
Abstract
The maximum productivity of plantation forestry and its role in climate change mitigation, adaptation and resilience cannot be met without proper management. Termites in the genus Macrotermes have been reported as a major challenge to Eucalyptus plantation forestry establishment. The current study evaluated the susceptibility of four Eucalyptus hybrid clones: GU 7, GC 796, GC 550 and GC 796/2, and E. grandis Hill ex Maid. to the most damaging Macrotermes bellicosus (Smeathman) and Macrotermes subhyalinus (Rambur) to identify tolerant material that can be planted in high incidence areas. The study involved exposure of moisture dry pieces of wood from E. grandis and the four hybrid clones to damage by M. bellicosus and M. subhyalinus. Results confirmed that M. bellicosus is the most aggressive. Results further revealed that E. grandis and GC 550 are the most susceptible whereas GC 796 is the most tolerant clone. The findings from the study will contribute to improved management of termites by planting tolerant material in high risk areas.
Introduction
Forests and trees are playing a vital role in mitigation, adaptation and resilience to climate change (FAO Citation2018). The forests have a role in reducing natural disasters hazards, including floods, droughts, landslides and other extreme events. Thus considerably contributing to Sustainable Development Goal 15 aimed at promoting the sustainability of terrestrial ecosystems and halting desertification, land degradation and biodiversity loss (UN General Assembly Citation2019). Additionally, the demand for forestry products and services has been increasing in response to population growth (FAO Citation2018). However, the world's natural forests have over the years faced serious degradation and deforestation (Duguma et al. Citation2019). To reduce the level of deforestation there is increasing investments in plantation forestry (FAO Citation2018).
Approximately half of all plantation forestry in the tropics and subtropics is composed of Eucalyptus species (Stanturf et al. Citation2013). Their high growth rates and their ability to grow within a wide range of climatic conditions make them attractive species for both commercial and social forestry applications. Worldwide, Eucalyptus plantation forestry is dominated by E. camaldulensis, E. grandis, E. tereticornis, E. globulus, E. nitens, E. urophylla, E. saligna, E. dunnii and E. pellita (Stanturf et al. Citation2013). In Uganda, E. grandis is the most predominant species (Jacovelli Citation2009; Kilimo Trust Citation2011). However, over the years there have been tree improvement initiatives focusing on the use of hybrid clones (Stanturf et al. Citation2013). Eucalyptus hybrid clones are genetically superior with important traits such as pest and disease resistance, fast and uniform growth as well as homogenous physical properties (Dehon et al. Citation2014).
Eucalyptus hybrid clones are currently the most preferred plantation species in Uganda (Kilimo Trust, Citation2011). The commonly preferred hybrids are from E. grandis and E. camaldulensis (GC) as well as from E. grandis and E. urophylla (GU) (Epila-Otara and Ndhokero Citation2010; Kilimo Trust Citation2011). It’s expected that the effective use of Eucalyptus hybrid clones in plantation forestry will reduce timber and wood shortage in Uganda and worldwide (Jacovelli Citation2009; Kilimo Trust Citation2011; Wingfield et al. Citation2003; Dehon et al. Citation2014). However, previous research reported termites as being a major threat to Eucalyptus plantations (Nyeko and Nakabonge Citation2008; Ssemaganda et al. Citation2011; Orikiriza et al. Citation2012; Namujehe and Orikiriza Citation2014).
The genus Macrotermes consist of fungus – growing termites that are known to build large mounds and the most destructive in exotic plantations (Nyeko and Nakabonge Citation2008; Ssemaganda et al. Citation2011; Orikiriza et al. Citation2012; Namujehe and Orikiriza Citation2014). The Macrotermes belongs to subfamily Macrotermitinae that are characterized by possessing a digestive system that is incapable of breaking down the cellulose. They have therefore co-evolved an obligate symbiotic relationship with fungi of the genus Termitomyces (Schuurman Citation2005). The fungus helps these termites to degrade the cellulose, thus are capable of decomposing wood at higher rates than other termite genera. Termites cause significant damage to Eucalyptus trees especially when young and establishing by destroying the root system. As a management strategy it is important to identify tolerant Eucalyptus hybrids that could be planted in areas with high termite infestation. Previous studies attempted to understand the natural durability of Eucalyptus hybrid clones grown in Uganda, against termite attack (Namujehe and Orikiriza Citation2014). However, the study evaluated only two hybrid clones (GU 7 and GC 796). This study evaluated the susceptibility of wood from four Eucalyptus hybrid clones: GU 7, GC 796, GC 550 and GC 796/2, and E. grandis to Macrotermes bellicosus (Smeathman) and Macrotermes subhyalinus (Rambur). The findings from the study will support efforts towards termite control and management in Uganda.
Materials and methods
The experiment was carried out in Kabanyoro village (0.4637°N, 32.6099°E) located in Nangabo sub county, Wakiso, Central Region, Uganda.
Five wood species of Eucalyptus grandis and four Eucalyptus hybrid clones, GU 7, GC 796, GC 550 and GC 796/2 that are widely grown in Uganda, were used in this study. For each Eucalyptus species, one standing tree was randomly selected and harvested from the plantations of Core woods company, Hoima district, western Uganda. The poles were cut from 1.5 m at diameter breast height, debarked and air seasoned for 2 weeks to about 20% moisture content. Short billets were obtained by cross cutting, thereafter, wood test pieces (length × width × thickness: 25 × 2 × 2 cm) were cut from timber boards as described by Namujehe and Orikiriza, (Citation2014).
For each genotype, 24 stakes were generated to make a total of 120. Wood samples were randomly selected and labeled appropriately by end painting for easy identification. The stakes were oven dried initially at 60°C followed by 80°C and weight measurements taken 6 hourly to obtain the final initial weights (W1) of the wood stakes.
Termite identification
The termite species were collected from the mounds and identified from Makerere University Agricultural Research Institute Kabanyolo (MUARIK). The soldier and worker castes were collected and identified at species and genus level with help of experts from the pest management section of MUARIK.
Field exposure of wood samples to termites
Field exposure was accomplished using a completely randomized experimental design (CRD) with a 5 × 2 factorial treatment, i.e. 5 timber genotypes and 2 termite species. The experiment had four replicates with 40 experimental units in each experimental setup.
Four plots of 1 × 1 sq. m. were established, 5 m away from the active termite mounds but in an area with termite tunnels. Five timber stakes of each Eucalyptus genotype were randomly distributed in a plot (replicates). The stakes were buried vertically into the soil, with approximately 1/5 of the total length protruded above the ground. The stakes were evaluated at 3 exposure periods, i.e. 2, 4 and 6 weeks.
At the end of each exposure period, the stakes were removed, cleaned and oven dried at 80°C until there was no change in weight. These were then conditioned to equilibrium moisture content and then weighed to get their final weight (W2) in order to calculate wood consumption percentage in terms of weight loss (P). The wood removed was defined as initial minus final weights with test duration periods equivalent to the time during which the stakes were exposed to termites in the field (2, 4 and 6 weeks). Data was analyzed using one-way ANOVA in IBM SPSS Statistics for Windows, version 19.0 (IBM Corp., Armonk, N.Y., USA) to determine the percentage wood loss in E. grandis and hybrid clones exposed to the two termite species under the three exposure periods.
Results
Comparison of M. bellicosus and M. subhyalinus wood destruction
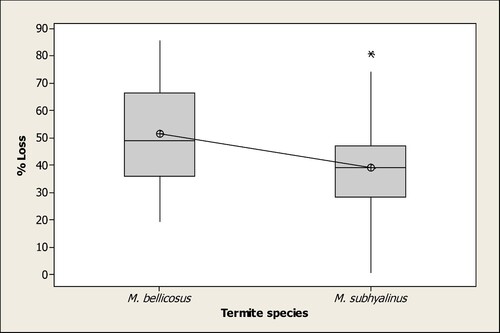
The results indicate that all wood samples used in the study were damaged by M. bellicosus and M. subhyalinus following a 6-week exposure period. However, the amount of damage due to the two species differed significantly (p < 0.000). M. bellicosus was more destructive with a mean percentage loss of 51.5%, R = 4 and SD 18.6 compared to M. subhyalinus with 39.2%, R = 4 and SD 17.3 (Figure ).
Figure 1. Percentage wood loss (data pooled) by two termite species Macrotermes bellicosus and M. subhyalinus over a 6-week exposure to Eucalyptus dry stakes in Uganda (*significantly different at p < 0.000).
Variation in susceptibility to wood destruction among E. grandis and hybrid clones
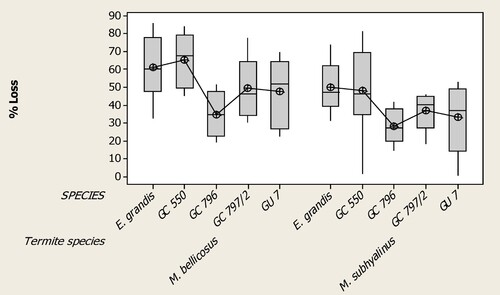
There were significant differences in wood damage due to M. bellicosus on E. grandis and the 4 hybrid clones used in the study. Clone GC 550 was the most susceptible with a mean percentage loss of 65.2% and SD 14.5% followed by E. grandis with a mean percentage value of 60.9% and SD 16.7, GC 797/2 with of 49.3% and SD 16.1, GU 7 with a mean value of 47.4% and SD 17.9 and GC 796 with the least mean weight loss of 34.5% and SD 12.3, respectively (Df = 4, SS = 7031, MS = 1758, F = 7.2, p = 0.000, N = 12). Although the mean weight loss amongst the wood species was lower for M. subhyalinus compared to M. bellicosus, there was significant differences observed (Figure ). E. grandis had the highest mean value of 49.9% and SD 14.3, followed by GC 550 with a mean loss of 48.3% and SD 22.4, GC797/2 with 36.7% and SD 9.5, GU 7 with mean weight loss of 33.2% and SD 18.0 and GC 796 presented the lowest mean weight loss of 27.9% and SD 9.6 (Df = 4, SS = 4407, MS = 1102, F = 4.54, p = 0.003, N = 12).
Figure 2. Percentage wood loss of 6 Eucalyptus genotypes (data pooled) by two termite species Macrotermes bellicosus and M. subhyalinus over a 6-week exposure to dry stakes in Uganda (*significantly different at p = 0.003).
Wood damage by M. bellicosus and M. subhyalinus at various exposure periods
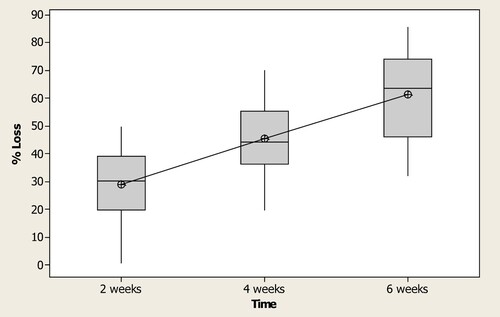
Wood damage differed significantly at the three exposure periods. Evaluation at 2 weeks revealed the lowest mean weight loss of 29.2% and SD 12.8, followed by 45.5% and SD 12.8 at 4 weeks and 61.4% and SD 15.4 being the highest at 6 weeks respectively (Df = 2, SS = 20,739, MS = 10,369, F = 55.21, p = 0.000, N = 40).
Discussion
The study evaluated the susceptibility of E. grandis and four Eucalyptus hybrid clones commonly grown in Uganda to two termite species, M. bellicosus and M. subhyalinus (Figure ). Results revealed varying susceptibility of Eucalyptus to the two termite species and also variation in damage where, M. bellicosus caused a higher weight loss percentage compared to M. subhyalinus (Figure ).
Figure 3. Eucalyptus wood damage following exposure to M. bellicosus and M. subhyalinus: (A) M. bellicosus mound, (B) M. subhyalinus mound, (C) wood stakes following exposure to termite damage, (D) wood stakes half inserted in the soil in proximity with termite mounds and mud tubes and (E) wood stakes after exposure to termite attack.

The study revealed significant (p < 0.000) differences in percentage weight loss between the two termite species under the conditions in this experiment (Figure ). M. bellicosus is recognized as important players in ecosystem balance and participate in improvement of soil fertility (Ajao et al. Citation2018). However, they are known to cause tremendous damage to trees and wood products. The higher weight loss percentage by M. bellicosus is a confirmation of previous studies that have indicated that it is a serious pest of Eucalyptus species (Nyeko and Nakabonge Citation2008; Ssemaganda et al. Citation2011; Orikiriza et al. Citation2012; Namujehe and Orikiriza Citation2014). M. bellicosus have got aggressive soldiers with large scissor like mandibles and high defensive mechanisms that make them voracious feeders (Ogedegbe and Eloka Citation2015). The damage on Eucalyptus wood species by M. bellicosus could also be attributed to their enormous fixed nests, which harbor large populations, that in search for food end up damaging, trees, wood and wood products (Orhue et al. Citation2007).
Although the percentage weight loss was lower for M. subhyalinus, there was considerable wood reduction observed (Figures and ). M. bellicosus and M. subhyalinus have previously been reported to be the most destructive of all Macrotermes species capable of destroying both fresh and dry wood (Mali et al. Citation2018).
Significant differences in the level of susceptibility were recorded among E. grandis and hybrid clones to termite damage. Clone GC 550 and E. grandis were the most susceptible to M. bellicosus followed by clones GC 797/2, GU 7 and GC 796 respectively. The findings confirm previous studies which indicated that GC 796 is more tolerant to termite attack than GU 7 (Namujehe and Orikiriza 2013). Genetic differences have been recorded for growth and wood traits in Eucalyptus populations, including diameter, wood basic density and lignin content (Chen et al. Citation2018). It is thus possible that the variation in susceptibility reported during this study is attributed to one or a combination of these factors. The response to the termite species by all clones used in the study followed by a similar trend, where E. grandis and GC 550 were the most susceptible followed by GC 797/2, GU 7 and GC 796 being the most tolerant. However, different from M. bellicosus, there was a significant difference in susceptibility between E. grandis and GC 550, for wood pieces subjected to damage by M. subhyalinus (Figure ).
Results further indicated that weight loss generally increased with increase in the exposure period (Figure ). This increase in consumption could be attributed to termites' mode of feeding where they begin with construction of tunnels that connect the nests to the sources of food (Peralta et al. Citation2004; Sim and Lee Citation2012).
Figure 4. Percentage wood loss of six Eucalyptus genotypes (data pooled) by two termite species over at a 2-week time interval up to a 6-week exposure period to dry stakes in Uganda (*significantly different at p < 0.000).
Hybrid clones were introduced to East Africa due to their fast growth rates, uniformity and resistance to pests and diseases (Kilimo Trust Citation2011). The information generated in this study further confirms that planting material with tolerance to termite damage is available in the region. The most tolerant clone GC 796 during this study is consistent with previous findings. It could thus serve as an option for planting in areas with high incidence of Macrotermes species. Additionally, further evaluations in other agro-ecologies could be assessed, since environmental factors may affect the behavior of the termites and resistance of the trees.
Acknowledgements
We acknowledge financial and technical support provided by the College of Agricultural and Environmental Sciences, Makerere University, Uganda.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement (DAS)
Data for the manuscript is available at doi:10.5061/dryad.9p8cz8wd3.
Additional information
Funding
References
- Ajao AM, Oladimej YU, Oladipo SO, Adepoju SA. 2018. Activity of mound-building Macrotermes bellicosus (isoptera: termitidae) around kwara state university campus Guinea savannah ecozone, Nigeria. Anim Res Int. 15(1):2918–2925.
- Chen S, Weng Q, Li F, Li M, Zhou C, Gan S. 2018. Genetic parameters for growth and wood chemical properties in Eucalyptus urophylla × E. tereticornis hybrids. Ann For Sci. 75:16. https://doi.org/10.1007/s13595-018-0694-x.
- Dehon SP, Rezende G, Resende MDV, Assis T. 2014. Eucalyptus breeding for clonal forestry. Challenges and opportunities for the world's forests in the 21st century. For. Sci. 81:393–424.
- Duguma LA, Atela J, Minang PA, Ayana AN, Gizachew B, Nzyoka JM, Bernard F. 2019. Deforestation and Forest degradation as an environmental behavior: Unpacking realities shaping community actions. Land (Basel). 8(2):26. https://doi.org/10.3390/land8020026.
- Epila-Otara JS, Ndhokero J. 2010. Selection and site matching of Eucalyptus clones in Uganda. J East Afr Natural Res Manage. 3(1):237–248.
- FAO. 2018. The State of the World’s Forests 2018 - Forest pathways to sustainable development. Rome. License: CC BY-NC-SA 3.0 IGO
- Jacovelli PA. 2009. Uganda’s Sawlog Production Grant Scheme: a success story from Africa. Int Forest Rev. 11:119–125.
- Kilimo Trust. 2011. Eucalyptus hybrid clones in East Africa. Meeting the demand for wood through clonal forestry technology. KILIMO TRUST. 3(1):1–24.
- Mali B, Okello S, Ocaido M, Nalule AS, Celsus S. 2018. Ethno charaterisation of termite species in the rangelands of central Uganda. Livestock Res Rural Dev. 30(8). http://www.lrrd.org/lrrd30/1/b3ma30008.html.
- Namujehe G, Orikiriza LJB. 2014. Natural durability of Eucalyptus clones against termite attack. Int J Sci Basic Appl Res. 10(1):176–183.
- Nyeko P, Nakabonge G. 2008. Occurrence of pests and diseases in tree nurseries and plantations in Uganda. Study Commissioned by the Sawlog Production Grant Scheme, 256 (October), 42
- Ogedegbe ABO, Eloka EV. 2015. Biodiversity of termites on Farmlands in Ugoniyekorhionmwon, Orhionwmon Local Government area, Edo State, Nigeria. Int J Pure Appl Sci Technol. 27(2):65–76.
- Orhue ER, Uzu OF, Osaigbovo UA. 2007. Influence of activities of termites on some physical and chemical properties of soils under different land use patterns: a review. Int J Soil Sci. 2(1):1–14.
- Orikiriza LJB, Nyeko P, Sekamatte B. 2012. Farmers’ knowledge, perceptions and control of pestiferous termites in Nakasongola district, Uganda. Uganda J Agric Sci. 13(1):71–83.
- Peralta RCG, Menezes EB, Carvalho AG, Aguiar-Menezes EL. 2004. Wood consumption rates of forest species by subterranean termites (Isoptera) under field conditions. Rev Árvore. 28(2):283–289.
- Schuurman G. 2005. Decomposition rates and termite assemblage composition in semiarid Africa. Ecology. 86(5):1236–1249.
- Sim S, Lee S-H. 2012. Measurement of the time required for a termite to pass through tunnels with different curvatures. J Insect Sci. 12(64):1–7.
- Ssemaganda IE, Mugabi P, Tumwebaze SB. 2011. Effectiveness of selected preservatives in protecting Ugandan grown Eucalyptus grandis wood against termite attack. Maderas Cien Tecnol. 13(2):135–142.
- Stanturf JA, Vance ED, Fox TR, Kirst M. 2013. Eucalyptus beyond its native range: environmental Issues in exotic bioenergy plantations. Int J Forest Res. http://dx.doi.org/10.1155/2013/463030.
- UN General Assembly. 2019. Transforming our world: the 2030 Agenda for Sustainable Development, 21 October 2015, A/RES/70/1. https://www.refworld.org/docid/57b6e3e44.html [accessed 7 June 2019]
- Wingfield M. J. 2003. Increasing threat of diseases to exotic plantation forestry in the southern hemisphere: lesson learnt from Cryphonectria canker. Australas. Plant Pathol. 32:133–136.