ52”W to 57°49ʹ03”N 6°49ʹ48”W at Stockinish, Isle of Harris, UK. Both species were harvested between April and July 2020 to obtain kinetics for the typical harvesting season. shows the seaweeds after harvesting and before sample preparation. Harvested seaweed was immediately placed in a sealed container filled with seawater and then refrigerated at 3 ± 1°C within 1 h of harvesting. Scoping experiments showed that A. esculenta deteriorated more rapidly than P. palmata and began to discolour and develop a sharp smell after ~80 h of refrigeration; therefore, any leftover seaweed was discarded after 72 h to ensure sample consistency.
When ready for use, samples of 2.5 ± 0.2 g by wet basis (w.b.) were prepared. Whilst P. palmata is a homogenous material, A. esculenta consists of a thick, dense stripe and a thin blade; therefore, care was taken to ensure that the planar stripe-to-blade ratio of each A. esculenta sample was approximately constant. The samples were rinsed in fresh water and then shaken for 5 s to remove surface water droplets. All drying and desorption tests were performed in triplicate and the mean taken of the results.
Drying
Drying was performed with a Kern DAB 100-3 moisture analyser that removed moisture from the samples by natural convection heating at a specified temperature. This was connected by RS-232 lead to a PC for automated data recording, as shown in . The time-based drying curves were obtained with a temporal sampling resolution of 1 min and a mass accuracy of ± 0.5 mg. Each sample was considered dry (MR = 0) when the sample mass decreased by less than 10–5 g min–1 for five consecutive minutes.
Figure 1. Block diagram to show the setup for the drying and desorption experiments.
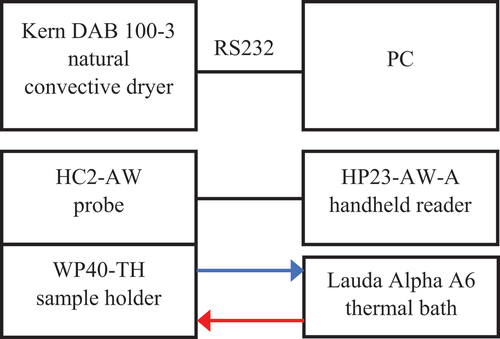
Figure 2. Photographs with a 30 cm ruler of (a) Alaria esculenta and (b) Palmaria palmata, as harvested before trimming to sample weight.

The experimental moisture ratio is calculated as
This was compared with the empirical models shown in . Lewis (Citation1921) model is often referred to in the literature as Newton’s as it derives from Newton’s law of cooling. Page (Citation1949) added the empirical constant n, which was modified by Overhults, White, Hamilton, and Ross (Citation1973).
Table 1. Drying models.
The coefficient of determination is defined as
where sum-of-squares subscripts res and T denote residuals and total, respectively.
For each model in , the fitting constants were obtained by nonlinear regression using the Trust-Region Method in MATLAB.
Desorption
Water activity was measured with Rotronic water activity metering equipment: samples were placed inside a WP-40TH water-jacketed sample holder, on which sat an HC2-AW capacitive measurement probe connected to the HP23-AW-A handheld reader, returning aw accurate to ± 0.01. The water-jacketed sample holder was fed by a thermostatically controlled Lauda Alpha A6 thermal bath.
To obtain the desorption isotherms, the samples were dried with the moisture analyser until the mass (± 0.025 g) for each data point was obtained, then transferred to the water activity meter, regulated to the desired temperature, and the water activity was read once stable for 5 min. Data points were obtained at relative humidities of [0.75, 0.5, 0.4, 0.3, 0.275, 0.25, 0.225 and 0.2] of the initial wet mass.
A comparison of the sorption models shown in was made. The Guggenheim–Anderson– de Boer (GAB) model (Equation (11)) was derived independently by Anderson (Citation1946), de Boer (Citation1953), and Guggenheim (Citation1966). It is a development of the Langmuir (Citation1918) and Brunauer, Emmett, and Teller (Citation1938) (BET) models and uses fitting parameters with physical meaning:
Table 2. Desorption models.
Table 3. Drying model parameters of A. esculenta.
Table 4. Drying model parameters of Palmaria palmata.
where c0 and k0 are entropic fitting parameters and subscripts m, n, and 1 denote monolayer, multilayer and liquid, respectively. This was compared with the empirical models of Oswin (Citation1946), Halsey (Citation1948) and Caurie (Citation1970). When evaluating these models, data points where aw > 0.95 were excluded as the models are not established for water activity values approaching unity.
The isosteric heat of sorption is:
where Hfg is the latent heat of vaporization of water at the sample temperature. The net isosteric heat of sorption is given by the Clausius–Clapeyron equation:
evaluated at fixed moisture content.
Results
Drying kinetics
shows the logarithmic drying curves for the seaweed species studied. For both species, the layer half-thickness was 0.75 mm. Across all drying temperatures, A. esculenta took longer to dry than P. palmata and the temperature effect was of the same order of magnitude: increasing air temperature from 40 to 70°C decreased total drying time by 62.4% and 61.7% for A. esculenta and P. palmata, respectively. show the drying model parameters for A. esculenta and P. palmata, respectively. All the curves are acceptably modelled by exponential functions; of the semi-empirical models, Page returned the highest mean r2 value. The Weibull model also returned excellent r2 values for both seaweeds. The β parameter was consistently lower for A. esculenta than P. palmata, indicating a drying curve that has a steeper initial profile but also levels off sooner (Bantle, Kolsaker, & Eikevik, Citation2011).
Figure 3. Effect of air temperature on drying curves of (a) Alaria esculenta and (b) Palmaria palmata.
Desorption isotherms
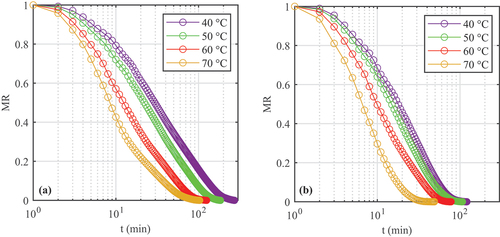
provide the desorption isotherms for A. esculenta and P. palmata, respectively; water activity was plotted on the x-axis according to convention, although it is the dependent variable in these tests. The “dry” datapoints corresponding to Xe = 0 kgwater kg–1 d.b. were not plotted; the water activities jumped notably from their previous point (as graphed) to aw < 0.5 when moisture content moved from the previous non-zero Xe to zero. From ), both species exhibit Category III isotherms according to the Brunauer classification (Brunauer, Deming, Deming, & Teller, Citation1940). Across the moisture contents tested, increasing temperature increased the measured water activity. show that all four sorption models evaluated fitted the data well, although the Oswin and Halsey models performed marginally poorer than the Caurie model. Previous seaweed desorption studies have not observed a significant performance variation between models tested (Arufe, Torres, Chenlo, & Moreira, Citation2018; Moreira, Chenlo, Sineiro, Sánchez, & Arufe, Citation2016).
Figure 4. Desorption isotherms of Alaria esculenta showing (a) full isotherms in log scale and (b) detail at low moisture content.
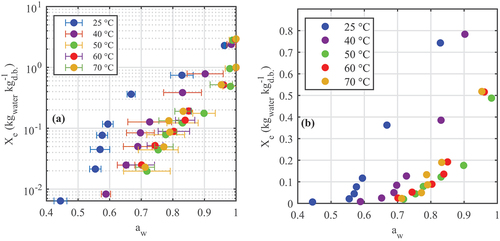
Figure 5. Desorption isotherms of Palmaria palmata showing (a) full isotherms in log scale and (b) detail at low moisture content.
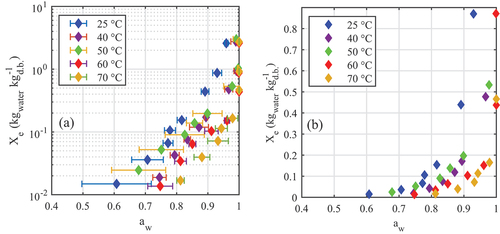
Table 5. Desorption model parameters of Alaria esculenta.
Table 6. Desorption model parameters of Palmaria palmata.
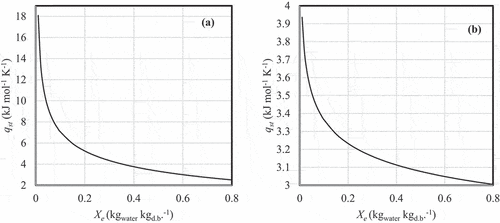
To obtain the isosteric heat of sorption, the Caurie model was used to calculate water activity for a given moisture content, and then substituted into EquationEquation (6)(6)
(6) to return the results shown in for A. esculenta and P. palmata, respectively. This shows that drying to an equilibrium moisture content of 0.01 kgwater kgd.b.–1, A. esculenta requires 4.59× the energy of P. palmata.
Figure 6. Net isosteric heat of desorption for (a) Alaria esculenta and (b) Palmaria palmata.
Nomenclature
| A | = | Sorption fitting constant |
| aw | = | Water activity |
| B | = | Sorption fitting constant |
| C | = | GAB fitting constant |
| H | = | Molar sorption enthalpy |
| H{fg} | = | Latent heat of vaporization |
| k | = | Constant |
| MR | = | Moisture ratio |
| n | = | Empirical constant |
| Qst | = | Isosteric heat of sorption |
| qst | = | Net isosteric heat of sorption |
| R | = | Gas constant = 8.314 |
| r2 | = | Correlation coefficient |
| SS | = | Sum-of-squares |
| T | = | Temperature |
| t | = | Time |
| X | = | Instantaneous moisture content |
| X0 | = | Monolayer moisture content |
| Xe | = | Equilibrium moisture content |
| Xs | = | Security moisture content |
| α | = | Weibull coefficient |
| β | = | Weibull coefficient |
Disclosure statement
No potential conflict of interest was reported by the author(s).
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
Additional information
Funding
References
- Al-Muhtaseb, A. H., McMinn, W. A. M., & Magee, T. R. A. (2002). Moisture sorption isotherm characteristics of food products: A review. Food and Bioproducts Processing, 80, 118–128.
- Anderson, R. B. (1946). Modifications of the Brunauer, Emmett and teller equation. Journal of the American Chemical Society, 68, 686–691.
- Arufe, S., Torres, M. D., Chenlo, F., & Moreira, R. (2018). Air drying modelling of mastocarpus stellatus seaweed a source of hybrid carrageenan. Heat and Mass Transfer, 54, 177–184.
- Badmus, U. O., Taggart, M. A., & Boyd, K. G. (2019). The effect of different drying methods on certain nutritionally important chemical constituents in edible brown seaweeds. Journal of Applied Phycology, 31, 3883–3897.
- Bantle, M., Kolsaker, K., & Eikevik, T. M. (2011). Modification of the Weibull distribution for modeling atmospheric freeze-drying of food. Drying Technology, 29, 1161–1169.
- Brunauer, S., Deming, L. S., Deming, W. E., & Teller, E. (1940). On a theory of the van der Waals adsorption of gases. Journal of the American Chemical Society, 62, 1723–1732.
- Brunauer, S., Emmett, P. H., & Teller, E. (1938). Adsorption of gases in multimolecular layers. Journal of the American Chemical Society, 60, 309–319.
- Caurie, M. (1970). A new model equation for predicting safe storage moisture levels for optimum stability of dehydrated foods. International Journal of Food Science & Technology, 5, 301–307.
- Chapman, A. S., Stévant, P., & Larssen, W. E. (2015). Food or fad? Challenges and opportunities for including seaweeds in a Nordic diet. Botanica Marina, 58. doi:10.1515/bot-2015-0044
- de Boer, J. H. (1953). The dynamical character of adsorption. Oxford: Clarendon Press.
- Erbay, Z., & Icier, F. (2010). A review of thin layer drying of foods: Theory, modeling, and experimental results. Critical Reviews in Food Science and Nutrition, 50, 441–464.
- Fontana, A. J. (2020). Appendix 4: Minimum water activity limits for growth of microorganisms. In Water activity in foods (2nd.ed., pp. 571–572). Chicago, IL: John Wiley & Sons.
- Fredersdorf, J., Müller, R., Becker, S., Wiencke, C., & Bischof, K. (2009). Interactive effects of radiation, temperature and salinity on different life history stages of the Arctic kelp Alaria esculenta (Phaeophyceae). Oecologia, 160, 483–492.
- Guggenheim, E. A. (1966). Application of statistical mechanics. Oxford: Oxford University Press.
- Halsey, G. (1948). Physical adsorption on non‐uniform surfaces. The Journal of Chemical Physics, 16, 931–937.
- Harnedy, P. A., & FitzGerald, R. J. (2013). In vitro assessment of the cardioprotective, anti-diabetic and antioxidant potential of Palmaria palmata protein hydrolysates. Journal of Applied Phycology, 25, 1793–1803.
- Jacobson, H. J. (2016). Values underpinning Iceland’s food system risk: Implications for resilience planning. Cambridge, MS: Massachusettes Institute of Technology. http://hdl.handle.net/1721.1/104989
- Kerrison, P. D., Innes, M., Macleod, A., McCormick, E., Elbourne, P. D., Hughes, A. D., … Kelly, M. S. (2020). Comparing the effectiveness of twine- and binder-seeding in the Laminariales species Alaria esculenta and Saccharina latissima. Journal of Applied Phycology, 32, 2173–2181.
- Kintisch, E. 2016. Greenland’s getting warmer, but farmers there are struggling more than ever. NPR. Retrieved from https://www.npr.org/sections/thesalt/2016/11/17/502349923/climate-change-is-making-greenland-warmer-but-farmers-there-are-struggling?t=1600688767834.
- Langmuir, I. (1918). The adsorption of gases on plane surfaces of glass, mica and platinum. Journal of the American Chemical Society, 40, 1361–1403.
- Lewis, W. K. (1921). The rate of drying of solid materials. Industrial and Engineering Chemistry, 13, 427–432.
- Lopes, D., Melo, T., Meneses, J., Abreu, M. H., Pereira, R., & Domingues, M. R. (2019). A new look for the red macroalga palmaria palmata: A seafood with polar lipids rich in EPA and with antioxidant properties. Marine Drugs, 17, 533.
- Lüning, K. (1990). Seaweeds: Their environment, biogeography, and ecophysiology. (C. Yarish & H. Kirkman, Eds.). New York, NY: John Wiley & Sons.
- Maehre, H. K., Edvinsen, G. K., Eilertsen, K. E., & Elvevoll, E. O. (2016). Heat treatment increases the protein bioaccessibility in the red seaweed dulse (Palmaria palmata), but not in the brown seaweed winged kelp (Alaria esculenta). Journal of Applied Phycology, 28, 581–590.
- Mai, K., Mercer, J. P., & Donlon, J. (1994). Comparative studies on the nutrition of two species of abalone, Haliotis tuberculata L. and Haliotis discus hannai Ino. Aquaculture, 128, 115–130.
- Mieszkowska, N., Kendall, M. A., Hawkins, S. J., Leaper, R., Williamson, P., Hardman-Mountford, N. J., … Southward, A. J. (2006). Changes in the range of some common rocky shore species in Britain – A response to climate change? Hydrobiologia, 555, 241–251.
- Moreira, R., Chenlo, F., Sineiro, J., Sánchez, M., & Arufe, S. (2016). Water sorption isotherms and air drying kinetics modelling of the brown seaweed Bifurcaria bifurcata. Journal of Applied Phycology, 28, 609–618.
- Morgan, K. C., & Simpson, F. J. (1981). The cultivation of Palmaria palmata. Effect of light intensity and temperature on growth and chemical composition. Botanica Marina, 24.
- Mouritsen, O. G., Dawczynski, C., Duelund, L., Jahreis, G., Vetter, W., & Schröder, M. (2013). On the human consumption of the red seaweed dulse (Palmaria palmata (L.) Weber & Mohr). Journal of Applied Phycology, 25, 1777–1791.
- Northern Periphery and Arctic Programme. (2016). Northern periphery and arctic cooperation programme 2014-2020. Retrieved from http://www.interreg-npa.eu/fileadmin/Programme_Documents/Approved_Cooperation_Programme_Jan2016.pdf.
- Onwude, D.I., Hashim, N., Janius, R.B., Nawi, N.M. & Abdan, K. (2016). Modeling the Thin-Layer Drying of Fruits and Vegetables: A Review. Comprehensive Reviews in Food Science and Food Safety, 15, 599–618.
- Oswin, C. R. (1946). The kinetics of package life. III. The isotherm. Journal of the Society of Chemical Industry, 65, 419–421.
- Overhults, D. G., White, G. M., Hamilton, H. E., & Ross, I. J. (1973). Drying soybeans with heated air. Transactions of the American Society of Agricultural Engineers, 16, 112–113.
- Page, G. E. (1949). Factors influencing the maximum rates of air drying shelled corn in thin layers. Retrieved from https://docs.lib.purdue.edu/dissertations/AAI1300089/.
- Reid, G. K., Chopin, T., Robinson, S. M. C., Azevedo, P., Quinton, M., & Belyea, E. (2013). Weight ratios of the kelps, Alaria esculenta and Saccharina latissima, required to sequester dissolved inorganic nutrients and supply oxygen for Atlantic salmon, Salmo salar, in integrated multi-trophic aquaculture systems. Aquaculture, 408–409, 34–46.
- Schiener, P., Black, K. D., Stanley, M. S., & Green, D. H. (2015). The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. Journal of Applied Phycology, 27, 363–373.
- Seterlund, W., Hoie, J., Sannan, F., & Raastad, N. (1968). The seaweed badderlocks (Alaria esculenta) as a supplement for chickens. Meldinger Fra Norges Landbrukshogskole, 47. Retrieved from https://www.cabdirect.org/cabdirect/abstract/19691408302
- Smith, A. M., & Ross, A. B. (2016). Production of bio-coal, bio-methane and fertilizer from seaweed via hydrothermal carbonisation. Algal Research, 16, 1–11.
- Springer, K., Lütz, C., Lütz-Meindl, U., Wendt, A., & Bischof, K. (2017). Hyposaline conditions affect UV susceptibility in the Arctic kelp Alaria esculenta (Phaeophyceae). Phycologia, 56, 675–685.
- Takase, T., Nakamura, A., Miyoshi, H., Koga, M., Toyomaki, A., Kino, R., … Atsumi, T. (2020, April 3). Effects of Palmaria palmata on lipid metabolism and glycemic control in participants with hypercholesterolemia in a randomized double-blind placebo-controlled trial. Phytotherapy Research, 34, 2303–2312.
- Uribe, E., Vega-Gálvez, A., García, V., Pastén, A., López, J., & Goñi, G. (2019). Effect of different drying methods on phytochemical content and amino acid and fatty acid profiles of the green seaweed, Ulva spp. Journal of Applied Phycology, 31, 1967–1979.
- van Ginneken, V. J., Helsper, J. P., de Visser, W., van Keulen, H., & Brandenburg, W. A. (2011). Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids in Health and Disease, 10, 104.
- Weibull, W. (1951). A statistical distribution function of wide applicability. Journal of Applied Mechanics, 18, 293–297.
- Werner, A., & Dring, M. (2011). Cultivating Palmaria palmata. In Aquaculture explained. Dun Laoghaire: Bord Iascaigh Mhara/Irish Sea Fisheries Board. Retrieved from http://www.bim.ie/media/bim/content/publications/Aquaculture,Explained,Issue,27,-,Cultivating,Palmaria,palmata.pdf
- Yesson, C., Bush, L. E., Davies, A. J., Maggs, C. A., & Brodie, J. (2015). Large brown seaweeds of the British isles: Evidence of changes in abundance over four decades. Estuarine, Coastal and Shelf Science, 155, 167–175.