Abstract
High Na stress induces deleterious effects on plants by ion toxicity, inadequate water status and nutrient imbalances. Exogenous Ca ions that can sustain a high absorption rate of both Ca and K against Na in plants improve plant growth. To examine the effect of adequate internal water status on the process of cation absorption, water spray treatments were compared with Ca addition treatments at high (40, 80 and 120 mol m−3) NaCl concentrations. The water spray treatments improved internal water status and led to a higher Ca accumulation (73.2–85.2% increase) in the shoots than Na treatments at the same external Na and Ca concentrations. Higher Ca contents of shoots in the water spray treatments than in the corresponding Na treatments were induced by the higher transpiration stream concentration factor of Ca. Examination of plants with an adequate water status in shoots revealed that the active absorption of Ca was sensitive to the internal water status, and that there was no antagonistic effect of Na on Ca. It was necessary for the NaCl-salinized plants that Ca be actively absorbed and transported to avoid salinity damage, which was achieved through adequate water status.
INTRODUCTION
Approximately 7% of the land surface of the earth and over 50% of irrigated land is affected by salinity (CitationFlowers et al. 1997). Salinity stress reduces the yield of most crop species by disturbing the plant metabolism as a result of water deficit, ion toxicity and nutrient imbalances (CitationPasternak 1987; CitationWerner and Finkelstein 1995). In the cytoplasm, high Na accumulation impairs K nutrition and inhibits the activity of many enzymes (CitationEpstein 1972; CitationMurguia et al. 1995; CitationWu et al. 1996). Therefore, attempts have been made to identify the physiological and biochemical processes that are affected by NaCl in order to modify them to increase salt tolerance (CitationApse and Blumwald 2002). In addition, the maintenance of a high cytosolic K/Na ratio is also considered to be important (CitationMaathuis and Amtmann 1999). These K transport steps are sensitive to Ca concentrations in the cell. Calcium is required to maintain or enhance selective K absorption by plants at high Na concentrations. Studies have shown that in wheat and many other plant species inhibition of the initial Na influx was directly correlated with the Ca-induced decrease in shoot Na accumulation and with the alleviation of growth inhibition by Na (CitationCramer 2002). In wheat, the inhibition of unidirectional Na influx by the addition of external Ca exerted a more significant effect on the Na concentration of shoots than roots (CitationReid and Smith 2000). Increasing the Ca concentration significantly decreased the shoot Na concentration, along with the increase of the K/Na ratio and decrease of the Na/Ca ratio (CitationEssah et al. 2003). However, because the internal Ca status cannot be manipulated and monitored, most experiments conducted to reveal the role of Ca in K/Na transport deal with the evaluation of the absorption of ions at various concentrations of K, Na and Ca in the medium in which the plant roots are located.
In the present study, we investigated the effects of internal water status on the growth of NaCl-salinized rice plants in relation to the absorption and transport of cations in the presence of 40, 80 and 120 mol m−3 NaCl. The purpose of the present study was to elucidate the antagonistic effect of Na on K and Ca. To avoid the
Table 1 Composition of the treatment solutions
MATERIALS AND METHODS
Rice seeds (Oryza sativa L. cv. Yamabiko) were soaked in tap water. Soon after germination the seedlings were transferred to vinyl nets that were floated on water. The control nutrient solution was supplied at the second leaf stage. The hydroponic composition of the control (CO) solution was as follows: (mol m−3) 2.0 N (NH4NO3), 0.4 P (NaH2PO4), 2.0 K (KCl), 1.0 Ca (CaCl2·2H2O) and 2.0 Mg (MgSO4·7H2O) for the macronutrients and (g m−3) 2.0 Fe (FeSO4·7H2O), 0.5 Mn (MnSO4·6H2O), 0.2 B (H3BO3), 0.1 Zn (ZnSO4·7H2O), 0.01 Cu (CuSO4·5H2O) and 0.005 Mo ((NH4)6Mo7O24·7H2O) for the micronutrients. The pH of the culture solution was adjusted to 5.5 by the addition of a 1:1 H2SO4 solution. Seedlings at the five-leaf stage were transferred to 4 L plastic pots with 5 plants per pot. The CO solution was replaced every 2 days. The treatments listed in were initiated 12 days after the transfer. Most seedlings were at the six-leaf stage. Six pots were used for each treatment. In the H2O treatments, distilled water was sprayed on the shoots with a supersonic humidifier. The spray was carried out for 90 min every morning and evening to avoid the influence of differences in leaf temperature among the treatments. The shoots and roots were harvested at 8 days after the onset of the treatments. The harvested tops and roots were rinsed thoroughly with distilled water, dried at 60°C and the dry weight (DW) was determined. After digestion with an acid mixture (CitationSong and Fujiyama 1996, Citation1998), the Na, K and Ca contents were analyzed using atomic absorption spectrophotometry.
RESULTS AND DISCUSSION
It is necessary to increase the concentrations of osmotic solutes in cells to maintain water absorption and adequate internal water status in NaCl-salinized plants. Sodium chloride can effectively decrease the osmotic potential of cells. Sodium, however, displays an ion phytotoxicity effect and brings about nutrient imbalances. Exogenous Ca can sustain K transport and K–Na selectivity in Na-challenged plants (CitationLäuchli 1990). It is generally considered that Na exerts an antagonistic effect on Ca absorption and transport (CitationTester and Davenport 2003) and that Ca is absorbed by passive transport. To examine the effect of adequate internal water status on the process of cation absorption, water spray and Ca addition treatments at high (40, 80 and 120 mol m−3) NaCl concentrations were compared.
The cumulative transpiration (CT) and water status of the shoots gradually decreased with increasing Na concentration (). The Na40 treatment did not result in a decrease in DW. Dry weight decreased with increasing Na concentrations above 40 mol m−3 in solution. Na80-H2O and Na80-Ca treatments improved the growth of the plants. The H2O treatments ameliorated the internal water status in shoots. The content of K and proportion of K to K+Na in shoots and roots gradually decreased with increasing Na concentrations in the nutrient solution (, ). The H2O and Ca treatments decreased the proportion of Na and increased the Ca content in shoots (). The Ca treatments led to a marked decrease in the proportion of Na to K+Na in roots because of the direct effect on roots. It would be interesting to determine why the Ca content of shoots in the water spray treatments was higher than that in the corresponding Na treatments, although the difference in CT between both treatments was negligible. Simultaneously, the water spray treatments led to a higher transpiration stream concentration factor (TSCF) (CitationRussell and Shorrocks 1959) of Ca, and increased the selectivity of K versus Na and Ca versus Na (SK,Na[CitationPitman 1976] and SCa,Na, respectively) in the shoots (). As the Ca treatments supplied a sufficient amount of Ca for rice, the Ca content was markedly enhanced. However, the SCa,Na of the shoots and roots in the Ca treatments was lower than that in the Na treatments and less than half of that in the H2O treatments. Obviously, external Ca increased the Ca content, while selective Ca absorption decreased. It is important to note that the H2O treatments resulted in similar SK,Na values in the shoots as the Ca treatments. These results indicated that the adequate internal water status of the NaCl-salinized rice shoots was modified so that selective absorption and transport of Ca and selective K transport could be enhanced. It is necessary for the NaCl-salinized plants that Ca be actively
Figure 1 Proportion of Na or K content and calcium content of rice plants grown at various salt concentrations (40, 80, 120 mol m−3), and the addition of calcium (10 mol m−3) into NaCl-salinized nutrient solution or mist-water spray onto shoots in the salt treatments. a, shoots; b, roots. The proportion of Na and K is denoted by □ bars and ![]()
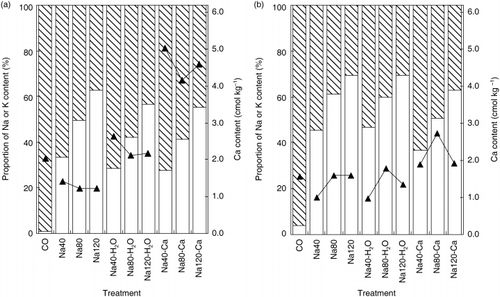
Table 2 Growth, water status and cumulative transpiration in rice 8 days after the beginning of the treatment
Although the external Na and Ca concentrations in the Na and H2O treatments were identical, the H2O treatments led to a higher Ca accumulation (73.2–85.2% increase) in the shoots. The contribution of increasing Ca content in the shoots was significantly higher than that of CT in the H2O treatments. There is strong evidence that the effect of Na on Ca did not result from the antagonistic interaction of these cations on the root surface. The decrease in the Ca concentration and the
Table 3 Transpiration stream concentration factor (TSCF-Na or TSCF-Ca) and K or Ca versus Na selectivity (SK,Na or SCa,Na) in rice 8 days after the beginning of the treatment
Shoot Na can only be returned to roots via the phloem. Evidence of extensive recirculation of shoot Na to roots is limited. Sodium transport is largely unidirectional to shoots (CitationTester and Davenport 2003). Leaves are more vulnerable than roots to Na, simply because Na accumulates and reaches higher concentrations in shoots than in roots (CitationTester and Davenport 2003). Exclusion of Na from the roots or translocation from shoots to roots is an important mechanism to minimize Na damage for some plants. Inward K channels in root protoplasts displayed a relatively high permeability to Na (CitationMaathuis and Sanders 1995). The reduction of Na accumulation could result from either a decrease in influx or an increase in efflux, at least in part, and Ca was able to decrease the unidirectional influx of Na into the roots (CitationTester and Davenport 2003). As Ca binding can increase the selectivity of plasma membrane K+ channels, the Na+ influx via these channels could be reduced (CitationJacoby and Hanson 1985; CitationRains and Epstein 1967). The SK,Na value of the shoots in the H2O treatments was similar to that in the Ca treatments. It is possible that the enhanced SCa,Na and adequate water status in the H2O treatments excluded Na from the shoots and roots, which resulted in a higher SK,Na value in the shoots. Reverse water-flow of plants was observed in wheat, corn, clover, tomato (CitationBreazeale et al. 1951), cotton (CitationBaker and Van Baver 1988) and sorghum (CitationXu and Bland 1993). Improvement in the water and nutritional status by the H2O treatments resulted in a higher DW in the Na80 treatment, unlike in the Na120 treatment in which Na injury was too severe.
Our findings suggest that adequate internal water status led to an increase in Ca transport and altered the Na influx or efflux rates because the shoot/root ratio of K in the H2O treatments (1.16/1.30) was higher than that (0.93/1.23) in the Ca treatments and vice versa for Na (0.52/0.75 in the H2O treatments and 0.61/0.90 in the Ca treatments). Salt-tolerant wild-type barley showed larger values for the contents of polysaccharides (39%) and Ca (double) than the cultivated-type plants when grown under high salt conditions (CitationSuhayda et al. 1994). The NaCl-salinized rice plants were able to maintain an adequate nutritional status by enhancing the transport of Ca and K within a short period of time (CitationBaba and Fujiyama 2003). In the present study, the results showed that this ability required adequate internal water status over a long period of time. The adequate internal water status that led to a larger reverse water-flow might, therefore, be important in excluding Na from shoots or roots.
We conclude that enhanced Ca absorption in the H2O treatments can be attributed to improvement in the internal water status in shoots. The antagonistic effect of Na on Ca did not take place in the treatments. The mechanism of Na and Ca absorption at high Na concentrations, which we proposed in this study and in which antagonism was not involved, should be added to the list of mechanisms that play a crucial role in protecting plants from damage.
REFERENCES
- Flowers , TJ , Garcia , A , Koyama , M and Yeo , AR . 1997 . Breeding for salt tolerance in crop plants–the role of molecular biology . Acta PhysiolPlant , 19 : 427 – 433 .
- Pasternak , D . 1987 . Salt tolerance and crop production. A comprehensive approach . AnnuRevPhytopathol , 25 : 271 – 291 .
- Werner , JE and Finkelstein , RR . 1995 . Arabidopsismutants with reduced response to NaCl and osmotic stress . PhysiolPlant , 93 : 659 – 666 .
- Epstein , E . 1972 . Mineral Nutrition of Plants: Principles and Perspectives , New York : Wiley .
- Murguia , JR , Belles , JM and Serrano , RA . 1995 . Salt-sensitive 3’(2’),5’-bisphosphate nucleotidase involved in sulfate activation . Science , 267 : 232 – 234 .
- Wu , SJ , Ding , L and Zhu , JK . 1996 . SOS1, a genetic locus essential for salt tolerance and potassium acquisition . Plant Cell , 8 : 617 – 627 .
- Apse , MP and Blumwald , E . 2002 . Engineering salt tolerance in plants . CurrOpinBiotech , 13 : 146 – 150 .
- Maathuis , FJM and Amtmann , A . 1999 . K+nutrition and Na+toxicity: the basis of cellular K+/Na+ratios . AnnBot , 84 : 123 – 133 .
- Cramer , GR . 2002 . “ Sodium–calcium interactions under salinity stress ” . In SalinityEnvironment–Plants–Molecules , Edited by: Läuchli , A and Lüttge , U . 205 – 227 . Dordrecht : Kluwer Academic Publishers .
- Reid , RJ and Smith , FA . 2000 . The limits of sodium/calcium interactions in plant growth . AustJPlant Physiol , 27 : 709 – 715 .
- Essah , PA , Davenport , R and Tester , M . 2003 . Sodium influx and accumulation in Arabidopsis . Plant Physiol , 133 : 307 – 318 .
- Läuchli , A . 1990 . Calcium, salinity and the plasma membraneCalcium in Plant Growth and Development , American Society of Plant Physiologists Symposium Series Edited by: Leonard , RT and Hepler , PK . vol4 , 26 – 35 . Rockville : American Society of Plant Physiologists .
- Tester , M and Davenport , R . 2003 . Na+tolerance and Na+transport in higher plants . AnnBot , 91 : 503 – 527 .
- Russell , RS and Shorrocks , VM . 1959 . The relationship between transpiration and the absorption of inorganic ions by intact plants . JExpBot , 10 : 301 – 316 .
- Pitman , MG . 1976 . “ Ion uptake by plant roots ” . In Transport in Plants, 2Part BTissues and Organs , Edited by: Lutge , U and Pitman , MG . 95 – 128 . Berlin : Springer-Verlag .
- Maathuis , FJM and Sanders , D . 1995 . Contrasting roles in ion transport of two K+-channel types in root cells of Arabidopsis thaliana . Planta , 197 : 456 – 464 .
- Jacoby , B and Hanson , JB . 1985 . Controls on 22Na+influx in corn roots . Plant Physiol , 77 : 930 – 934 .
- Rains , DW and Epstein , E . 1967 . Sodium absorption by barley roots: its mediation by mechanism 2 of alkali cation transport . Plant Physiol , 42 : 319 – 323 .
- Breazeale , EL , McGeorge , WT and Breazeale , JF . 1951 . Water absorption and transport by leaves . Soil Sci , 72 : 239 – 242 .
- Baker , JM and Van Baver , CHM . 1988 . Water transfer through cotton plants connecting soil regions of differing water potential . AgrJ , 30 : 993 – 996 .
- Xu , X and Bland , WL . 1993 . Reverse water-flow in sorghum roots . AgrJ , 85 : 384 – 387 .
- Suhayda , CG , Redmann , RE and Wang , XY . 1994 . “ Salinity alters root cell wall properties and trace metal uptake in barley ” . In Biochemistry of Metal Micronutrients in the Rhizosphere , Edited by: Manthey , JA , Crowley , DE Luster , DG . 325 – 342 . Boca Raton : Lewis Publishers .
- Baba , T and Fujiyama , H . 2003 . Differences in short-term responses of rice and tomato to sodium salinization and supplemental potassium and calcium . Soil SciPlant Nutr , 49 : 669 – 675 .
- Song , JQ and Fujiyama , H . 1996 . Ameliorative effect of potassium on rice and tomato subjected to sodium salinization . Soil Sci. Plant Nutr. , 42 : 493 – 501 .
- Song , JQ and Fujiyama , H . 1998 . Importance of Na content and water status for growth in Na-salinized rice and tomato plants . Soil Sci. Plant Nutr. , 44 : 197 – 208 .