
Abstract
Objective
Elevated spider fear is characterised by distinctive behavioural and emotional components. The former involves an unusually strong tendency to behaviourally avoid spiders, whereas the latter involves unusually strong growth of state anxiety as proximity to a spider increases. Numerous studies have been conducted to investigate whether biased attentional responding to spider information causally contributes to the behavioural symptoms of spider fear. The findings of these studies suggest that such bias may not causally underpin the behavioural symptoms of spider fear. However, no study has yet examined whether attentional bias to spider information influences the emotional symptoms of spider fear. This was the purpose of this study.
Method
Participants were exposed to an attentional bias modification (ABM) procedure, configured to induce either a decrease (avoid spider training) or an increase (attend spider training) in attentional bias to spider information, and then required to approach a live spider. The impact of this ABM training on the behavioural (how close participants approached the spider), and emotional symptoms (rate of growth in state anxiety as proximity to the spider increased), of spider fear was assessed.
Results
The induction of a group difference in attentional bias to spider information influenced the degree to which state anxiety became elevated as proximity to a spider increased, but not patterns of behavioural approach to a spider.
Conclusions
These results suggest that attentional bias to spider information causally contribute to the emotional component of spider fear, but may not functionally underpin the behavioural component of spider fear.
What is already known about this topic?
Elevated spider fear is characterised by distinctive behavioural and emotional components. The former involves an unusually strong tendency to behaviourally avoid spiders, while the latter involves unusually strong growth of state anxiety as proximity to a spider increases.
There is considerable evidence showing that such elevated levels of spider fear are characterised by an attentional bias to spider information.
Using an attentional bias modification (ABM) approach, the findings from previous studies suggest that attentional bias to spider information may not causally contribute to the behavioural symptoms of spider fear. It is yet not known, however, whether such bias causally contributes to the emotional component of spider fear.
What this topic adds?
We employed ABM to investigate whether biased attentional responding to spider information influences the degree to which state anxiety becomes elevated as proximity to a spider increased (i.e., the emotional component of spider fear).
Our present findings suggest that attentional bias to spider information can causally contribute to the emotional component of spider fear. Like previous research, we found no evidence that attentional bias to spider information makes a causally contribution to the behavioural component of spider fear.
The present demonstration that the modification of attentional responding to spider information can influence the emotional component of spider fear suggests that such ABM may usefully contribute to the treatment of spider phobia.
Elevated trait anxiety reflects an increased tendency to experience heightened state anxiety in response to a diverse range of stressors. This type of general anxious disposition can be distinguished from more specific fears, such as spider fear. Elevated spider fear reflects heightened anxiety responses only to spider‐related stimuli, and such specific fear is characterised by distinctive behavioural and emotional components (Merckelbach, de Jong, Muris, & van den Hout, Citation1996). The behavioural component involves an unusually strong tendency to behaviourally avoid spiders, while the emotional component involves unusually strong growth of state anxiety as proximity to a spider increases. Spider fear is assessed by questionnaire measures such as the Fear of Spiders Questionnaire (FSQ; Szymanski & O'Donohue, Citation1995), and extreme levels are characteristic of spider phobia (SP), which involves dysfunctional levels of anxiety symptoms under circumstances that involve potential or real exposure to spiders.
It is widely accepted that both elevated levels of general anxious disposition, and elevated levels of spider fear, are characterised by biased patterns of selective attention, respectively operating to favour the processing of generally threatening information, such as images depicting physical violence or illness, and spider information, such as images of spiders (c.f. Bar‐Haim, Lamy, Pergamin, & Bakermans‐Kranenburg, Citation2007; Mathews & MacLeod, Citation2005). A very common procedure adopted to assess attentional bias is the attentional probe task (MacLeod, Mathews, & Tata, Citation1986). In this task, participants are briefly exposed, usually for 500-ms, to emotionally discrepant stimulus pairs comprising one threatening stimulus and one non‐threatening stimulus. A probe (e.g., a small line that is either horizontally or vertically aligned) subsequently appears in the location of either one of the stimulus pair members. Participants are required to rapidly discriminate the identity of this probe (e.g., whether it is a horizontal line or a vertical line). Using this probe task, it has been repeatedly shown that participants characterised by high levels of general anxious disposition are relatively speeded to discriminate probes appearing in the location of generally threatening stimuli compared to non‐threatening stimuli, supporting the hypothesis that general anxious disposition is characterised by an attentional bias to generally threatening information (Bradley, Mogg, White, Groom, & Bono, Citation1999; Broadbent & Broadbent, Citation1988; Mogg, Mathews, & Eysenck, Citation1992). Of most relevance to the present research, it also has been demonstrated that participants who report elevated levels of spider fear are relatively speeded to discriminate probes that appear in the location of spider‐related stimuli compared to non‐spider stimuli (Mogg & Bradley, Citation2006; Vrijsen, Fleurkens, Nieuwboer, & Rinck, Citation2009), suggesting that they preferentially attend to the locus of spider information.
Thus, there is robust evidence that elevated spider fear is associated with an attentional bias to spider information. However, the demonstration of such an association does not permit the conclusion that such attentional bias makes a causal contribution to spider fear. To test this causal hypothesis, it would be necessary to directly modify biased attentional responding to spider information, to determine whether such attentional bias modification (ABM) impacts upon spider fear. Researchers have developed procedures designed to elicit this ABM process. These procedures are commonly training variants of attentional assessment tasks, configured to systematically induce attentional avoidance of, or attentional vigilance for, a specific category of threatening stimuli. The most frequently used ABM procedure is based on the attentional probe task (MacLeod et al., Citation1986). In the assessment version of this task, probes are displayed equally often in the location of threatening and non‐threatening members of stimulus pairs. In the training version of this task used in ABM studies, however, a contingency between the position of the probe, and the position of the threatening stimulus member, is introduced. Specifically, probes are always displayed either in the locus opposite to where the threatening stimuli appeared (avoid threat training), or in the same locus as where the threatening stimuli appeared (attend threat training). Often researchers will deliver a single session of such ABM training, to induce a transient group difference in attentional selectivity, and immediately afterwards will expose participants a stressor task, designed to evoke the anxiety responses of interest. If attentional bias towards a specific category of threatening stimuli makes a causal contribution to the corresponding anxious disposition, then inducing a group difference in attentional selectivity to such stimuli will lead to differential patterns of anxiety responses during the stressor task.
The first to employ this probe‐based ABM procedure were MacLeod, Rutherford, Campbell, Ebsworthy, and Holker (Citation2002), who used it to test the hypothesis that biased attentional responding to generally threatening information causally contributes to general anxious disposition. The investigators recruited non‐clinical participants who scored in the middle tercile of the STAI‐T trait anxiety score distribution. Half received a single session of the avoid threat ABM condition, and half received a single session of the attend threat ABM condition. To determine whether this ABM manipulation induced the intended group difference in attentional selectivity, an assessment version of the probe task, in which no training contingency was present, was delivered after the ABM training. Immediately thereafter, participants were exposed to a general stressor, comprising a demanding anagram task. Anxious mood state was assessed before and after this stressor, to reveal the degree to which it served to elevate state anxiety, which provided an index of general anxious disposition. MacLeod et al. (Citation2002) found that the ABM procedure successfully induced a group difference in attentional bias to generally threatening stimuli, as intended. Of most importance, when participants were subsequently exposed to the stressor, those who had just completed the avoid threat ABM training showed attenuated anxiety responses to the stressor compared to those who had just completed the attend threat ABM training. Hence, these findings provide compelling support for the hypothesis that an attentional bias to generally threatening stimuli causally contributes to general anxiety vulnerability. This finding has been replicated numerous times in both non‐clinical and clinical samples (c.f. Grafton & Macleod, Citation2014; MacLeod & Grafton, Citationin press). The conclusion that this ABM manipulation exerts a genuine impact on anxious disposition, that cannot be attributed to experimenter demand or expectancy effects, has been bolstered by the demonstration that the manipulation does not influence self‐reported expectancies concerning the anticipated impact of the stressor on anxiety symptomatology, despite influencing the anxiety responses that are experienced when the stressor is actually delivered (See, MacLeod, & Bridle, Citation2009).
Clearly, a similar methodological framework can be adopted to determine whether biased attentional responding to spider information causally contributes to elevated spider fear. Participants can be given an ABM procedure configured to induce either attentional avoidance of spider information or attentional vigilance for spider information, then be required to approach a spider, permitting the experimenter to assess the impact of this ABM manipulation on both the behavioural symptoms of spider fear (by measuring how closely they approach the spider), and on the emotional symptoms of spider fear (by measuring the rate at which state anxiety grows as proximity to the spider increases).
A number of researchers now have taken the important step of employing ABM to investigate whether biased attentional responding to spider information causally contributes to spider fear (Luo et al., Citation2015; Reese, McNally, Najmi, & Amir, Citation2010; Van Bockstaele et al., Citation2011). In these studies, non‐clinical participants have been exposed to a probe‐based ABM procedure, usually configured to induce attentional avoidance of, or attentional vigilance for, spider information. Following this ABM training, participants have been required to complete a stressor task that involves approaching a live spider, to determine whether ABM influences symptoms of spider fear. In each study, the ABM procedure employed has proven capable of inducing the intended group difference in attentional responding to spider information. However, it has consistently been found that participants in the two ABM conditions have not differed in terms of their degree of behavioural approach to the spider, suggesting that biased attentional responding to spider stimuli may not causally contribute to this behavioural symptom of spider fear. Each previous study also took a measure of state anxiety, and failed to find evidence that participants in the two ABM conditions came to differ in anxiety experienced during the stressor task. However, none of these studies have examined whether ABM influences the rate at which state anxiety grows as proximity to a spider increases. Specifically, Reese et al. (Citation2010) only compared anxiety levels reported at the highest level of behavioural approach achieved on the stressor task when it was delivered prior to the ABM procedure, with anxiety levels reported at the corresponding level of behavioural approach (which may not have been the highest level achieved) when the stressor task was delivered the day after training. Van Bockstaele et al. (Citation2011) asked participants to retrospectively report only the average level of anxiety they experienced during the stressor task. Lastly, while Luo et al. (Citation2015) assessed participants’ heart rate during the stressor task (to index state anxiety), only average heart rate was considered in their data analysis. Thus, while the findings of these previous studies suggest that biased attentional responding to spider information may not causally contribute to the behavioural symptoms of spider fear, these studies have not directly tested whether such attentional selectivity causally contributes to individual differences in degree to which state anxiety becomes increasingly elevated as a function of increasing proximity to a spider. So it remains unknown whether variability in attentional selectivity to spider information can make a causal contribution to individual differences in this emotional component of spider fear.
In this study, we sought to overcome these methodological limitations by directly examining whether the induction of a group difference in attentional responding to spider information, using ABM, serves to produce a corresponding group difference in the degree to which state anxiety becomes increasingly elevated as proximity to a spider increases. Participants reporting mid‐range scores (i.e., scores that fell within the second and third quartiles) on the FSQ were exposed to a probe‐based ABM procedure configured to induce attentional avoidance of, or attentional vigilance for, spider stimuli. The efficacy of this training procedure was determined by analysing probe discrimination latencies obtained from assessment trials, presented before and after the attentional training, in which the training contingencies were eliminated. Immediately after this ABM procedure, participants were exposed to a stressor task in which they were asked to take 13 evenly spaced steps towards a live spider, enclosed within a clear container. After each step taken, they were required to rate their level of state anxiety on a scale ranging from 0 (Not at all anxious) to 100 (Worst anxiety you can imagine). Thus, like previous research, we were able to examine whether participants in the two attentional training conditions approached the spider to a differing degree. However, of critical importance, we now were also able to determine whether or not participants in the two ABM conditions exhibited different rates of change in state anxiety as they approached the spider, as would be predicted if biased attentional responding to spider stimuli causally contributes to this emotional component of spider fear.
METHOD
Participants
Approximately 800 introductory psychology students at the University of Western Australia were screened for spider fear using the FSQ (Szymanski & O'Donohue, Citation1995). To avoid ceiling and floor effects in terms of spider fear symptomatology, we sought participants who were mid‐range in terms of spider fear, and so only students who scored in the middle 50% of the FSQ scores (i.e., within the second and third quartiles; 10–35) were targeted for recruitment. Forty such participants accepted an invitation to participate in the study, in exchange for course credit. Participants were randomly allocated to one of two groups (n = 20 in each case). One group of participants (9 female and 11 male) was exposed to a training contingency designed to induce a relative reduction in attentional bias to spider information, and was designated the ‘avoid spider’ group. The other group of participants (13 female and 7 male) was instead exposed to a training contingency designed to induce a relative increase in attentional bias to spider information, and was designated the ‘attend spider’ group. This gave rise to a between‐group factor of training condition (avoid spider training vs attend spider training). The two groups did not differ in terms of FSQ scores (avoid spider condition: M = 20.45, SD = 7.70 vs attend spider condition: M = 22.10, SD = 7.57), t (38) = .68, ns, gender ratio, χ2 (1, n = 40) = 1.62, ns, or age (avoid spider condition: M = 18.30, SD = 1.17 vs attend spider condition: M = 17.85, SD = 1.76), t (38) = .95, ns.
Fear of Spiders Questionnaire
The FSQ was employed to select participants for the present study (FSQ; Szymanski & O'Donohue, Citation1995). The FSQ comprises 18 statements, each descriptive of people with heightened fear of spiders (e.g., ‘spiders are one of my worst fears’). Respondents are instructed to rate each item on an 8‐point Likert scale ranging from 0 (totally disagree) to 7 (totally agree). Total scores on the FSQ range from 0 to 126, with higher scores indicating greater levels of spider fear. The FSQ has been shown to have high internal consistency, test–retest reliability (Muris & Merckelbach, Citation1996), and good convergent validity (Szymanski & O'Donohue, Citation1995).
Experimental hardware
A HP Compaq dc 7800 Small Form Factor PC (Hewlett‐Packard Company, Palo Alto, CA, USA) with a 1680 × 1050-pixel resolution, 22‐inch colour monitor, a standard keyboard, and a two button mouse were used to deliver the attentional probe task.
Experimental stimuli
In previous research, images pairs each comprising one spider image and one non‐spider image (e.g., butterfly) have been employed in the attentional probe task. A potential problem with this approach is that participants in the avoid spider ABM condition may learn only to attend to butterflies (rather than to avoid spiders), and, conversely, that participants in the attend spider ABM condition may learn only to avoid butterflies (rather than attend to spiders). One way to circumvent this problem is to pair spider images and butterfly images not with each other, but with abstract images that have no representational content. In such a design, the attend spider ABM condition encourages participants to consistently attend towards spiders while equally often attending to and avoiding the abstract images that accompany representational image, whereas the avoid spider training condition encourages them to consistently avoid spiders, again while equally often attending to and avoiding the abstract images that accompany representational images. Hence, in this study, we employed stimulus pairs that each comprised one representational image (either a spider or butterfly) and one abstract image.
The present attentional probe task made use of 512 such pairs of images. One member of each pair was a representational image and the other was a non‐representational image. The representational images (n = 256) were photographs of spiders (n = 128) or butterflies (n = 128), whereas the non‐representational images (n = 256) were images of abstract art. All of the images used in this study were downloaded from publically available sources, following a Google Image Search. Using Adobe Photoshop, each image was cropped into a square shape (350 × 350-pixels), and converted to bitmap format.
Experimental tasks
Attentional probe task
Each trial began with a fixation display, followed 1200-ms later by the presentation of one of the image pairs for 500-ms, on alternate sides of the screen. Each member of the image pair measured 80 × 80-mm, and the centres of the left and right images were positioned 115-mm to the left and right of the screen centre, respectively. The representational member of the image pair appeared with equal frequency in either screen location. The display was then cleared, and a target probe immediately appeared in one of these two screen regions. This target probe was a red line measuring 1-cm, oriented either horizontally or vertically. Participants were required to make a discriminative response based on the orientation of this line, using the mouse to respond. The latency and accuracy of this probe discrimination response were recorded. The screen was then cleared, and the next trial commenced after a 1000-ms inter‐trial interval. In total, 448 trials were presented across the attentional probe task. The first 32 and final 32 trials were attentional assessment trials, whereas the intervening 384 trials were attentional training trials. The distinction between these blocks of trials was as follows:
Attentional assessment trials
In the attentional assessment trials, probes appeared in the locus of a previously presented representational image, and in the locus of a previously presented non‐representational image, with equal frequency, for all participants. The probe discrimination latencies obtained from these attentional assessment trials were used to compute an Attentional Preference for Spider Images Index. Specifically, selective attention to the spider images was revealed by disproportionate speeding to discriminate probes appearing in the locus of the representational images, compared to probes in the locus of the abstract images, when these representational images were spiders rather than butterflies (for a conceptually similar approach, see Basanovic & MacLeod, Citation2016). The Attentional Preference for Spider Images Index can be formally expressed as follows:
Index of Attentional Preference for Spider Images = (probe discrimination latencies for target probes in locus opposite to spider image − probe discrimination latencies for target probes in same locus as spider image) − (probe discrimination latencies for target probes in locus opposite to butterfly image − probe discrimination latencies for target probes in same locus as butterfly image).
Greater attentional preference for the spider images compared to butterfly images will be revealed by higher scores on this Attentional Preference for Spider Images Index.
Attentional training trials
In the attentional training trials, the position in which probes appeared depended upon the group the participant was assigned to. For participants in the attend spider group, whenever the representational member of the image pair was a photograph of a spider the probe always appeared in the same locus as the spider image, whereas for trials on which the representational member of the image pair was a photograph of a butterfly the probe always appeared in the locus opposite to this image. In contrast, for participants in the avoid spider group, whenever the representational member of the image pair was a photograph of a spider the probe always appeared in the locus opposite to this image, whereas for trials on which the representational member of the image pair was a photograph of a butterfly the probe always appeared in the same locus as this image.
In vivo exposure task
In this in vivo exposure task (IET), participants were asked to approach a live spider, enclosed within in a clear plastic container. The spider was an Australian Huntsman spider (Iospedella flavida), 5-cm in length. A room was set up with the clear container housing the spider being placed on a table, on the opposite side of the room from where the participant was required to enter. Thirteen steps (0 = no approach; 13 = full approach), each separated one foot (30.48-cm) in distance, were marked out on the floor leading up to the spider. The final step, if taken, involved the participant picking up the container with the spider in it. After each step was taken, the participant was required to rate their anxiety on a scale ranging from 0 (Not at all anxious) to 100 (Worst anxiety you can imagine). If the participant indicated that they did not want to approach the spider any closer, they were given 5 s to confirm their decision. Hence, in keeping with previous research, we were able to compute a behavioural approach score for each participant, reflecting the number of steps taken by the participant. However, of critical importance, we were also able to assess the rate of elevation in state anxiety as proximity to the spider increased, for participants in the two attentional training conditions, thus enabling us to evaluate the impact of ABM on this emotional component of spider fear.
Self‐reported Expectancy Assessment Task (SEAT)
To determine whether any observed effects may potentially have been the result of experimenter demand and/or participant expectancy, we asked the participant prior to actually approaching the spider to report whether they anticipated being able to take each step, and how anxious they expected they would be after doing so. Specifically, for each of the 13 steps they were asked ‘Can you imagine yourself moving up to Step x’, and were required to respond with a ‘Yes’ or ‘No’ answer. For each of the imagined steps, they were asked to report how anxious they expected they would feel at that particular step, using the same rating scale as employed when participants actually approached the spider. From these data, we were able to compute and analyse the participants’ self‐reported expectancies concerning both their expected behavioural approach to the spider, and the degree to which they anticipated that their subjective anxiety would increase as a function of their increasing proximity to the spider.
Procedure
Each participant was individually tested in a sound attenuated room. The session began with the participant reading an approved information sheet, and signing a consent form. They were then given instructions for the SEAT and IET. For the latter, the instructions emphasised that the participant should stop approaching the spider only if they believed they could not approach any closer. Following completion of the pre‐attentional training SEAT and IET, the participant was briefed about the requirements of the attentional probe task. They were instructed to discriminate the identity of the probe as quickly as possible, and to respond without delay as soon as probe was accurately identified. A brief practice period (16 trials) was completed before the attentional probe task was delivered. Upon completion of the attentional probe task, the post‐attentional training SEAT and IET were administered. At the end of the session, the participant was debriefed as to the purpose of the study, and thanked for their participation.
RESULTS
Impact of attentional training procedure on attentional preference for spider images
Probe discrimination accuracy on the attentional assessment trials, delivered before and after the attentional training trials, was high (M = 93.95%, SD = 4.78%), indicating good compliance with task instructions. Accuracy rates did not differ for participants in the two attentional training conditions, F(1, 38) = 0.97, p = .33, η p 2 = .02. To determine whether the attentional training procedure induced the required group difference in attentional responding to spider images, we first computed each participant's mean latency to accurately discriminate probes in each experimental condition, after eliminating outlying latencies that fell more than 1.96 standard deviations from the participant's mean latency for that experimental condition. This led to the exclusion of 2.96% of latencies. Using these mean latencies, we then computed the Attentional Preference for Spider Images Index score for each participant, as described in the Method section, and these are shown in Table .
Table 1. Mean and standard deviation (in parentheses) of Attentional Preference for Spiders Images Index scores, obtained from the attentional assessment trials when delivered before, and after, the attentional training trials
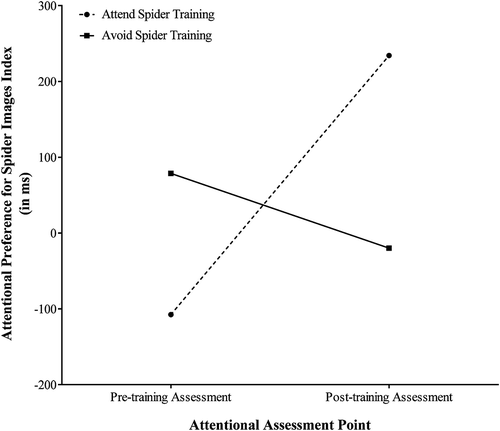
These attentional preference index scores were subjected to a 2 × 2 analysis of variance (ANOVA) that considered the between‐group factor Training Condition (avoid spider vs attend spider), and the within‐group factor Attentional Assessment Point (pre‐training assessment vs post‐training assessment). The sole significant effect to emerge from the analysis was a two‐way interaction of Training Condition × Attentional Assessment Point, F(1, 38) = 7.65, p = .01, η p 2 = .17. Fig. 1 illustrates the nature of this significant two‐way interaction. As can be seen, participants in the two training conditions did not differ significantly in terms of their Attentional Preference for Spider Images scores immediately prior to the ABM training, t (38) = 1.34, p = .19 Cohen's d = .43 (attend spider training condition: M = −107.61, SD = 456.47 vs avoid spider training condition: M = 78.67, SD = 420.84), but did differ significantly in these scores after the ABM training, t (38) = 2.16, p = .04, Cohen's d = .70.Footnote1 The nature of this significant difference was that, following the attentional training, participants in the attend spider training condition displayed higher Attentional Preference Index scores than did participants in the avoid spider training condition (M = 234.22, SD = 453.44 vs M = −19.95, SD = 265.25, respectively). Thus, this pattern of effects confirms that the attentional training procedure induced a group difference in biased attentional responding to spider stimuli, as intended.
Impact of attentional training procedure on spider fear
Self‐reported expectations concerning behavioural avoidance of spider
The behavioural approach self‐reported expectation scores obtained by participants in the two attentional training conditions, when asked to imagine approaching the spider, are shown in the two uppermost columns of Table . These data were subjected to a 2 × 2 ANOVA that considered the between‐group factor of Training Condition (avoid spider vs attend spider) and the within‐group factor Self‐reported Expectancy Assessment Point (pre‐attentional training vs post‐attentional training). The analysis did not reveal any significant effects, and of most relevance, the two‐way interaction of Training Condition × Self‐reported Expectancy Assessment Point did not approach significance, F(1, 38) = 0.98, p = .33, η p 2 = .02. Thus, there was no evidence to suggest that the ABM training induced a group difference in self‐reported expectancies concerning anticipated behavioural approach to the spider.
Table 2. Mean and standard deviation (in parentheses) of behavioural approach expectation scores, and behavioural approach scores, respectively, obtained from the Self‐reported Expectancy Assessment Task, and the in vivo Exposure Task, when delivered before, and after, the attentional training procedure
Observed behavioural avoidance of spider
The behavioural approach scores obtained by participants in the two attentional training conditions, when performing the IET, are shown in the two lowermost columns of Table . These data were subjected to a 2 × 2 ANOVA that considered the between‐group factor Training Condition (avoid spider vs attend spider) and the within‐group factor in vivo Exposure Task Assessment Point (pre‐attentional training vs post‐attentional training). No significant effects emerged from the analysis. Of most relevance, the two‐way interaction of Training Condition and in vivo Exposure Task Assessment Point did not reach significance, F(1, 38) = 0.28, p = .60, η p 2 = .01. Thus, in keeping with previous research, there was no evidence to suggest that the induction of differential attentional responding to spider stimuli induced a group difference in actual behavioural approach to the spider.
Self‐reported expectations concerning elevation in state anxiety as function of proximity to spider
The expected anxiety mood scores given by participants in the two attentional training conditions, when asked to report how they anticipated they would feel at each step of approach towards the spider, are shown in Table . These data were subjected to a multilevel analysis that considered the between‐group factor Training Condition (avoid spider vs attend spider), the within‐group factor Self‐reported Expectancy Assessment Point (pre‐attentional training vs post‐attentional training), and Spider Proximity, which reflected the expected anxiety mood scores reported by participants as they imagined themselves approaching the spider. This analysis revealed a significant effect of Spider Proximity that was quadratic in nature, F(1, 67.03) = 7.55, p = .01, indicating that participants self‐reported the expectancy that they would experience increasing growth in state anxiety as they approached the spider. No other significant effects emerged from the analysis. Of critical importance, there was no evidence that this self‐reported expectancy, concerning the growth of anxiety as spider proximity increased, was affected by training condition, as the higher order interaction involving Training Condition × Self‐reported Expectancy Assessment Point did not approach significance, F(1, 993.03) = .53, p = .47. Thus, there was no evidence to suggest that the ABM training induced a group difference in self‐reported expectancies concerning the rate at which anxiety would become elevated as proximity to the live spider increased.
Table 3. Mean and standard deviation (in parentheses) of expected anxiety mood scores obtained from Self‐reported Expectancy Assessment Task, when delivered before, and after, the attentional training procedure.
Experienced elevations in state anxiety as function of proximity to spider
The anxiety mood scores, representing the actual emotional experience of participants after each step taken towards the spider, are shown in Table . The hypothesis under test predicts that, before the attentional training, the rate of elevation in the anxiety mood scores as proximity to the spider increases will not differ between participants assigned to the two ABM conditions. However, after the attentional training, the rate of elevation in the anxiety mood scores as proximity to the spider increases will be less steep in those participants trained to attend away from spiders compared to those participants trained to attend towards from spiders. To test this hypothesis, we carried out a multilevel analysis that considered the between‐group factor Training Condition (avoid spider vs attend spider), the within‐group factor in vivo Exposure Task Assessment Point (pre‐attentional training vs post‐attentional training), and Spider Proximity, which reflected the mean anxiety mood scores indexing the emotional experienced by participants as they approached the spider. This analysis revealed a significant effect of Spider Proximity that was quadratic in nature, F(1, 67.18) = 5.27, p = .02, reflecting increasing growth in state anxiety as participants approached the spider. Critically, however, this effect of Spider Proximity on experienced anxiety was further modified by training condition, as evidenced by a significant high‐order interaction involving Training Condition × in vivo Exposure Task Assessment Point, F(1, 986.11) = 7.94, p = .005, which was the only other significant effect to emerge from the analysis.
Table 4. Mean and standard deviation (in parentheses) of anxiety mood scores obtained from the in vivo exposure task, when delivered before, and after, the attentional training procedure
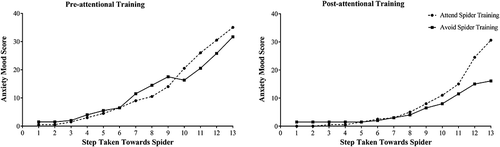
Consistent with the hypothesis under test, this higher order interaction, which is displayed in Fig. 2, resulted from the fact that, prior to delivery of the attentional training, the elevation of state anxiety as a function of increasing proximity to the spider did not differ between the groups assigned to the two ABM conditions, as reflected by a non‐significant simple two‐way interaction of Training Condition × Spider Proximity, F(1, 40.28) = .002, p = .97. However, after the attentional training, the elevation of state anxiety as a function of increasing proximity to the spider did come to differ between these two groups, as reflected by a simple two‐way interaction of Training Condition × Spider Proximity, F(1, 42.28) = 2.72, p = .05, which fell right on the border of statistical significance. Specifically, subsequent to the attentional training manipulation, the rate of elevation in anxiety mood scores as a function of increasing proximity to the spider was less steep for participants who had completed the avoid spiders ABM condition (steepness indicated by second degree parameter in quadratic function = .17), than was the case for participants who had completed the attend spider ABM condition (steepness indicated by second degree parameter in quadratic function = .34). Thus, this pattern of findings indicates that participants who had been exposed to the training contingency designed to attenuate attentional bias to spider stimuli subsequently came to display less pronounced elevations in state anxiety as their proximity to the spider increased than did participants who had been exposed to the training contingency designed to increase attentional bias to spider stimuli.
DISCUSSION
In this study, we delivered an ABM procedure closely based on those employed in previous research (Luo et al., Citation2015; Reese et al., Citation2010; Van Bockstaele et al., Citation2011), which was configured to induce attentional avoidance of, or attentional vigilance for spider information. The efficacy of this ABM procedure was determined by delivering 32 attentional assessment trials immediately after its completion. Like prior work, which has delivered as many as 96 assessment trials (Reese et al., Citation2010; Van Bockstaele et al., Citation2011), and as few as 20 assessment trials (Luo et al., Citation2015), we found that our ABM procedure was capable of inducing a group difference in attentional responding to spider information, suggesting that ABM procedures designed to modify such attentional selectivity can reliably do so. The induction of such differential attentional responding to spider information did not subsequently impact patterns of behavioural approach to a spider, but did influence the degree to which state anxiety became elevated as proximity to this spider increased. Thus, indicating that variability in attentional selectivity to spider information may not make a causal contribution to individual differences in the behavioural component of spider fear, but can causally contribute to individual differences in the emotional component of spider fear, reflected by the rate at which anxiety rises as a function of approaching a spider. This pattern of findings highlights the power of the ABM approach to illuminate the causal contributions made (or not made) by biased patterns of attention selectivity to different components of emotional disposition.
It is possible that the current experimental design may even have led to an underestimation of the impact of ABM on the rate of elevation in anxiety mood scores as a function of increasing proximity to a spider. This is because participants were required to behaviourally approach a spider before the ABM manipulation. It is well known that exposure to a feared stimulus can attenuate anxious responding to that stimulus (Meulders, Van Daele, Volders, & Vlaeyen, Citation2016). Thus, to the extent that participants habituated to the spider during this initial exposure, this would have reduced anxious responding observed when participants were required to behaviourally approach a spider after the ABM manipulation, potentially resulting in underestimation of the impact of ABM on the rate at which anxiety rises as proximity to a spider increases. We suggest that future researchers motivated to replicate the present findings should consider requiring participants to behaviourally approach the spider only once, after the ABM manipulation, to reduce the prospect of attenuating fear responses through habituation.
This findings represent the first support for the hypothesis that biased attentional responding to spider information can causally contribute to the emotional component of spider fear, at least within a non‐clinical population. As noted in the introduction, existing research has not yet adequately tested this hypothesis, as no previous study has assessed the impact of ABM on the rate at which state anxiety elevates as a function of proximity to spiders. Instead prior research has assessed only whether an ABM manipulation can induce a group difference in self‐reported state anxiety at a single assessment point during spider approach (Reese et al., Citation2010), or influences average levels of state anxiety experienced across the full duration of the extended spider approach procedure (Luo et al., Citation2015; Van Bockstaele et al., Citation2011). Such assessment approaches are unlikely to sensitively capture the type of group difference, in the rate of elevation in state anxiety as proximity to a spider increases, which was found to result from the ABM manipulation in this study. The insensitivity of these alternative emotional assessment approaches to the presently obtained effect can readily be demonstrated by subjecting our own anxiety data to these same analytical approaches. When our current anxiety data are analysed using the same approaches as previous research, we likewise find no evidence of ABM induced group differences in these alternative indices of anxious responding. Specifically, when following the approach adopted by Reese et al., who tested for an ABM induced group difference in state anxiety reported at the level of spider approach obtained post‐attentional training that corresponded to the highest level of spider approach obtained pre‐attentional training, we find no evidence of a group difference in state anxiety at this single assessment point, F(1, 37) = 2.40, p = .13. Likewise, when using the approach adopted by Van Bockstaele et al. and Luo et al., we find no evidence of an ABM induced group difference in average state anxiety experienced across the full duration of the spider approach procedure, F(1, 38) = .48, p = .50. Thus, not only do our data serve to confirm that training induced difference in attentional responding to spider information does give rise to a corresponding difference in the rate at which anxiety is increased by behavioural approach to spiders, but they also illustrate that previously adopted approaches to the analysis of anxiety scores collected during behavioural approach tasks do not adequately capture this emotional consequence of ABM.
It has been pointed out in some previous reviews of the ABM literature that anxiety changes resulting from ABM manipulations could, in principle, represent demand or expectancy effects (MacLeod, Koster, & Fox, Citation2009). Hence, we took care in this study to assess whether or not our current ABM manipulation exerted an impact on participants’ self‐reported expectances concerning the anxiety responses they would experience during the behavioural approach task. Our findings showed that, following the ABM training, participants in the two attentional training conditions did not differ in terms of these self‐reported expectancies, but did differ in terms of their anxiety mood scores when required to actually approach the spider. This difference between participants expected versus actual anxiety mood scores is important, as it mitigates against the possibility that demand or expectancy effects can account for the observed group difference in anxious responding during the behavioural approach task. While excluding the possibility of expectancy and demand effects is highly desirable in ABM studies, there are risks associated with this presently adopted approach. Specifically, it is possible that asking participants to initially report the degree to which they expect their state anxiety will rise as a function of increasing proximity to a spider, prior to them actually approaching it, may obscure genuine effects of ABM on anxious responding, if participants tend to stick with their self‐reported expectations when reporting their actual anxious experience during the behavioural approach task. This was not an issue in this study, as participants’ reported anxious experience during the approach task differed as a function of ABM condition, despite no such difference being evident on their expected anxiety. Nevertheless, we encourage future researchers to adopt other methods of reducing the potential influence of expectancy and demand effects, which potentially may compromise self‐report measures of anxiety. One such approach would be to employ physiological measures of anxious responding, such as heart rate variability or skin conductance (Schartau, Dalgleish, & Dunn, Citation2009), to assess the influence of ABM on arousal increase during spider approach. The data obtained from such measures could provide important converging support for the conclusions presently drawn on the basis of participant self‐reported anxious mood state.
In keeping with previous research (Luo et al., Citation2015; Reese et al., Citation2010; Van Bockstaele et al., Citation2011), we did not find any support for the hypothesis that biased attentional responding to spider stimuli causally contributes to the behavioural component of spider fear (i.e., avoidance of spiders) in the present study. Of course, one potential implication is that this hypothesis is invalid. However, before drawing the strong conclusion that attentional selectivity exerts no impact on behavioural facets of spider fear, it is appropriate to reflect upon the experimental methodologies hitherto employed to test this hypothesis. In this study, almost all participants reached the final step of the behavioural approach task, which involved them picking up a container with a live spider in it. Thus, it is possible that our present capacity to shed light on the hypothesis that biased attentional responding to spider stimuli causally contributes to the behavioural component of spider fear may have been compromised by ceiling effects. A more behaviourally intense approach task involving, for example, direct and prolonged physical interaction with a spider, may be a more powerful experimental method for addressing this hypothesis. It should also be noted that the present behavioural approach task, and previous such tasks, only have assessed a relatively narrow aspect of behavioural responding. Specifically, the behavioural approach task directly instructs participants to approach as spider as closely as they are able to, and so assesses only the ability to execute this specified behavioural objective. But in naturalistic situations, the patterns of behaviours exhibited by individuals are affected not only by behavioural abilities but also by behavioural preferences. Two individuals who have an equivalent ability to behaviourally approach a spider when instructed to so by an authority figure may have a very different tendency to do so when making their own unconstrained behavioural choices. It is possible that biased attentional responding to spider information may causally contribute to such behavioural choices. Testing this possibility within the laboratory would require amending behavioural assessment approaches to provide participants with alternative behavioural options, such as the option of approaching either of two glass boxes, only one of which contains a spider. A further limitation of the presently adopted approach is that only the immediate and direct impact of the ABM manipulation on behavioural approach was tested. It has long been argued that characteristic patterns of behavioural responding to a feared stimulus, such as avoidance, develop as a secondary consequence of the emotional response elicited by that stimulus (Mowrer, Citation1939; Rachman, Citation1977). Thus, it is possible that biased attentional responding to spider stimuli does functionally contribute to behavioural component of spider fear, but that this impact is indirect, mediated by the emotional response to spider proximity. If this is the case, then it might be expected that participants may need to be exposed to a spider on multiple occasions following ABM training, before the ABM elicited change in emotional responding to spider proximity translates into a change in degree of behavioural approach to a spider. We believe that future researchers should now seek to empirically test these possibilities, as work of this type could better reveal whether or not biased attentional responding to spider information makes a causal contribution to behavioural avoidance of spiders. For the moment, however, the present findings provide no support for the hypothesis that such attentional selectivity functionally contributes to behavioural approach/avoidance responses to spiders, but do support the hypothesis that attentional bias to spider information can causally influence the degree to which behavioural approach to spiders serves to elicit increased levels of anxiety.
Our present findings invite reconsideration of previous claims that ABM may have little potential as a therapeutic technique for attenuating high levels of spider fear (Luo et al., Citation2015; Reese et al., Citation2010; Van Bockstaele et al., Citation2011). Such claims have been based on the fact that prior studies did not assess the degree to which anxiety was elevated by spider proximity, and found no evidence on their alternative measures that the successful modification of attentional responding to spider stimuli significantly altered anxiety responses to spiders. Our present demonstration that the modification of such attentional selectivity alters the rate at which anxiety grows with increasing proximity to spiders suggests that the potential clinical applicability of such ABM should now be explored. For example, future researchers could seek to determine whether conventional cognitive behavioural therapy interventions for SP can be enhanced by the concurrent delivery of ABM. However, it will be important for future researchers addressing this issue to include a no‐ABM training condition. While resolving our present question did not require such a baseline condition, its absence prevents us from determining whether the ABM training condition configured to induce attentional avoidance of spider stimuli significantly attenuated the rate at which anxiety became elevated by increasing proximity to the live spider, or whether the ABM training condition configured to induce attentional vigilance for spider stimuli significantly increased the rate of anxiety elevation evoked by approaching the spider. Either or both training conditions may have made a contribution to the presently observed group difference, but the prospect of ABM for spider fear being of therapeutic benefit depends upon the validity of the former effect. Hence, future researchers should now employ a baseline no‐ABM training condition, to investigate whether the inducing attentional avoidance of spider information can attenuate the growth of anxiety elicited by spider approach, relative to this baseline condition, in participants selected on the basis of problematic levels of spider fear.
For the moment, the present findings demonstrate that variation in selective attentional responding to spider information can make a causal contribution to individual differences in the degree to which state anxiety becomes elevated as proximity to a spider increases, but does not appear to exert a direct and immediate causal impact on variability in the degree which individuals behaviourally approach a spider when instructed to do so. We hope that these findings, and in particular the new approach we have employed to assess the impact of ABM on the emotional component of spider fear, will be of value to future investigators seeking to better understand the contribution of attentional bias to spider fear.
ACKNOWLEDGEMENTS
This work was supported by Australian Research Council Grants DP140104448 and DP140103713, and by a grant from the Romanian National Authority for Scientific Research, CNCS–UEFISCDI, project number PNII‐ID‐PCCE‐2011‐2‐0045.
Additional information
Funding
Notes
1. Visual inspection of this significant two‐way interaction suggests the possibility that following ABM training, the degree to which participants in the attend spider ABM condition came to increase their attentional bias to spider information may have been greater than the degree to which participants in the avoid spider ABM condition came to reduce their attentional bias to spider information. However, statistical comparison indicated that there was no significant difference between the magnitude of the corresponding change in attentional bias evoked by each of these two ABM conditions, F (1, 38) = 2.33, p = .14, η p 2 = .06, which consequently can be considered equally effective in evoking their intended pattern of attentional change.
REFERENCES
- Bar‐haim, Y. , Lamy, D. , Pergamin, L. , Bakermans‐kranenburg, M. J. , & Van ijzendoorn, M. H. (2007). Threat‐related attentional bias in anxious and nonanxious individuals: A meta‐analytic study. Psychological Bulletin, 133(1), 1–24.
- Basanovic, J. , & Macleod, C. (2016). Does anxiety‐linked attentional bias to threatening information reflect bias in the setting of attentional goals, or bias in the execution of attentional goals? Cognition and Emotion. doi:https://doi.org/10.1080/02699931.2016.1138931
- Bradley, B. P. , Mogg, K. , White, J. , Groom, C. , & Bono, J. (1999). Attentional bias for emotional faces in generalized anxiety disorder. British Journal of Clinical Psychology, 38(3), 267–278. doi:https://doi.org/10.1348/014466599162845
- Broadbent, D. , & Broadbent, M. (1988). Anxiety and attentional bias: State and trait. Cognition and Emotion, 2(3), 165–183. doi:https://doi.org/10.1080/02699938808410922
- Grafton, B. , & Macleod, C. (2014). Cognitive bias modification training. In P. Emmelkamp & T. Ehring (Eds.), The Wiley handbook of anxiety disorders (pp. 1325–1342). Chichester, UK: John Wiley and Sons. doi:https://doi.org/10.1002/9781118775349.ch64
- Luo, X. , Ikani, N. , Barth, A. , Rengers, L. , Becker, E. , & Rinck, M. (2015). Attention bias modification in specific fears: Spiders versus snakes. Journal of Behaviour Therapy and Experimental Psychiatry. 49, 30–36. doi:https://doi.org/10.1016/j.jbtep.2015.04.006
- Macleod, C. , & Grafton, B. (in press). Anxiety‐linked attentional bias and its modification: Illustrating the importance of distinguishing processes and procedures in experimental psychopathology research. Behaviour Research and Therapy.
- Macleod, C. , Koster, E. H. , & Fox, E. (2009). Whither cognitive bias modification research? Commentary on the special section articles. Journal of Abnormal Psychology, 118(1), 89–99. doi:https://doi.org/10.1037/a0014878
- Macleod, C. , Mathews, A. , & Tata, P. (1986). Attentional bias in emotional disorders. Journal of Abnormal Psychology, 95(1), 15–20. doi:https://doi.org/10.1037/0021-843X.95.1.15
- Macleod, C. , Rutherford, E. , Campbell, L. , Ebsworthy, G. , & Holker, L. (2002). Selective attention and emotional vulnerability: Assessing the causal basis of their association through the experimental manipulation of attentional bias. Journal of Abnormal Psychology, 111(1), 107–123. doi:https://doi.org/10.1037/0021-843X.111.1.107
- Mathews, A. , & Macleod, C. (2005). Cognitive vulnerability to emotional disorders. Annual Review of Clinical Psychology, 1, 167–195. doi:https://doi.org/10.1146/annurev.clinpsy.1.102803.143916
- Merckelbach, H. , de Jong, P. J. , Muris, P. , & van Den hout, M. A. (1996). The etiology of specific phobias: A review. Clinical Psychology Review, 16(4), 337–361. doi:https://doi.org/10.1016/0272-7358(96)00014-1
- Meulders, A. , Van daele, T. , Volders, S. , & Vlaeyen, J. W. (2016). The use of safety‐seeking behavior in exposure‐based treatments for fear and anxiety: Benefit or burden? A meta‐analytic review. Clinical Psychology Review, 45, 144–156. doi:https://doi.org/10.1016/j.cpr.2016.02.002
- Mogg, K. , & Bradley, B. P. (2006). Time course of attentional bias for fear‐relevant pictures in spider‐fearful individuals. Behaviour Research and Therapy, 44(9), 1241–1250. doi:https://doi.org/10.1016/j.brat.2006.05.003
- Mogg, K. , Mathews, A. , & Eysenck, M. (1992). Attentional bias to threat in clinical anxiety states. Cognition and Emotion, 6(2), 149–159. doi:https://doi.org/10.1080/02699939208411064
- Mowrer, O. H. (1939). A stimulus‐response analysis of anxiety and its role as a reinforcing agent. Psychological Review, 46(6), 553–565. doi:https://doi.org/10.1037/h0054288
- Muris, P. , & Merckelbach, H. (1996). A comparison of two spider fear questionnaires. Journal of Behaviour Therapy and Experimental Psychiatry, 27(3), 241–244.
- Rachman, S. (1977). The conditioning theory of fear acquisition: A critical examination. Behaviour Research and Therapy, 15(5), 375–387.
- Reese, H. E. , Mcnally, R. J. , Najmi, S. , & Amir, N. (2010). Attention training for reducing spider fear in spider‐fearful individuals. Journal of Anxiety Disorders, 24(7), 657–662. doi:https://doi.org/10.1016/j.janxdis.2010.04.006
- Schartau, P. E. , Dalgleish, T. , & Dunn, B. D. (2009). Seeing the bigger picture: Training in perspective broadening reduces self‐reported affect and psychophysiological response to distressing films and autobiographical memories. Journal of Abnormal Psychology, 118(1), 15. doi:https://doi.org/10.1037/a0012906
- See, J. , Macleod, C. , & Bridle, R. (2009). The reduction of anxiety vulnerability through the modification of attentional bias: A real‐world study using a home‐based cognitive bias modification procedure. Journal of Abnormal Psychology, 118, 65–75. doi:https://doi.org/10.1037/a0014377
- Szymanski, J. , & O'donohue, W. (1995). Fear of spiders questionnaire. Journal of Behaviour Therapy and Experimental Psychiatry, 26(1), 31–34. doi:https://doi.org/10.1016/0005-7916(94)00072-T
- Van bockstaele, B. , Verschuere, B. , Koster, E. H. , Tibboel, H. , De houwer, J. , & Crombez, G. (2011). Effects of attention training on self‐reported, implicit, physiological and behavioural measures of spider fear. Journal of Behaviour Therapy and Experimental Psychiatry, 42(2), 211–218. doi:https://doi.org/10.1016/j.jbtep.2010.12.004
- Vrijsen, J. N. , Fleurkens, P. , Nieuwboer, W. , & Rinck, M. (2009). Attentional bias to moving spiders in spider fearful individuals. Journal of Anxiety Disorders, 23(4), 541–545. doi:https://doi.org/10.1016/j.janxdis.2008.11.002