
Abstract
The orchid mantis Hymenopus coronatus (Insecta: Mantodea) is famous for its unique appearance, which resembles a flower blossom with petal-like legs and a whitish pink colour. Based on this appearance, H. coronatus has been hypothesized to be an aggressive mimic that attracts pollinators. Research on the orchid mantis has been sparse due to the difficulty of collection and maintenance. The first record of orchid mantis from Xishuangbanna, China, was originally named H. coronatoides but later considered synonymous with H. coronatus based on a morphological study. In this study, the complete mitochondrial genome of the orchid mantis from Xishuangbanna, China, was determined to be a circular molecule of 15,589 bp containing 37 typical mitochondrial genes in the same gene arrangement as those of other praying mantises. The relatively low degree of genetic difference between orchid mantises from China and Malaysia suggests that they belong to the same species. The phylogenetic relationships inferred from 13 protein-coding genes (PCGs) show that H. coronatus is a sister to Parymenopus davisoni, another rare species from Malaysia, and supports monophyletic Hymenopodinae, whereas Acromantinae is polyphyletic. This study provides useful data for studies on the population genetics, phylogeny and mimicry mechanism of the orchid mantis.
Mantises comprise more than 2300 species with morphological and ecological diversity. These species occupy a wide variety of habitats, including tropical rainforests, temperate and arid forests, and deserts, and they employ different hunting strategies (Ehrmann Citation2002; Svenson and Whiting Citation2004). The most attractive species of praying mantis, Hymenopus coronatus, commonly known as the orchid mantis, belongs to subfamily Hymenopodidae and is mainly distributed in the tropical forests of Southeast Asia, including in Malaysia, Burma, Thailand and Indonesia. This species is characterized by brilliant colours, particularly in juveniles, and it is predominantly white with pink accents, similar to some orchids. The four walking legs expand outwards, similar to flower petals (O’hanlon et al. Citation2013). In addition, H. coronatus shows significant sexual dimorphism: males are less than half the size of females (Svenson et al. Citation2016). Wang et al. (Citation1994) reported H. coronatoides, the first record of orchid mantis in Xishuangbanna, China, which differs from H. coronatus in the morphological characteristics of the wing apex and spine number of the fore tibia. However, Delfosse (Citation2007) proposed that H. coronatoides is synonymous with H. coronatus since traits vary among individuals. Although the orchid mantis is a good mimicry model, it is an understudied group of mantises due to the difficulty of collection and maintenance. Molecular sequences from orchid mantis are relatively scarce, impeding studies of the phylogeny and mimicry mechanisms of orchid mantis. In this study, we assembled the mitochondrial genome of orchid mantises from Xishuangbanna, China, based on Illumina sequencing data of genomic DNA.
Specimens were deposited at the Institute of Insect Science and Technology, South China Normal University, Guangzhou, China (registration number: Li-Hco-20200606). Total DNA was extracted from leg muscle using a DNeasy Blood and Tissue Kit (Qiagen, Germany). Based on the Illumina sequencing data of genomic DNA (our unpublished data), we used GetOrganelle-1.7.5 (Jin et al. Citation2020) to assemble the mitochondrial genome and annotated it using Mitos (Bernt et al. Citation2013) and GeSeq (Tillich et al. Citation2017). The circular representation of the mitochondrial genome was plotted using CGview Server (Grant and Stothard Citation2008).
The complete circular mitochondrial genome (Figure ) is 15,589 bp in length with a significant A + T bias (A + T content: 75.49%, A: 39.89%, T: 35.60%, G: 9.57% and C: 14.94%). It encodes 37 typical metazoan mitochondrial genes. ATN (N representing A, T, C, G) is used as the start codon for the 12 PCGs, except cox1, which uses TTG. Most PCGs end in TAA, while the termination codon for nad5 is an incomplete codon (T–). The PCGs are arranged in the same order as in other praying mantises (Ye et al. Citation2016).
Figure 1. The mitochondrial gene organizations of the Hymenopus coronatus from Xishuangbanna, China.
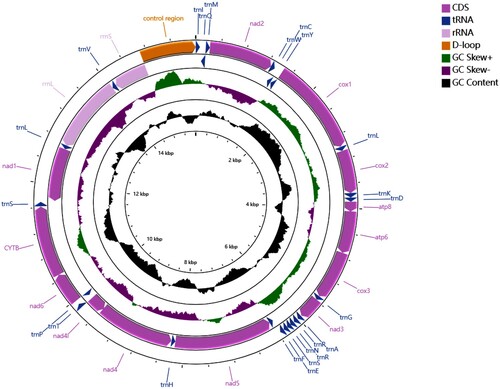
Our mitochondrial genome of orchid mantis from China is highly similar to that of H. coronatus from Malaysia (GenBank accession no. KX434861), with an overall genetic distance of 0.04. Their nucleotide differences are 3.64% ∼ 6.92% for 13 protein-coding genes (3.8% for 658 bp DNA barcode sequences), 0% ∼ 6.85% for 22 tRNA genes, 3.15% ∼ 5.03% for two ribosomal RNA genes, and 6.52% for the noncoding control region. The main difference between the two is that there are three copies of trnR in H. coronatus from Malaysia but only two copies in H. coronatus from China. However, the variation of trnR repeats is a common feature of mantis mitogenomes (Zhang et al. Citation2018; Xu et al. Citation2021). Overall, the relatively low degree of genetic difference suggests that the orchid mantises from China and Malaysia belong to the same species, consistent with the morphological study of Delfosse (Citation2007).
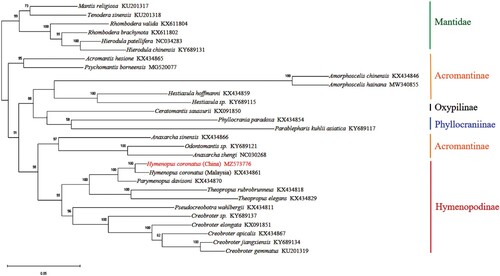
Twenty-two available complete mitochondrial genomes of Hymenopodidae were retrieved from the GenBank database in July 2021, including ten species from Hymenopodinae, nine species from Acromantinae, one species from Oxypilinae, and two species from Phyllocraniinae. Six species of Mantidae retrieved from GenBank were used as outgroups. The amino acid sequences of 13 PCGs from 29 species were aligned and concatenated with PhyloSuite-1.2.2 (Zhang et al. Citation2020). Based on maximum likelihood (ML) analysis, phylogenetic relationships were constructed using MEGA-10.1.2 (Kumar et al. Citation2018) and RAXML-8.2.12 (Stamatakis Citation2014) (Figure ). JTT + F was selected as the optimal model for ML analysis. All analyses yielded the same topology: H. coronatus is sister to Parymenopus davisoni, another rare species native to remote jungles in Malaysia, and the monophyly of Hymenopodinae is well supported. However, species of Acromantinae are separated into two clades by Oxypilinae and Phyllocraniinae.
Figure 2. The phylogenetic tree of Hymenopodidae based on amino acid sequences of 13 PCGs using the maximum-likelihood method in RAxML. Six species of Mantidae were used as outgroups. The GenBank accession numbers are given following each species name.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The mitochondrial genome sequence data that support the findings of this study are openly available in NCBI at www.ncbi.nlm.nih.gov under the accession number MZ573776.
Additional information
Funding
References
- Bernt M, Donath A, Jühling F, Externbrink F, Florentz C, Fritzsch G, Pütz J, Middendorf M, Stadler SF. 2013. MITOS: improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 69(2):313–319.
- Delfosse E. 2007. Nouvelles localisations et nouvelle synonymie pour la Mante-orchidée Hymenopus coronatus (Olivier, 1792) (Mantodea, Hymenopodidae). Bulletin de la Société entomologique de France. 112(4):529–530.
- Ehrmann R. 2002. Mantodea: Gottesanbeterinnen der Welt. Münster: Natur und Tier Verlag.
- Grant JR, Stothard P. 2008. The CGView Server: a comparative genomics tool for circular genomes. Nucleic Acids Res. 36:W181–W184.
- Jin JJ, Yu WB, Yang JB, Song Y, dePamphilis CW, Yi TS, Li DZ. 2020. Getorganelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 21:241.
- Kumar S, Stecher G, Li M, Knyaz C, Tamura K. 2018. MEGA x: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 35(6):1547–1549.
- O’hanlon JC, Li D, Norma-Rashid Y. 2013. Coloration and morphology of the orchid mantis Hymenopus coronatus (Mantodea: Hymenopodidae). J Orthoptera Res. 22(1):35–44.
- Stamatakis A. 2014. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 30(9):1312–1313.
- Svenson GJ, Brannoch SK, Rodrigues HM, O’Hanlon JC, Wieland F. 2016. Selection for predation, not female fecundity, explains sexual size dimorphism in the orchid mantises. Sci Rep. 6:37753.
- Svenson GJ, Whiting MF. 2004. Phylogeny of Mantodea based on molecular data: evolution of a charismatic predator. Syst Entomol. 29(3):359–370.
- Tillich M, Lehwark P, Pellizzer T, Ulbricht-Jones ES, Fischer A, Bock R, Greiner S. 2017. Geseq – versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 45(W1):W6–W11.
- Wang T, Liu X, Yin H. 1994. A new species of Hymenopus (Mantodea: Hymenopodidae: Hymenopodinae) from China. Entomotaxonomia. 16(2):79–81.
- Xu XD, Guan JY, Zhang ZY, Cao YR, Storey KB, Yu DN, Zhang JY. 2021. Novel tRNA gene rearrangements in the mitochondrial genomes of praying mantises (Mantodea: Mantidae): translocation, duplication and pseudogenization. Int J Biol Macromol. 185:403–411.
- Ye F, Lan X, Zhu W, You P. 2016. Mitochondrial genomes of praying mantises (Dictyoptera: Mantodea): rearrangement, duplication, and reassignment of tRNA genes. Sci Rep. 6:25634.
- Zhang D, Gao F, Jakovlić I, Zou H, Zhang J, Li WX, Wang GT. 2020. Phylosuite: an integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol Ecol Resour. 20(1):348–355.
- Zhang LP, Yu DN, Storey KB, Cheng HYI, Zhang JY. 2018. Higher tRNA gene duplication in mitogenomes of praying mantises (Dictyoptera, Mantodea) and the phylogeny within Mantodea. Int J Biol Macromol. 111:787–795.