Abstract
Viruses play important roles in biogeochemical nutrient cycles and act as genomic reservoirs in marine and freshwater environments, the understanding of which brought about the so-called ‘third age’ of virus ecology in aquatic environments. Unfortunately, the third age is in oceanography and limnology and outside the soil world. The main reason why virus ecology in soils has shown less progress is that agronomical and epidemiological interests were the primary motivation of viral studies by soil microbiologists. In this review, past research on viruses in soils is summarized after the introduction of the ecological traits of viruses, which are the effects of viruses on beneficial bacteria and soil-borne plant pathogens, adsorption of viruses to soils, and soil factors affecting viral inactivation and survival in soils. Horizontal gene transfer (transduction) in soils is also reviewed. Second, the abundance of viruses and their roles in biogeochemical nutrient cycles are summarized in aquatic environments. Five to 25% of the carbon fixed by primary producers is estimated to enter into the microbial loop via virus-induced lysis at different trophic levels in aquatic environments. The diversity of virus communities in aquatic environments estimated from analyses of the frequency distribution of capsid sizes and the morphology of virus populations are reviewed, and recent findings on the genomic diversity of viruses and their roles as the greatest genomic reservoirs in aquatic environments follow in the subsequent section. Viral genomics is elucidating the viral diversity and phylogenetic relationships among viruses in different environments. As the soil environment is a more diverse habitat for viruses than aquatic environments, viruses in soils have great potential to play roles comparable in quantity, which are unique in quality, to those in aquatic environments. Therefore, the potentiality and characteristics of viruses in soils are discussed in the final section for future research on virus ecology in soils from the viewpoints of biogeochemistry and genomic diversity. Synecological approaches to viruses in soils may open up a new era of soil virus ecology.
INTRODUCTION: THE THIRD AGE OF VIRUS ECOLOGY
Viruses exist wherever life is found in the environment. Viruses are the most abundant biological entities on Earth, with an estimated total population size of > 1030 viruses in the ocean alone (CitationBreitbart and Rohwer 2005; CitationSuttle 2005). Known as therapeutic tools before the 1940s and model systems for modern biology before the 1970s (CitationWilhelm and Suttle 1999), viruses are increasingly recognized as a major driving force of global biogeochemical nutrient cycles (CitationFuhrman 1999), a key factor influencing microbial diversity (CitationMühling et al. 2005), the greatest genetic reservoirs on Earth (CitationFrost et al. 2005; CitationPaul and Sullivan 2005; CitationWeinbauer and Rassoulzadegan 2004), and mediators of horizontal gene transfer (CitationBreitbart et al. 2004b; CitationMillard et al. 2004). These understandings have led to a resurgence of interest in virus ecology over the past decades, the so-called ‘third age’ of virus ecology (CitationMann 2005). Unfortunately, the third age is in oceanography and limnology and outside the soil world!
To the best of our understanding, soil microbiologists have consistently been indifferent to the progress of virus ecology on global perspectives in marine and freshwater environments, and they have conducted viral studies from agronomic and epidemiological concerns. In this review, the ecological traits of viruses are introduced, followed by a summary of past research on viruses in soils. This encompasses the effects of viruses on beneficial bacteria and soil-borne plant pathogens, adsorption of viruses to soils, soil factors influencing viral inactivation and survival in soils, and horizontal gene transfer in soils. The abundance of viruses and their roles in biogeochemical nutrient cycles are then considered in relation to aquatic environments. The diversity of viruses and their importance as genetic reservoirs in aquatic environments are then explained with comments on the conceivable contributions of viruses in soils in relation to these matters. Equivalent numbers of bacteria exist in soils to those in the sea, and are densely populated in soils. The soil environment is a more diverse habitat for viruses than aquatic environments. Therefore, viruses in soils have great potential to play roles comparable in quantity, but unique in quality, in those issues. Potentiality and characteristics of viruses in biogeochemical nutrient cycles and as genomic reservoirs in soils are discussed in the final section for future research on virus ecology in soils from the viewpoints of biogeochemistry and genomic diversity.
This review covers broad topics on virus ecology in soils and aquatic environments and many excellent reviews have been published on the respective topics (see ).
Table 1 Books, reviews and primer articles that deal with viruses/phages in the environment in relation to the topics of this review
VIRAL LIFE IN THE ENVIRONMENT
What are viruses?
Viruses cannot capture and store free energy and they are not functionally active outside their host cells. In one sense viruses are not living organisms, and they occupy a unique position in biology. A virus has intrinsic properties (e.g. its size) and relational properties (e.g. its host), the second type of property existing only by virtue of a relationship with other objects (CitationVan Regenmortel 2000).
Bacteriophages (phages) are estimated to comprise the majority among viruses in aquatic environments from circumstantial evidence, such as the greater abundance of bacteria over that of other planktonic hosts, the lack of a significant correlation between algal biomass and viral abundance, the predominance of viruses within the virioplankton with phage-sized genomes, and the ability of changes in bacterial abundance to predict changes in viral abundance (CitationWommack and Colwell 2000). Between 1 and 4% of visibly infected bacterial cells and more than 10 virus particles released per cell on bacterial cell lysis (burst size) support this inference (CitationWommack and Colwell 2000). The findings that phages comprise the majority among viruses in aquatic environments cannot simply be extended to soil environments because fungal biomass is often larger than bacterial biomass in soils (CitationAnderson and Domsch 1980).
Phages are classified by morphotype and host genus. The six basic phage types, named A to F, were first defined by CitationBradley (1967) using gross morphology and the nature of their nucleic acid. Phages are presently divided into 21 morphotypes, and tailed phages are currently deemed to constitute the order Caudovirales (CitationAckermann 1998, Citation2001, Citation2003). Tailed phages are grouped into Myoviridae with a contractile tail, Siphoviridae with a long non-contractile tail and Podoviridae with a short non-contractile tail (CitationFauquet et al. 2005), among which Siphoviridae or phages with long, non-contractile tails comprise 61% of the tailed phages that have been examined using transmission electron microscopy (TEM) since 1959 (CitationAckermann 1998, Citation2001, Citation2003). Phages belong to 13 virus families and occur in over 140 bacterial genera. A recently developed genome-based taxonomy for phages divided Podoviridae into two groups of T7-like and PZA-like podophages (CitationRohwer and Edwards 2002).
Lifestyle of viruses
Viruses perform either lytic, lysogenic or pseudolysogenic infection. A temperate phage in the lysogenic state is referred to as a prophage and lysogeny involves integration of the phage genome into the host cell chromosome as a prophage. The production of a temperate phage in the lysogenic state is independent of host cell density. In contrast, a virulent virus in the lytic state redirects the host metabolism toward the production of new viruses, resulting in lysis of the cells. Production and survival of a virulent virus requires frequent, successful host–phage encounters to exceed the rate of phage destruction and inactivation (CitationWeinbauer and Suttle 1996). Lysogeny is an effective strategy to maintain viral populations when host abundance is too low and/or where hosts are inactive for viral populations to be maintained by lytic infection (CitationChibani-Chennoufi et al. 2004; CitationMann 2003; CitationSmit et al. 1996; CitationWeinbauer and Suttle 1996; CitationWeinbauer et al. 2003; CitationWilliamson et al. 2002). Myoviruses are typically lytic and often have a broader host range than other tailed phages in marine environments, whereas many siphoviruses can achieve their genomes in host cells, tying their replication rate to that of the host until an environmental cue triggers the lytic cycle, and CitationSuttle (2005) grouped the former as r-strategists (short generation times and high reproductive rates) and the latter as K-strategists (longer generation times and lower reproductive rates) from their lifestyles.
Although virulence is a common strategy for viruses in marine and freshwater environments, a greater incidence of lysogeny was noted for bacterial strains isolated from oligotrophic marine environments (CitationJiang and Paul 1994, Citation1998). In contrast, lysogeny in soil has a distinct advantage over virulence for phage and host survival where long periods of host inactivity can be survived by the residence of phages within host populations (CitationMarsh and Wellington 1994; CitationPantastico-Caldas et al. 1992). Viruses are lysogenic in certain strains of hosts and purely lytic in others.
A pseudolysogen is a phage-infected cell that grows and divides even though the virus is pursuing a lytic infection, where the nucleic acid of the phage resides within its starved host in an unstable, inactive state. CitationRipp and Miller (1997) found that the resolution of the pseudolysogenic state depended on nutrient addition, resulting in either the establishment of true lysogeny or the initiation of the lytic cycle. As cells become more starved, the frequency of pseudolysogens increases (CitationRipp and Miller 1998). CitationCampbell et al. (1995) isolated five virulent phages of a fluorescent Pseudomonas strain from barley rhizosphere, but they grew very slowly in the host cells, resulting in their co-occurrence in the soil environment. Microorganisms in soils commonly exist in a starved state, but no study has been conducted to examine the pseudolysogenic state of microorganisms in this habitat.
Coexistence of bacteria and their phages in the environment
The coexistence of bacteria and their phages in the same habitats is a common occurrence and is explained using the theory of coevolution. CitationKleczkowska (1945) observed that resistant hosts appeared readily in soil cultures of Rhizobium sp. with its phage. Furthermore, when aseptically grown clover was infected with a pure culture of a Rhizobium strain susceptible to the phage, resistant strains appeared spontaneously in the nodules in the absence of the phage. Experimentally, CitationLevin et al. (1977) demonstrated that there are stable states of coexistence for one resource, one prey and one predator system, as well as for one resource, two prey and one predator system using Escherichia coli K12 and its phage T2.
To account for the lack of elimination of bacterial prey or hosts by predators from terrestrial ecosystems, CitationAlexander (1981) proposed: (1) interactions among predators, (2) predators under biological control, (3) prey mutation to resistant forms, (4) refuge to safe microsites, (5) switching to a second prey or host species, (6) density dependence, (7) replication to compensate for killing. Coevolution of bacteria and phages leads to an endless “arms race” between resistant bacterial mutants and corresponding host-range phage mutants. The phage adsorption site on the bacterial surface is one of the battlefields in the arms race, where the bacterium and its phage race for the site modification and the adaptation to it quantitatively and qualitatively (CitationLenski 1984, Citation1988). However, the soil environment is resource limiting and CitationLenski and Levin (1985) demonstrated experimentally that bacteria resistant to T2 and T7 phages have marked disadvantages to sensitive bacteria under resource-limiting chemostat conditions because the modification of surface phage receptors changes the abilities of the bacterium to exploit its environment, in general, in less efficient directions (CitationLenski 1988; CitationLenski and Levin 1985). The work by CitationSzmelcman and Hofnung (1975) provided evidence of superiority of sensitive bacteria to resistant ones, in which E. coli mutants that are resistant to a λ-phage are unable to grow on maltose at low concentrations, although they can grow at higher concentrations. In marine environments, Synechococcus communities are believed to dominate because of cells resistant to their co-occurring phages, and these phages are maintained by scavenging on the relatively rare, sensitive cells in the communities (CitationWaterbury and Valois 1993). Spatial heterogeneity is another important factor in bacterial escape from phage infection in soil (CitationBurroughs et al. 2000).
Lysogeny may be a solution for the arms race in coevolution in various environments, and seems to be reflected in the fact that approximately 21–60% of member species are lysogens for most taxonomic groups of bacteria (CitationAckermann and DuBow 1987a). Bacterium–phage interaction consists of mutual inhibition, competition, amensalism and commensalism.
STUDIES ON SOIL VIRUSES
Many studies have been conducted on viruses in soils from the viewpoints of crop production and epidemiology. With either viewpoint, proliferation, inactivation, survival and the fate of specific viruses were the main concerns, for example, viruses infecting useful and undesirable microorganisms such as rhizobia and plant pathogens in agriculture and infectious to mammals in epidemiology. In other words, the main concerns were focused on the autecology of specific viruses in soils.
Agronomic concerns
For soil microbiologists, agronomic interests have continuously been the motivation for viral studies in soil since the 1970s. Many studies were conducted to elucidate rhizobiphage populations, the host range of rhizobiphages among Rhizobium spp., and the effects of rhizobiphages on the nodule formation and yield of leguminous plants (CitationAbebe et al. 1992; CitationHussein et al. 1994; CitationNovikova and Limeschenko 1992; CitationNovikova et al. 1993). CitationEl Didamony and Abdel-Fattah (1998) examined the effect of phage infection (lysogenic and lytic to rhizobia) on the growth stimulation of leguminous plants by the arbuscular fungus Glomus mosseae and two Rhizobium leguminosarum bv. viciae strains. Shoot dry weight and nodulation of faba bean plants inoculated with G. mosseae were significantly increased compared with non-mycorrhizal plants in the presence and absence of the phage of the R. leguminosarum strains. The number of nodules produced by the lysogenic rhizobial strain was significantly increased in mycorrhizal plants, but was still less than the number produced by the non-lysogenic strain. The levels of mycorrhizal colonization were markedly enhanced in the presence of the non-lysogenic rhizobial strain.
The effects of viruses on beneficial bacteria and soil-borne plant pathogens have also been extensively studied (CitationErskine 1973; CitationEzuka and Kaku 2000; CitationGross et al. 1991; CitationHashem et al. 1996; CitationKeel et al. 2002; CitationMcKenna et al. 2001; CitationToyoda et al. 1991). In general, phages are abundant in soils where bacterial hosts proliferate, such as rhizobiphages in the fields of legumes (CitationSharma et al. 2002). In relation to this context, it is noteworthy that no attention has been paid to soil phages infecting anaerobic bacteria, such as methanogens. As for phages of general soil bacteria, CitationAshelford et al. (1999, Citation2000) observed clear seasonal population dynamics of podophages and their host, Serratia liquefaciens, in sugar beet fields.
Bacterial blight caused by Xanthomonas oryzae pv. oryzae is a destructive disease of rice plants in Asia. It is an air-borne disease and several tadpole-shaped and filamentous phages of the pathogen were isolated from paddy soils and diseased rice leaves (e.g. CitationKamiunten and Wakimoto 1979; CitationKuo et al. 1967; CitationYoshii et al. 1953, CitationYoshimura et al. 1960). However, biocontrols of bacterial blight by those phages were unsuccessful, probably because changes in the phage population depend on that of the host population in nature (CitationEzuka and Kaku 2000). However, as a large population of the phages was detected from the irrigation water of severely infected fields prior to the disease appearance (CitationTagami 1959), a disease forecasting method was established and widely practiced in Japan from surveys of the phage population in paddy fields and channel water during the nursery and early paddy field stages (CitationJapanese Ministry of Agriculture and Forestry 1971; CitationTagami 1959), where one to two bacterial isolates sensitive to phages prevalent in the survey area were used to enumerate the phage population.
Soil and environmental factors, such as nutrient availability, pH, temperature, moisture content and soil structure, affect virus–host interactions, and it is often the case in soil environments that unexpected, contradictory viral effects are observed for host control, although such results tend not to be published (CitationAshelford et al. 2000). The inefficiency of viruses often experienced in host control results, in part, from an inactive and consequently insensitive state of host cells in undisturbed soils (CitationCasida and Liu 1974; CitationSmit et al. 1996). Under nutrient-limited conditions, phages are lytic when hosts are infected in their logarithmic phase, although they are pseudolysogenic for some time in the stationary phase (CitationMoebus 1996).
Virus adsorption to soils: A key factor in virus ecology in soils
Virus adsorption to soils
Adsorption of viruses to soils is a specific phenomenon profoundly influencing their infectivity to host organisms, and clay minerals play a central role in this phenomenon (CitationMarsh and Wellington 1994; CitationPantastico-Caldas et al. 1992; CitationStotzky 1985, Citation1986). The adsorption follows either the Freundlich or Langmuir isotherm (CitationBurge and Enkiri 1978; CitationMoore et al. 1981): the former under conditions of low surface coverage by viruses, and the latter under conditions of nearly saturated surface coverage (CitationMoore et al. 1981). The amount of viral adsorption was found to be linearly related to the square root of time by CitationBurge and Enkiri (1978). In addition, CitationChattopadhyay and Puls (2000) suggested that viruses adsorb to colloidal clays using van der Waals forces. Phages were observed using TEM to adsorb to kaolinite particles by their tails (CitationBystricky et al. 1975).
The degree of adsorption of viruses to soils is generally more than 90% for all viruses (CitationBitton et al. 1978; CitationGoyal and Gerba 1979; CitationMoore et al. 1981, Citation1982). A variety of soil, environmental and viral factors affect the adsorption of viruses to soils as shown in . These parameters interact in viral adsorption on soils because viruses are, in general, negatively charged above the isoelectric point, whereas soils have negatively and positively charged sites as well as hydrophobic sites. The pH influences the charge on both viruses and soils, and ionic strength and its constituents determine the binding force between them. In contrast, organic matter (both soil-associated and dissolved) weakens the electrostatic binding of viruses to soils. The adsorption of the T2 phage of E. coli and Type 1 poliovirus on clay particles (kaolinite, montmorillonite and illite) was first demonstrated by CitationCarlson et al. (1968) with an increase in viral adsorption associated with an increase in Na and Ca concentrations. The mechanism of viral adsorption to clays was via the clay–cation–virus bridge. Among cations, divalent cations were more effective as bridging agents than monovalent cations. And naturally occurring river clays (the Missouri River) showed very similar responses of viral adsorption to those of pure clay minerals.
Table 2 Soil, environmental and viral factors affecting the adsorption of viruses to soils
Type of clay minerals and type of viruses
Clay minerals have both positively and negatively charged sites, and those sites contribute to viral adsorption to clay minerals. CitationSchiffenbauer and Stotzky (1982) demonstrated that coliphages T1 and T7 had a greater affinity to montmorillonite than to kaolinite, and T7 showed a greater affinity to both clays than T1. T1 adsorbed primarily on positively charged sites of kaolinite and on both positively and negatively charged sites of montmorillonite. In contrast, T7 adsorbed more on negatively than on positively charged sites of both clays, which was reflected in an increase in its adsorption with an increase in the cation exchange capacity (CEC) of the clays. Reovirus type 3 had a similar adsorption tendency to T7 phage with more affinity to montmorillonite than to kaolinite, and a good correlation of adsorption to clays with CEC of the clays was observed because adsorption primarily occurred on negatively charged sites of the clays (CitationLipson and Stotzky 1983). A phage PBS1 of Bacillus subtilis behaved differently to reovirus Type 3 in its adsorption to clays, and there was no correlation between adsorption of PBS1 and CEC of montmorillonite and kaolinite (CitationVettori et al. 1999). Reovirus Type 3 and coliphage T1 did not share common adsorption sites on montmorillonite and kaolinite, which suggests the importance of using more than one type of virus, especially in combination, to predict virus behavior (e.g. adsorption, loss of infectivity) in soils and sediments containing clay minerals (CitationLipson and Stotzky 1985b). The clay minerals, kaolinite and montmorillonite, suspended in both distilled water and mineral medium (M-9), were better adsorbers for coliphage T1 or T7 than were bacteria, actinomycetes and yeasts. Except for the host bacteria, the microbial cells adsorbed few or no coliphages (CitationSchiffenbauer and Stotzky 1983).
CitationCarlson et al. (1968) first revealed the competition of egg and bovine albumin with viruses for adsorption sites on clay particles. Soils, especially soils rich in organic matter, are in general weak adsorbers compared with pure clay minerals. CitationLipson and Stotzky (1984b) examined the effects of chymotrypsin, ovalbumin and lysozyme on the adsorption of a reovirus to kaolinite and montmorillonite. Chymotrypsin and ovalbumin suppressed viral adsorption to both types of clay minerals, whereas lysozyme only suppressed viral adsorption to montmorillonite. The suppression mechanism of these proteins on viral adsorption to clay minerals was the competition of proteins with the reovirus for sites on the minerals. Among the minerals studied, montmorillonite, glauconite and bituminous shale were the least effective, whereas the most effective adsorbers were magnetite sand and hematite, both of which are predominantly oxides of iron (CitationMoore et al. 1981). Similar suppression effects of soil organic matter (mineral-associated and dissolved) on virus adsorption to soil were also observed for the somatic ssDNA φX174 phage and the unenveloped ssRNA MS-2 phage (CitationZhuang and Jin 2003a).
pH
CitationGoyal and Gerba (1979) conducted a comparative adsorption study of a number of different types and strains of human enteroviruses and bacteriophages to nine different soils, and found that soils with a pH of less than 5 were generally good adsorbers. CitationLoveland et al. (1996) studied the adsorption of an enteric phage PRD1 to quartz and ferric oxyhydroxide-coated quartz surfaces over a range of pH values and found irreversible adsorption of the virus to the surface below a critical pH (a pH approximately 2.5 to 3.5 pH units above the isoelectric pH point), but reversible adsorption above that critical pH.
Ionic strength and its constituents
Adsorption of viruses to low concentrations of clays was greater in estuarine water than in distilled water because the higher ionic strength of the estuarine water reduced the electrokinetic potential of both clay and virus particles (CitationLipson and Stotzky 1983). Thus, the addition of cations to distilled water enhanced viral adsorption, with divalent cations being more effective than monovalent cations. The sequence of the amount of adsorption to homoionic montmorillonite was Al > Ca > Mg > Na > K, whereas that to kaolinite was Na > Al > Ca > Mg > K (CitationLipson and Stotzky 1983). Soil adsorption was generally accelerated with the addition of Ca2+ and Mg2+ to the soil (CitationAzuaga et al. 1990; CitationGoyal and Gerba 1979; CitationMoore et al. 1982). Al3+ was the most effective cation in limiting poliovirus Type 1 penetrations through a soil column, probably owing to flocculation of the viruses, whereas organic compounds retarded the adsorption (CitationLance and Gerba 1984). Both natural humic material and sewage sludge organic matter increased the unsaturated flow of phage MS-2 in soil (CitationPowelson et al. 1991). As a counter anion, NO− 3, SO−2 4 and H2PO− 4 anions were more effective than Cl− in promoting the adsorption of poliovirus Type 1 (CitationLance and Gerba 1984). However, virus adsorption on Al-oxide coated sand was suppressed in the presence of HPO2− 3 compared with HCO− 3, and the effect of H2PO− 4 was more significant on phage φX174 than on phage MS-2. The presence of the divalent cations Ca2+ and Mg2+ suppressed viral adsorption on Al-oxide coated sand because the cations partially screened the negative charges on the phages, thereby decreasing the electrostatic attraction between the positively charged sand surface and the negatively charged phages (CitationZhuang and Jin 2003b). In addition, even small amounts of hydrophobic material in porous media can retard virus transport (CitationBales et al. 1993).
Viral adsorption to soil is highly strain dependent; less adsorption to soils was found by some echoviruses (Types 1, 2 and 29) and a simian rotavirus (SA1) (CitationGoyal and Gerba 1979), whereas insignificant effects of specific surface area and pH were observed on poliovirus adsorption to soils (CitationMoore et al. 1981). As coliphages MS2 and F2 adsorbed poorly to most soils, CitationBales et al. (1989) proposed to use them as an indicator for better estimating the maximum subsurface transport rate of colloidal contaminants through a porous formation. Although CitationBurge and Enkiri (1978) found a fair correlation between the specific surface area of five soils and the adsorption rate of phage φX174 of E. coli, adsorption of reovirus Type 3 was not related to the surface area of kaolinite and montmorillonite (CitationLipson and Stotzky 1983).
Efficient buffer systems for extracting viruses from soils, therefore, are absolutely important to establish virus ecology in soils, although various organic and inorganic buffers have been examined for greater extraction of viruses from soils and sediments. CitationWilliamson et al. (2003) suggested the importance of multiple extraction and enumeration approaches before undertaking ecological studies of phages in a particular soil.
Movement of viruses in soils (epidemiological concerns)
Epidemiological concerns in relation to virus ecology in soils are to trace the fate of pathogenic viruses discharged from wastewater treatment facilities and those present in aquifers and groundwater. The movement of viruses in soils is the opposite phenomenon of viral adsorption in soils, and is adversely related to viral adsorption to soils. Infiltration of viruses through soil columns, their soil surface run-off and their survival in soils were evaluated for various viruses, including coliphage, echoviruses, polioviruses, coxsackieviruses and rotavirus (e.g. CitationGoyal and Gerba 1979; CitationYates et al. 1985). Although soils are efficient adsorbers of viruses, viruses can migrate distantly in horizontal and vertical directions from several to dozens meters. Factors affecting the survival and migration of viruses in groundwater include soil texture, organic matter (humic acids), cations, adsorption property, pH, ionic strength, CEC, iron oxide level and permeability (CitationKeswick and Gerba 1980). In addition, virus type, microbial antagonism, rainfall, temperature, desiccation, flow rate and sunlight are also factors. However, rapid transport of the bacteriophage Salmonella typhimurium 28B was observed in a willow-cropped lysimeter experiment with clay soil compared with sandy soil because of bypass transport of the phage through macropore formation in the clay soil (CitationCarlander et al. 2000). Viruses penetrated deeper into the ground and moved longer distances than did coliform bacteria (CitationKeswick and Gerba 1980). Faster transport of F pilus-specific RNA coliphages than fecal coliforms was also observed in irrigated effluent through an alluvial aquifer near Christchurch, New Zealand (CitationSinton et al. 1997). CitationJin et al. (1997) evaluated virus transport through sand columns (9.2-cm diameter and 10.5 or 20-cm long) under saturated flow conditions using two bacteriophages, MS2 and φX174, and found that MS-2 was not sorbed to the Ottawa sand, whereas a significant amount of the applied φX174 was retained without inactivation in the columns, probably because of the difference in the isoelectric points of their surfaces. Similarly, CitationSchijven et al. (2002) observed 3–10-fold higher transport rates of phage φX174 than of phages MS2 and PRD1, and this was attributed to the lower electrostatic repulsion of the less negatively charged φX174. Among five different spherical phages (MS2, PRD1, QB, φX174 and PM2) with different isoelectric points (pI 3.9, 4.2, 5.3, 6.6 and 7.3, respectively), the smaller phages (MS2, QB and φX174) with similar diameters exhibited maximum transport that correlated with their isoelectric points. In addition, when virus particles are more than 60 nm in diameter, their viral dimensions are believed to become an overriding factor (CitationDowd et al. 1998).
In a survey of the rapid infiltration of wastewater containing a tracer virus, coliphage F2, indigenous enteroviruses and enteric indicator bacteria through unconsolidated silty sand and gravel, coliphage F2 concentration in a 18.3-m deep well began to increase within 48 h after application to the soil and stabilized after 72 h at a level of approximately 47% of the average applied concentration, irrespective of its concentration in the upper soil layers. In addition, indigenous enteroviruses and tracer F2 were sporadically detected in the groundwater at horizontal distances of 183 m from the application site (CitationSchaub and Sorber 1977).
CitationPowelson et al. (1993) assessed soil-aquifer treatment of sewage effluent for removing bacteriophages MS2 (25 nm in diameter) and PRD1 (62 nm). PRD1 was removed more rapidly during percolation than MS2. Effluent type did not significantly affect viral removal rate for MS2, but the PRD1 removal rate was nearly threefold greater for secondary effluent than for tertiary effluent with virus removals at the 4.3-m depth ranging from 37 to 99.7%. The phage MS2 was adsorbed to clay minerals in the order of fine Na-montmorillonite > coarse Na-montmorillonite > Ca-montmorillonite and kaolinite, and the amounts of clay minerals that were transported through the columns followed the same order (CitationJin et al. 2000), which demonstrated that colloid-mediated transport was a dominant mechanism for virus transport through porous media. CitationKapuscinski and Mitchell (1980) emphasized the importance of viral association with colloidal and particulate materials in their transport in sewage coastal water management.
Environmental factors affecting the inactivation and survival of viruses in soils
A variety of soil and environmental factors affect viral inactivation and infectivity to host organisms, among which are temperature, pH, clay type and its content, organic matter content (nutrient status), heavy metals and acid pollutants, aerobicity (anaerobiosis), ionic strength (salinity), and moisture content.
Temperature
Bacteria are grouped into psychrophilic, mesophilic and thermophilic depending on their optimal growth/lethal temperature. Inactivation and survival of phages in soils are also temperature dependent, and the temperature dependency of the phages is in some cases not the same as that of their hosts.
Temperature is a major controlling factor for viral survival in soil. CitationYeager and O’Brien (1979a) studied the effect of temperature (4, 22 and 37°C) on the inactivation of poliovirus Type 1 in saturated soil and found that p.f.u. were recovered for up to 12 days at 37°C, whereas at 4°C p.f.u. were recovered from the soil for up to 180 days. CitationHurst et al. (1980) studied several viruses, coxsackieviruses (A9 and B3), echovirus Type 1, poliovirus Type 1, rotavirus SA11 and bacteriophages T2 and MS2, to elucidate the effects of temperature, soil moisture content, presence of aerobic microorganisms, degree of virus adsorption to soil, concentrations of resin-extractable P and exchangeable Al and soil pH on their survival, and temperature and virus adsorption to soil appeared to be the most important factors affecting their survival. CitationYates et al. (1985) revealed from multiple regression analysis that temperature was the only variable among numerous chemical and physical factors that correlated significantly with the decay rates of poliovirus, echovirus and coliphage in groundwater. In general, lower temperatures resulted in longer periods of survival, longer latent periods and reduced burst size of phages (CitationLeonardopoulos et al. 1996a,Citationb; CitationStraub et al. 1992; CitationZachary 1978). It is probably because higher microbial and enzymatic activities, such as proteinase under higher soil temperature conditions, result in faster inactivation and decomposition of viruses. For example, phages of Geobacillus stearothermophilus (previously called Bacillus stearothermophilus) are ubiquitous in soil, and they were greatest in abundance at 45°C when they were pre-incubated in soil with the host, whereas their proliferation was usually much less at 55°C (CitationReanney and Marsh 1973). Retention of plaque forming ability at 50°C, but not at 70°C, was also found for 24 phages of thermophilic Bacillus strains that were isolated from compost, soil, silage and rotting straw (CitationSharp et al. 1986). Similarly, a phage infecting Bacillus circulans grew most at 45°C compared with 30, 37 or 55°C in a soil incubation experiment (CitationTan and Reanney 1976).
The inactivation of naturally occurring bacterial indicators of sludge and sewage contaminants (E. coli, fecal streptococci and spores of sulfite-reducing clostridia) and phages (somatic coliphages, RNA F-specific phages and phages infecting Bacteroides fragilis strain RYC 2056) was studied for the effect of thermal treatment on dewatered sludge at 80°C and raw sewage at 60°C, and those phages were shown to be significantly more resistant to thermal inactivation than bacterial indicators (CitationMoce-Llivina et al. 2003).
pH
Soil pH affects not only host growth, but also viral survival. CitationSykes et al. (1981) attempted to isolate the bacteriophages of five neutrophilic and three acidophilic streptomycetes from various soils with different pH levels, and no phage was detected in soils with a pH below 6.0, despite the presence of acidophilic streptomycetes in those soils. However, some phage isolates from neutral soils could lyse acidophilic streptomycetes at pH 5.5. CitationSykes and Williams (1978) found a loss in the infective potential of an actinophage to its host at pH 6.1 and 4.9 under the conditions of kaolin-adsorbed and free states, respectively. This was attributed to the accumulation of protons around the kaolin surfaces, 10-fold larger (1 pH unit), in comparison with the proton concentration in bulk soil water. Acidity might have multiple effects on adsorption, penetration and the length of the latent period in phage–host interactions (CitationSykes et al. 1981).
Soil pH also influences viral survival indirectly. The adsorption of viruses to a solid surface is a common occurrence that results in a longer duration of survival. Viruses are hydrophilic colloidal materials, and their adsorption depends largely on the electrostatic properties of the soil surface to which the pH greatly contributes (CitationKapuscinski and Mitchell 1980).
Clay type and its content
Poliovirus Type 1 and bacteriophages MS2 and PRD-1 added with sewage sludge to desert soils in Arizona, USA, survived for a longer period in a clay loam soil than in a sandy soil (CitationStraub et al. 1992). Similar findings of better persistence and resultant suppression of nodule formation by phage Zag-1 of R. leguminosarum bv. trifolii in clay soil than in sandy soil were also reported from cowpea fields in Egypt (CitationHussein et al. 1994). The rate of inactivation of phage φ11M15 of Staphylococcus aureus RN 450 in natural, autoclaved and filtrated lake water was greatly reduced in the presence of attapulgite and vermiculite and, to a lesser extent, of montmorillonite and kaolinite (CitationBabich and Stotzky 1980). Loss of reovirus infectivity in synthetic estuarine and distilled water containing montmorillonite or kaolinite was less than that in water without clays (CitationLipson and Stotzky 1985a). It has also been reported that viral association with colloidal and particulate materials prolonged survival in coastal waters (CitationKapuscinski and Mitchell 1980). Although adsorption of poliovirus Type 1 to sandy loam soil protected the virus from inactivation, adsorption to sand had little effect on the rate of poliovirus inactivation (CitationYeager and O’Brien 1979a). CitationLipson and Stotzky (1986) studied the mechanisms whereby particulates affect the specific infectivity of viruses. They used reovirus Type 3, kaolinite and L-929 mouse fibroblasts as the model system. Settling of the clay–virus complex on the L-929 cell monolayer increased the number of infectious virions reaching the cell surface. And the affinity of the reovirus to the cells was greater than to the clay, which was reflected in the difference in the energies of adsorption and binding between the virus and the cells or kaolinite. Thus, kaolinite enhanced the transfer of viral particles, in conjunction with diffusion and Brownian movement, to receptors of the reovirus on the host cell surface, primarily as a result of the more rapid settling of the kaolinite–virus complex on the cell surface (CitationLipson and Stotzky 1987). Clay minerals can protect phages against inactivation and loss of transducing ability by ultraviolet (UV) radiation, partly because they shield phages from UV light. CitationVettori et al. (2000) studied the effects of UV radiation on the survival of and transduction by phage PBS1 of B. subtilis under free or adsorbed states on montmorillonite and kaolinite. The titer and transduction frequency of clay-associated phages remained essentially constant between 1 and 10 min of UV radiation, whereas the titer of free phage decreased by approximately one order of magnitude after 5 min of UV radiation.
In addition, almost complete protection of E. coli from phage attack was provided by fine montmorillonite particles, although particles with an effective spherical diameter greater than 0.6 µm did not protect the bacterium from phage lysis (CitationRoper and Marshall 1978).
Thus, clay minerals protect viruses against both biological and abiological inactivation, thereby enabling them to persist in soils for longer periods of time in the absence of their hosts (CitationStotzky 1986).
Organic matter content (nutrient status)
CitationDelisle and Levin (1972) tested plaque formation on a strain of Pseudomonas putrefaciens using psychrophilic phages that had been isolated from seawater and haddock meat and found that plaque formation was successful when the strain was grown on trypticase soy broth, but was unsuccessful when grown on trypticase soy broth with oxalate. This finding indicates that differences in substrate composition, for example, the kind of soils, influence the infectivity of phages to host bacteria, the susceptibility of the host bacteria, or a combination of the two in soils. One of the reasons of immunity of host cells to phage infection probably results from changes in the surface property of the host cells. For example, B. subtilis W23 was shown to lack wall teichoic acid when grown under conditions of phosphate limitation and did not adsorb phage SP50, the first step in phage infection (CitationArchibald and Coapes 1976). CitationLechevalier and Lechevalier (1970) analyzed some 600 strains of aerobic actinomycetes for their whole-cell sugar patterns and cell-wall compositions and classified them into 10 taxonomically different groups (Groups I to X). Polyvalent phages that were infectious to Streptomyces spp. infected hosts belonging to Group I genera, whereas all strains within Groups II, III, IV and VI (31 strains from 11 genera) were resistant to phages infectious to Group I genera (CitationWellington and Williams 1981).
Soil is an environment that generally provides poor substrates for microbial communities, and indigenous host cells may be in an insensitive state to viral infection in soil. Phages of Arthrobacter globiformis were rarely detected in soil, but they were continuously produced in nutrient-amended soil and the addition of host cells was not required for phage detection (CitationCasida and Liu 1974). Indigenous phages seem to be present in an inactivated state in soils, and the immunity of host cells to phages as a result of their physiological state may be phage specific or host specific. For example, CitationSchrader et al. (1997) examined five phages specific for P. aerginosa and E. coli for their ability to multiply in host cells in stationary phase and found that the P. aeruginosa phage BLB was incapable of multiplication in a stationary phase host, but that significant multiplication of phage ACQ occurred in P. aeruginosa RM754 host cells that had been starved for periods of 24 h, several weeks or 5 years. Thus, starvation did not reduce bacterial hosts refuging from phage infection in the phage–host system of the latter.
Heavy metals and acid pollutants
A 10 mmol L−1 concentration of Zn2+ did not significantly affect the survival of coliphages T1, T7, P1 and φ80. However, the toxicity of zinc to those coliphages increased with the addition of high concentrations of NaCl (1 and 5 mol L−1) because of greater toxicities of the anionic mixture of ZnCl− 3/ZnCl2− 4 that was formed by NaCl addition (CitationBabich and Stotzky 1978b). In contrast, mixtures of anionic HgCl− 3/HgCl2− 4 complexes were less toxic to phages (φ11M15 of S. aureus and P1 of E. coli) than were equivalent concentrations of cationic Hg2+. The toxicity of 1 ppm Hg to φ11M15 was less in seawater than in lake water, indicating that the lower toxicity of Hg in seawater was a result of the formation of HgCl− 3/HgCl2− 4 complexes in the seawater (CitationBabich and Stotzky 1979). Organic materials seem to alleviate the toxicity of heavy metals to viruses by forming organo-metal complexes. The toxicity of nickel to coliphage T1 was more serious at respective concentrations (5, 50 and 100 × 10−3 g Ni L−1) in fresh water > estuarine water > sea water in this order. However, 100 × 10−3 g Ni L−1 was not toxic to T1 in nutrient broth, probably because of the protective effect of the organic compounds in the broth (CitationBabich et al. 1983).
The effect of sulfur dioxide (SO2) in acid rain on coliphage T1 was evaluated in a test tube by changing pH and HSO− 3 and SO2− 3 concentrations. HSO− 3 was always more inhibitory than SO2− 3, and the inhibitory effects of HSO− 3 and SO2− 3 increased with a decrease in pH (CitationBabich and Stotzky 1978a). No study has been conducted on the mechanism of toxicity of heavy metals and acid pollutants to viruses. As viruses are imagined to be inert in a free state, the mechanism is an interesting subject for future study.
Aerobicity (anaerobiosis)
Replication under anaerobic conditions resulted in longer latent periods of phages nt-1 and nt-6 of Vibrio natriegens. However, the effect of anaerobiosis on burst size was different between these phages: phage nt-6 had a reduced burst size, whereas nt-1 had an increased burst size (CitationZachary 1978). Therefore, the composition of phages in lowland areas, including rice fields, may vary according to reclamation and water management.
Ionic strength (salinity)
Two phages (nt-1 and nt-6) of the marine bacterium Beneckea natriegens (Vibrio natriegens) were examined to determine the effect of ionic strength on their stability and replication. They were more resistant to low NaCl levels than their host bacterium, which appeared to confine their growth to marine waters because of the optimal higher salinity of B. natriegens survival (CitationZachary 1976). CitationMoebus (1987) summarized the effects of ionic strength of Ca2+, Mg2+ and Na+ on adsorption on host cells, burst size, latent period and the survival of phages in seawater. Virulence of the phage PK-101 of Pseudomonas solanacearum K-101 increased with the addition of CaCl2–MgCl2 (0.15 mol L−1) (CitationToyoda et al. 1991). Adsorption of viruses to clay particles is greater under higher ionic strengths because higher ionic strengths reduce the electrokinetic potential of both clay and virus particles. And viral association with colloidal and particulate materials prolongs the survival and infectivity of viruses. Thus, ionic strength is an important environmental factor coupling with clay particles for the survival of viruses.
Moisture content
Soil drying results in profound effects on viral inactivation. There appears to be a critical point of soil moisture below which virucidal effects of evaporation are amplified. For example, poliovirus Type 1 was inactivated at essentially the same rate between 18 and 2.9% moisture content, but the rates of viral inactivation increased markedly between 1.2 and 0.6% moisture for a sandy loam soil (CitationYeager and O’Brien 1979a). Viral inactivation by soil drying resulted from the virucidal effects of the evaporative process per se as well as from the effects of low soil moisture levels, and evaporation was estimated to be the primary factor responsible for viral inactivation in drying soil for poliovirus Type 1 (CitationYeager and O’Brien 1979a). The irreversible binding of viruses that may occur as a result of drying in soil environments is another factor affecting viral inactivation. CitationYeager and O’Brien (1979a,Citationb) used radioactively RNA-labeled and capsid-labeled poliovirus Type 1 and found that both the RNA and capsid portions of the virions were recovered from moist soils, but that only the RNA label was recovered efficiently from the dried soils. Inactivation of poliovirus also occurs in moist soils because of the damage of RNA before its release from the capsid. In contrast, the dissociation of virions into intact RNA and isoelectrically altered capsids was noted in dried soils (CitationYeager and O’Brien 1979b). Inactivation of poliovirus Type 1 strain CHAT by cleavage of vrial proteins followed by nicking of encapsulated RNA was also observed in digested sludge (CitationWard and Ashley 1976).
To assess sanitary risks by pathogenic, enteric viruses present in the sludge, laboratory studies were conducted to measure the inactivation rate of poliovirus Type 1 and bacteriophages MS2 of E. coli and PRD-1 of Salmonella typhimurium in two sludge-amended desert agricultural soils (sandy loam and clay loam soils) (CitationStraub et al. 1992). As the temperature increased from 15 to 40°C under constant moisture (30% content, approximately –0.05 × 105 Pa for both soils) and temperature conditions, the inactivation rate increased significantly with the increase in temperature for poliovirus and MS2, whereas a significant increase in the rate was observed only at 40°C for PRD-1. For all three viruses, drying to less than 5% soil moisture resulted in a more rapid loss of infectivity than at a constant moisture. Evaporation completely inactivated all three viruses within 7 days at 15°C, within 3 days at 27°C and within 2 days at 40°C, regardless of soil type. CitationWilliamson et al. (2005) measured viral-like particles (VLPs) in six Delaware soils, USA, using epifluorescence microscopy and found a significantly positive correlation between VLP abundance and soil water content.
In addition, as CitationWilliamson et al. (2005) found a significant difference in VLP abundance between agricultural soils and forest soils, land-use form and land management practices might be important underlying factors controlling viral abundance in soil environments.
Host specificity of phages in soil
Host specificity of rhizobiphages may match the infection range of Rhizobium spp. Eleven phages of Rhizobium loti, R. leguminosarum bv. trifolii, Rhizobium galegae and Rhizobium meliloti were isolated from fields where legumes grew and they were used for phagotyping of nodule bacteria with unknown taxonomic position from milk vetch (Astragalus), liquorice (Glycirrhiza), rest-harrow (Ononis), French honeysuckle (Hedysarum) and crown vetch (Coronilla) (CitationNovikova and Limeschenko 1992). Phages of R. meliloti, R. leguminosarum and R. galegae lysed only bacterial strains of homologous species. However, in a follow-up study, CitationNovikova et al. (1993) did not find a relationship between the infectivity of phages to various Rhizobium spp. isolated from R. loti, R. meliloti, R. leguminosarum and R. galegae root nodules and the nodule-forming capacity of those Rhizobium strains among legume plants. CitationHashem et al. (1996) proposed a rapid method for identifying and characterizing Sinorhizobium fredii strains from their susceptibility to various phages. Host specificity of phages to X. oryzae pv. oryzae (pathogen of bacterial blight of rice plant) is different according to the strains of phages and isolates of the bacterium. And the bacterium was classified according to the lysotype to various phage strains (CitationWakimoto 1967). Although the reason was not clear, CitationKankila and Lindström (1994) found an inverse relationship between the host range of phages of R. leguminosarum bv. trifolii and the level of susceptibility of phage DNA to restriction enzymes. Host range of phages is, in general, within the genus level (CitationLong et al. 1993).
Horizontal gene transfer by viruses
Transduction is a mechanism for gene transfer mediated by viruses among bacteria. There are two mechanisms of transduction: generalized transduction and specialized transduction. In generalized transduction, lytic phages package DNA from the infected bacterium, whose chromosome has been cleaved into small segments by nucleases, into the phage capsid. As any single or group of bacterial genes may be incorporated in this transduction, it is designated as generalized transduction. The transducing phages carrying bacterial DNA inject their DNA, including the bacterial DNA fragment, into new bacterial host cells. Specialized transduction is mediated by a lysogenic phage, where the phage genome is integrated into the bacterial chromosome, usually at a specific site, and replicates faithfully with the bacterial chromosome. Upon induction of the lytic cycle of phage replication, imprecise excision of the phage genome results in the packaging of adjoining bacterial genes on the chromosome into the virion with the prophage DNA. Thus, as this transduction transfers only genes that are close to the site of phage integration, it is termed specialized transduction (CitationStotzky 1989; CitationStotzky et al. 1990; CitationYin and Stotzky 1997).
Generalized transduction of E. coli K-12 by P1 coliphage was demonstrated using auxotrophic recipient cells (thr-leu-thi-rpsL) and transducing phage lysates from transposon 10 (Tn10) in sandy and silty clay loam soils. The frequency of transduction in soil was approximately 10−6 (CitationGermida and Khachatourians 1988). The specialized transduction of bacterial resistance genes for chloramphenicol and mercury into E. coli was demonstrated when the transducing coliphage P1 was added to non-sterile soil as either lysates or E. coli lysogens (CitationZeph et al. 1988; CitationZeph and Stotzky 1989). CitationRipp and Miller (1995) examined the effects of mackaloid clay and particulate matter in freshwater from Lake Carl Blackwell water, Oklahoma, USA, on the frequency of transduction among Psuedomonas aeruginosa mediated by temperate phage F116, and observed an increased frequency in generalized transduction by 100-fold, because aggregations of phages and bacterial cells are stimulated by the presence of these suspended particulates.
Lysogeny is common and occurs frequently in bacterium–phage relationships, and up to 40% of isolates of P. aeruginosa from natural ecosystems were found to contain DNA sequences homologous to phage genomes, evaluated using radiolabeled bacteriophage DNA probes (CitationOgunseitan et al. 1992). Lysogenization is a mechanism of gene transfer in soil among closely related bacterial communities. CitationHerron and Wellington (1994) observed that the temperate phage KC301 incorporated into Streptomyces lividans TK23 (S. lividans TK23[KC301]) was recovered from S. lividans TK24 as S. lividans TK24(KC301) in soil. Transfer of KC301 from TK24(KC301) to a Streptomyces violaceolatus strain was also recognized in natural soil amended with nutrients (CitationCresswell et al. 1992). In general, the lysogenized organisms are considered to be less adapted to the soil environment in comparison with the original prophage-free organisms because of DNA reproduction of inserted prophage sites (CitationHerron and Wellington 1994; CitationLenski and Levin 1985), although CitationAshelford et al. (2000) did not observe any difference in the survival of Serratia liquefaciens CP6RS and its lysogen (CP6RS-ly-Φ1), which had been inoculated to sugarbeet seeds during a 194-day growth period.
Enumeration of viruses in soils
For accurate evaluation of viral abundance, reliable methods of extraction of viruses from soils, separation of viruses from bacterial communities, storage of extracted viruses, and their enumeration are required.
Extraction of viruses from soils
As the majority of viruses are adsorbed to the solid phases of soils, solutions of various composition have been devised for better extraction of viruses from soils and sediments; for example, nutrient broth (CitationGermida and Casida 1981; CitationLanning and Williams 1982; CitationNakayama et al. 2007a), nutrient broth containing egg albumen (CitationLanning and Williams 1982; CitationWilliams et al. 1987), soil extract (CitationGermida and Casida 1981), beef extract (CitationHu 1998; CitationMonpoeho et al. 2001; CitationPaul et al. 1993; CitationWilliamson et al. 2003), beef extract with NaCl (CitationMonpoeho et al. 2001), beef extract containing borate (CitationMonpoeho et al. 2001), cation-complete medium (CitationGermida and Casida 1981), glycine buffer (CitationHu 1998; CitationMonpoeho et al. 2001; CitationWilliamson et al. 2003), sodium pyrophosphate (CitationDanovaro et al. 2001; CitationWilliamson et al. 2003, Citation2005), potassium citrate (CitationWilliamson et al. 2003, Citation2005), AlCl3 (pH 3.5) (CitationMonpoeho et al. 2001), and mineral buffer (CitationMonpoeho et al. 2001). In general, beef extract (6–10%) has most widely been used for the extraction of viruses from soils.
Separation of viruses from bacterial communities
Filtration through membranes of 0.22–0.45 µm pore size is most commonly used for the separation of viruses from bacterial communities. The procedure, in general, results in a significant loss of viruses in extracts (CitationLanning and Williams 1982; CitationWilliams et al. 1987), with a two-third reduction in some cases (CitationPaul et al. 1991). Moreover, the selection of the filter material is important because of viral adsorption on filter membranes (CitationTartera et al. 1992). It should be noted that separation of viruses from bacterial communities using such filtration techniques results in disregarding viruses in host cells.
Storage of extracted viruses
A 2–2.5% final concentration of glutaraldehyde is generally used for the preservation of water samples and viral extracts (CitationCochlan et al. 1993; CitationCochran and Paul 1998; CitationDanovaro et al. 2001) in addition with 2% final concentration of formaldehyde (CitationDanovaro et al. 2001; CitationNoble and Fuhrman 1998). However, as formaldehyde and glutaraldehyde are known to induce significant reductions in virus counts after 24 h of storage of marine sediments (CitationDanovaro et al. 2001), immediate measurement of viral abundance in water samples and extracts is recommended. Deep freezing at −20°C or colder is recommended for viral storage.
Enumeration
As the infectivity of viruses is specific with respect to hosts, enumeration of viral abundance by plaque formation is used only for detecting and isolating specific viruses. Because of the very strict host specificity of viruses, precondensation of samples and/or enrichment culture are generally necessary (CitationBishop et al. 1977). The double agar layer method developed by CitationAdams (1959) is the standard method for plaque counting. Enumeration of viral abundance by TEM has been commonly used since the first application of TEM to viral enumeration in seawater by CitationTorrella and Morita (1979), who counted phage particles collected on a 0.2-µm Nuclephore filter. They observed different morphologies of phages as well as phage–bacterium interactions. Specimen preparation for the quantitative collection of viral and bacterial communities in water samples and extracts on microscope grids was developed by CitationNomizu and Mizuike (1986). Although TEM is the established method for viral enumeration in water samples and viral extracts (CitationBratbak and Heldal 1993) and has been commonly used in the past and at present, epifluorescence microscopy has been introduced for viral enumeration in the environment because of the reasonable price of equipment, easy sample preparation and a similar degree of accuracy to the TEM method (CitationSuttle 1993) since the method was applied to marine waters by CitationHara et al. (1991). Yo-Pro (CitationBird et al. 2001; CitationHennes and Suttle 1995), YoYo-1 (CitationHennes et al. 1995), PoPo-1 (CitationHennes et al. 1995), DAPI (CitationHara et al. 1991, Citation1996; CitationSuttle 1993) and SYBR Green I (CitationNoble and Fuhrman 1998) have been used as fluorescent dyes for the estimation of viral abundance in marine and sediment samples. SYBR Green I enabled the rapid and accurate determination of viral abundance collected on 0.02-µm pore-size filters after staining for 15 min (CitationNoble and Fuhrman 1998). Viral counts by SYBR Green I were, on average, 1.28-fold higher than those made by TEM (CitationNoble and Fuhrman 1998). These researchers attributed the underestimate of viral abundance by TEM to a loss when uranyl acetate is wicked away from the grids and viruses may be obscured by other larger, darkly stained particles on the grids.
ABUNDANCE OF VIRUSES AND THEIR ROLES IN BIOGEOCHEMICAL NUTRIENT CYCLES IN AQUATIC ENVIRONMENTS
The importance of viruses in biogeochemical nutrient cycles and as genetic reservoirs in marine and freshwater environments results from their large abundance and great diversity. However, comparable information on viral abundance in soil environments has only been collected by CitationWilliamson et al. (2005) and CitationNakayama et al. (2007a). Therefore, this section introduces viral abundances in various marine and freshwater environments. Then, the significant role of viruses in biogeochemical nutrient cycles in aquatic environments is summarized. The importance of viruses in this matter results from the large abundance of viruses, the short generation time of host organisms, and the significant proportion of virus-mediated mortality in aquatic environments.
Table 3 Studies on temporal and spatial variations of viral abundance
Abundance
Viral abundance increases with the productivity of the system, and it is generally lowest in the deep sea (104–105 VLP mL−1), intermediate in offshore surface waters (105–106 VLP mL−1) and highest in coastal environments (106–107 VLP mL−1) (CitationWeinbauer 2004; CitationWommack and Colwell 2000). Viral abundance in estuaries or productive lakes can be as high as 108 VLP mL−1, with the highest viral number of 9.6 × 108 VLP mL−1 recorded in a cyanobacterial mat (CitationHennes and Suttle 1995). Viral abundance tends to be higher in freshwater than in marine systems (CitationMaranger and Bird 1995).
Numerous studies have examined temporal and spatial variations in total viral abundance as shown in . Sunlight (CitationBoehme et al. 1993; CitationGarza and Suttle 1998; CitationHedal and Bratbak 1991; CitationNoble and Fuhrman 1997; CitationSuttle and Chen 1992; CitationWilhelm et al. 1998; CitationWommack et al. 1996) and annual phytoplankton blooms (CitationBratbak et al. 1990; CitationHennes and Simon 1995; CitationMathias et al. 1995) are characteristic factors for viral abundance in aquatic environments. Viral abundance is larger where suspended particles (CitationHewson et al. 2001) and substrates (CitationPaul et al. 1991; CitationWeinbauer et al. 1993) are larger. Thus, factors controlling viral inactivation and abundance are different from those in the soil environment, where temperature, pH, clay particles, ionic strength (salinity) and moisture content are important factors. In marine environments, temperature and salinity are stable and seem not to affect viral communities directly, but they appear to be correlated with microbial production (e.g. phytoplankton blooms) and substrate concentration, such as the decrease in viral abundance along the transect from freshwater, estuarine to open ocean. The pH is generally stable in aquatic environments. Moreover, pelagic environments are generally free from clay particles in the strict sense.
In general, viral-to-bacterial ratios (VBRs) are between 3 and 25, and are higher in more nutrient-rich, productive environments, which is one of the reasons for viruses being the greatest genomic reservoirs (CitationFuhrman 1999; CitationWommack and Colwell 2000). Both positive and negative relationships between VBR and bacterial abundance have been observed in oligotrophic and eutrophic seawater. Although positive relationships were noted in both oligotrophic waters in the North Pacific (CitationHara et al. 1991) and eutrophic waters in the northern Adriatic Sea (CitationWeinbauer et al. 1995), the most commonly observed relationships are inverse between them in various seawaters (CitationJiang and Paul 1994; CitationMaranger and Bird 1995; CitationMaranger et al. 1994; CitationWommack et al. 1992). In the floodwaters of a Japanese paddy field, the relationship was also significantly inverse (CitationNakayama et al. 2007a).
Studies on viral abundance in sediments are surprisingly few in comparison with the voluminous studies in water columns in aquatic environments. Viral and bacterial abundances in sediments are of the order of 107–109 VLP mL−1 and 106–108 cells mL−1 of sediment, respectively, with VBRs fluctuating from 0.1 to 100 (CitationDanovaro and Serresi 2000; CitationDanovaro et al. 2001, Citation2002; CitationDrake et al. 1998; CitationHewson and Fuhrman 2003; CitationHewson et al. 2001; CitationMaranger and Bird 1996; CitationMiddelboe et al. 2003; CitationPaul et al. 1993; CitationSteward et al. 1996). As the measurements were conducted with either pore water or 10 mmol L−1 pyrophosphate buffer in these studies, wide variations in VBRs seem to be derived from the specificities of the study locations.
The VLP and bacterial abundances in the floodwater of a Japanese paddy field ranged from 5.6 × 106 to 1.2 × 109 VLP mL−1 and from 9.2 × 105 to 4.3 × 108 cells mL−1 during the period of rice cultivation from June to September, with mean abundances of 1.5 × 108 VLP mL−1 and 5.1 × 107 cells mL−1, respectively, indicating that the floodwater was rich in viral and bacterial abundances among various aquatic environments, and abundances increased with an increase in the turbidity of the floodwater (CitationNakayama et al. 2007a). Thus, the magnitude of seasonal variation was more than 50-fold for VLP abundance and 100-fold for bacterial abundance.
CitationWilliamson et al. (2005) extracted 109 VLP g−1 of soil with potassium citrate buffer from six Delaware soils, USA, under different vegetation (two agricultural soils and four forest soils). The abundance of VLPs was highest in the wetland forest soils (2.9–4.2 × 109 VLP g−1), followed by upland forest soils (1.3–1.5 × 109 VLP g−1) and agricultural soils (0.87–1.1 × 109 VLP g−1). The VBRs were extremely high in the agricultural soils (approximately 3000), whereas the VBRs in the forested soils were approximately 10. The extremely high VBRs in the agricultural soils were attributed to less efficient extraction of bacteria from agricultural soils. This was the greatest record of VLP abundance in soil ever published. Their results suggested seemingly conflicting strategies used by viruses to exist in soils: the large VBRs indicate that the virulent phages are dominant in those soils (the environment for r-strategists) even if the VBRs were approximately 10 in those soils. In contrast, the viral abundance per se indicates that lysogeny is a common lifestyle of viruses in soils (the environment for K-strategists) because most soils commonly contain bacteria in a range from 108 to 1011 cells g−1. More information on viral abundance in soils with more reliable extraction methods is a prerequisite for evaluating the roles of viruses in biogeochemical nutrient cycles and genomic diversity in soils.
Microbial loop in biogeochemical nutrient cycles in aquatic environments
Many studies indicate the ecological importance of viruses in primary production and microbial food webs in marine environments (e.g. CitationDucklow et al. 1986; CitationWilhelm and Suttle 1999). At present, it is generally estimated that viruses are responsible for approximately 10–50% of the total bacterial mortality in marine surface waters, and 50–100% in environments that are hostile to protists, such as low-oxygen lake waters. The viral effect is probably larger in eutrophic waters than in oligotrophic waters (CitationFuhrman 1999). shows the pelagic food chain model and virus-mediated carbon flow constructed by CitationWeinbauer (2004), where 6–26% of the carbon fixed by primary producers is estimated to enter into the dissolved organic carbon (DOC) pool via virus-induced lysis at different trophic levels. The situation is similar in freshwater environments, and CitationFischer and Velimirov (2002) estimated that the viral control of bacterial production ranges from 56 to 63%, with occasionally up to 1.6% h−1 of the bacterial standing stock being removed from the water column, while grazing of heterotrophic nanoflagellates accounted, on average, for only 5% of the bacterial mortality in a eutrophic oxbow lake of the Danube River.
Figure 1 Pelagic food chain model and virus-mediated carbon flow (Weinbauer 2004). The dotted lines show virus-mediated pathways. All values are based on the flux of carbon fixed by primary producers (100%).
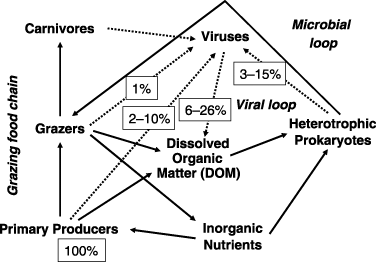
Much of the DOC derived from viral lysis of bacteria is not transferred to higher trophic levels, but is recycled through the microbial community, which is termed the “microbial (bacterium–phage–DOC) loop” (CitationAzam et al. 1983; CitationFuhrman 1992). The total production of heterotrophs can greatly exceed primary production because of the recycling mediated by phages in the marine environment (CitationFuhrman 1992). Thus, repeated cycling of organic materials in the microbial loop causes the bacteria to be efficient sinks for C and in the regeneration of N and P in environments where viruses are important agents of microbial mortality (CitationWeinbauer 2004; CitationWommack and Colwell 2000).
The importance of viruses in the microbial loop in biogeochemical nutrient cycles in marine environments is a result of the large amount of microbial biomass and their fast mortality rate by viral lysis. According to the summaries by CitationWeinbauer (2004), as phage-infected bacterial cells ranged from 1.4 to 26% in oceanic waters and from not detectable to 43% in coastal/shelf waters in bacterial communities, phage-mediated mortality of bacteria was 25% in oceanic waters and 58% in coastal waters. As viral turnover time ranged from 0.1 to 25 days, roughly 10–20% of bacterial members are estimated to lyse through phage infection on a daily basis in marine environments (CitationSuttle 1994; CitationWhitman et al. 1998; CitationWommack and Colwell 2000).
The significant role of viruses in biogeochemical nutrient cycles is only as a driver of the microbial loop. The total biovolume of VLPs ranged from 0.027 to 2.9 cm3 m−3 with a median volume of 0.31 cm3 m−3 in the floodwater of a Japanese paddy field, where the virus-to-bacterium biovolume ratios were estimated to be less than 0.002 because the ratio (0.002) referred to the case of bacterial communities being coccal with a diameter of 0.45 µm (the pore size diameter of the Nuclepore filter used for bacterial enumeration by epifluorescence microscopy; CitationNakayama et al. 2007b). As viruses are functionally active only inside host cells, their life cycles synchronize to those of the host organisms. In addition, not all the host cells are burst by viruses synchronously. Therefore, the contribution of biovolume of virus communities per se is negligible in nutrient cycles in the floodwater in comparison with that of bacterial communities, which is in contrast to the important contribution of microbial biomass in nutrient supply to crops in soils (CitationJenkinson and Ladd 1981).
Lytic viruses may contribute more to the activation of the microbial loop in the biogeochemical nutrient cycles than lysogenic viruses. And as a greater incidence of lysogenic bacteria was isolated from oligotrophic marine environments compared with coastal or estuarine environments (CitationJiang and Paul 1998), the contribution of viruses to biogeochemical nutrient cycles seems to be less significant in environments such as the pelagic ocean. The situation is not so simple in other environments, and CitationJiang and Paul (1996) showed that the percentage of lysogens that was estimated from prophage induction by mitomycin C, UV radiation, sunlight, temperature and pressure was highest in estuarine waters among estuarine, coastal and oligotrophic offshore environments. We have to remember that the existing lysogens are the remnants after factors such as detrimental temperature, pH, ionic strength or mutagens have already conducted a significant proportion of the host communities in a sample to prophage induction (CitationWeinbauer 2004). In summary, the contribution of lysogenic viruses in biogeochemical nutrient cycles compared with that of lytic viruses might depend on the degree of induction of lysogens by natural stresses in respective environments in time and space.
Lysogeny seems to be chosen in the evolutionary process by viruses as a survival strategy at low host abundance and activity (K-strategic). CitationWilcox and Fuhrman (1994) observed in coastal surface seawater that lytic infection occurred only when the product of viral and bacterial numbers equaled 1012 mL−1 or more. However, because of the specificity of virus–host interactions, infection is also dependent on bacterial species composition and diversity. This value may be different in soil environments because the threshold value was determined mainly by the frequency of viral encounters with bacterial hosts in seawater (CitationWilcox and Fuhrman 1994). Higher values may be expected from the adsorption of most viruses on soil particles, resulting in the restriction in the free movement of viruses to encounter their hosts, and lower values from the accumulation of viruses and hosts on soil particles.
The microbial world in soils is very different from that in aquatic environments in terms of composition and turnover time of microorganisms. Fungal biomass is often larger than that of bacteria in soils (CitationAnderson and Domsch 1980). And the turnover time of microbial communities is, on average, 1 year or longer in undisturbed soils (CitationJenkinson and Rayner 1977; CitationWhitman et al. 1998). No attention has been paid to the viral mortality of soil microorganisms that may contribute to the determination of turnover time of soil microorganisms. Moreover, as higher plants are the primary producer and the microbial loop, if any, is driven predominantly by heterotrophs growing on plant residues and humic substances in soils, the significance of the microbial loop in biogeochemical nutrient cycles in soil and the roles that viruses may play in it are different from those in aquatic environments.
DIVERSITY OF VIRUSES AND THEIR IMPORTANCE AS GENOMIC RESERVOIRS IN AQUATIC ENVIRONMENTS
Until recently, direct TEM examination of aquatic viruses was the most common method of recording viral diversity in aquatic environments. Although morphological data give only a limited view of viral diversity, these data have been cited as evidence that bacteriophages comprise the majority of viruses within viral communities. Many observations of temporal and spatial changes in the frequency distribution of viral capsid sizes have been recorded for viral populations (CitationWommack and Colwell 2000). Now evaluation of viral diversity and exploration for novel genes in aquatic environments from genomic analyses open a new view of aquatic virus communities, the studies of which are the central topics of “the third age of virus ecology”.
Size distribution of viruses
Viruses in aquatic environments are generally dominated by the 30–60-nm capsid size class (CitationCochlan et al. 1993; CitationProctor 1997; CitationWeinbauer 2004; CitationWommack and Colwell 2000), as shown in . The dominant viruses in the floodwater of paddy fields also fell into a similar size class (CitationNakayama et al. 2007b). Size distribution changes temporally and spatially from coastal to open ocean sites (CitationCochlan et al. 1993), with depth (CitationCochlan et al. 1993), and over a 1-month sampling period during spring diatom blooms (CitationBratbak et al. 1990; CitationMaranger et al. 1994). A difference in viral size class distribution was also noted among water samples collected from 22 Canadian lakes (CitationMaranger and Bird 1995).
Table 4 Size distribution (%) of viral communities in various aquatic environments
Not every study has documented the change in the distribution of capsid size classes with a change in time or the location of the water samples collected. CitationMathias et al. (1995) noted no change in the frequency of four classes of capsid diameters among the virioplankton in river water samples collected over a 2-year period. CitationNakayama et al. (2007b) also did not find a change in capsid size distribution in the floodwater of a Japanese paddy field under different fertilizer treatments (no fertilization; chemical N, P and K fertilizers; chemical N, P, K and Ca fertilizers; and compost with chemical N, P, K and Ca fertilizers) during the entire period of field flooding from transplanting to harvesting.
CitationWeinbauer and Peduzzi (1994) showed for Adriatic Sea water samples that the 30–60-nm capsid size class comprised 74 and 100% of intracellular viruses in rods and spirilla, respectively, whereas cocci more often contained larger viruses (60–110 nm) than the smaller 30–60 nm viruses (65%vs 35%) in the cells. This finding is indicative of the predominance of the 30–60-mn capsid size class of viruses in the marine environment because bacterial communities are predominated by rod-shaped members (CitationBratbak et al. 1990; CitationMathias et al. 1995; CitationWeinbauer and Peduzzi 1994; CitationWeinbauer et al. 1993).
The predominance of viruses within the 30–60-nm size class is attributed to the predominance of phages in viral communities and the selective predation of larger viruses by heterotrophic nanoflagellates. The average capsid size for viruses of eukaryotic algae is reported to be 152 nm, although 28% of them are less than 60 nm (CitationVan Etten et al. 1991). In general, phages are smaller, ranging from 34 to 160 nm with a peak sharply at 60 nm based on 251 phages (CitationAckermann 1998). CitationAckermann and DuBow (1987b) also found that phages with isometric capsids have a clear preference for the 55–65-nm range from approximately 180 species descriptions of classified phages of various bacteria. Viruses with different capsid sizes were ingested at different rates, with the smallest virus being ingested at the slowest rate (CitationGonzález and Suttle 1993). And these researchers calculated that when there are 106 bacteria mL−1 and 107–108 viruses mL−1, viruses may represent 0.2–9% of the carbon, 0.3–4% of the nitrogen and 0.6–28% of the phosphorus that the flagellates obtain from the ingestion of bacteria (CitationGonzález and Suttle 1993).
The clonal diversity of viral communities is evaluated on genome size using pulsed-field gel electrophoresis (PFGE), although the resolution of PFGE is not very high and the number of bands on the agarose gel is a conservative estimation of the number of viral species (CitationWeinbauer 2004). CitationWommack et al. (1999a) used PFGE to monitor the population dynamics of Chesapeake Bay virioplankton for an annual cycle. The PFGE analysis detected several distinct bands ranging from 50 to 300 kb, and statistical analyses elucidated changes in virioplankton community structure in relation to sampling time, geographical location and the extent of water column stratification. And the hybridization of PFGE-separated samples with DNA probes specific to single viral strains and a group of viruses with similar genome sizes demonstrated that the abundances of specific viruses changed in time and space in Chesapeake Bay (CitationWommack et al. 1999b).
Morphology of viruses
Although most studies on the morphology of free viruses in aquatic environments did not present quantitative data on the proportions of tailed versus non-tailed forms, most free viruses in aquatic environments appear to be predominantly tailed forms (CitationProctor 1997). The exception was the study by CitationTapper and Hicks (1998), in which they recorded the presence or absence of tails as well as capsid size distribution in water samples from Lake Superior, Canada (). On average, 70% of viruses were tailed and the proportion of tailed viruses increased with an increase in the capsid size from 65, 74 to 100% for those with < 30-nm, 30–60-nm and > 60-nm capsid diameters, respectively.
In six Delaware soils studied by CitationWilliamson et al. (2005), most soil viruses were phages belonging to Caudovirales. Among the tailed phages, podophages and siphophages were more dominant than myophages in these soils. In addition, a much higher frequency of filamentous and elongated capsids was found in these soils. In particular, approximately 10% of the viral community in a silt loam soil consisted of elongated capsid phages. The predominance of tailed viruses in aquatic and soil environments is natural because 96% of phages were tailed among over 5100 phages examined by TEM (CitationAckermann 1998, Citation2001, Citation2003). It is important to note that non-tailed forms may include free viruses that have lost their tails during the preparation for TEM observation.
Genomic diversity in aquatic viral communities
Although Siphoviridae or phages with long, non-contractile tails comprised 61% of tailed phages in the examinations using TEM since 1959 (CitationAckermann 1998, Citation2001, Citation2003), a predominance of Siphoviridae among tailed viruses is not necessarily the case in marine environments. CitationBreitbart et al. (2002, Citation2004a) examined a viral-community DNA library created from the Mission Bay sediment, California, and from two marine waters at Mission Bay and Scripps Pier, California, using linker-amplified shotgun cloning. They found a predominance of T7-like podophages, λ-like siphophages and T4-like myophages in two marine waters and λ-like siphophages, T7-like podophages and T4-like myophages in the sediment, in this order, among clones significantly similar to previously reported phage sequences, although more than 65% of the sequences were not affiliated with any known sequences. In contrast, a majority of phage isolates from marine environments were generally myophages and siphophages (CitationDemuth et al. 1993; CitationLu et al. 2001; CitationSullivan et al. 2003; CitationSuttle and Chan 1993; CitationWaterbury and Valois 1993; CitationWichels et al. 1998).
Until recently, it was commonly understood among viral taxonomists that there is no single gene that is common to all virus genomes and that a total diversity of uncultured virus communities cannot be evaluated using approaches analogous to 16S ribosomal DNA as for bacteria (CitationRohwer and Edwards 2002). However, the genome of a lytic phage P60 of marine Synechococcus WH7803 contained approximately 47,872 bp with 80 potential open reading frames that were mostly similar to the genes found in lytic podophages (T7, φYeO3-12 and SIO1). In addition, according to the 109 bacteriophage genomic databases available in 2001, lysogenic phage was the dominant form among the known myophage and siphophage genomes and integrase genes were found in all the lysogenic myophage and siphophage genomes, whereas eight of nine podophage genomes contained the DNA polymerase, primase and helicase genes (CitationChen and Lu 2002). It was also noted that three Prochlorococcus phages (a podophage and two myophages) were quite similar to T7-like (P-SSP7) and T4-like (P-SSM2 and P-SSM4) phages containing 15 of 26 core T7-like genes and 43 and 42 of 75 core T4-like genes, respectively (CitationSullivan et al. 2005). These findings indicate that comparable genomic information is preserved among viral subsets and that genomic information may be an effective tool for the phylogenetic classification of viruses and the phylogenetic evaluation of viral diversity in the environment (CitationWeinbauer and Rassoulzadegan 2004). Metagenomic analyses are elucidating the remarkable diversity of environmental viral communities, and CitationBreitbart and Rohwer (2005) estimated that there were possibly 5000 viral genotypes in 200 L of seawater and one million different viral genotypes in 1 kg of marine sediment.
Furthermore, some functional gene sequences in the host bacterial genomes were found to be highly conserved in viral genomes, examples of which are the DNA polymerase gene pol (CitationChen and Lu 2002; CitationCulley et al. 2003), the photosynthesis genes psbA and hliP (CitationLindell et al. 2004; CitationSullivan et al. 2005), the aldolase family gene talC (CitationSullivan et al. 2005), and the phosphate-inducible genes phoH and pstS (CitationSullivan et al. 2005). These findings substantiate the possibility of phylogenetic classification of viral subsets in the environment by comparing these genomic sequences. CitationRohwer and Edwards (2002) presented the “Phage Proteomic Tree” based on the overall similarity of 105 completely sequenced phage genomes. The resulting taxonomy was compatible with the system of International Committee on the Taxonomy of Viruses (ICTV), indicating that phage taxonomy has entered the post-genomic era. The presence of common functional genes related to fundamental physiological processes, energy-acquiring processes and gene expression processes across virus, prokaryote and eukaryote kingdoms strongly indicates horizontal gene transfer among the kingdoms, which may contribute to an increase in the biodiversity of respective genes via transduction. In addition, phage-encoded virulence factors are found in a variety of phages, members of Myoviridae, Siphoviridae, Podoviridae and Inoviridae, with some phages having characteristics of more than one family. A phage–bacterium interaction is, therefore, not a simple parasite–host interaction, but an instance of coevolution of phages and prokaryotic cells (CitationBoyd and Brüssow 2002).
The following sections provide brief summaries of the viral diversity in aquatic environments estimated from the structural genes (g20, g23) of T4-like phages, the polymerase gene (pol) of T7-like phages, and the photosynthesis gene (psbA) of marine cyanophages, as well as evidence of horizontal gene transfer among the living world.
Structure genes of T4-like phages
A marine cyanomyophage S-PM2 has a genome homology to coliphage T4 at a 10-kb region as a contiguous block from gene g18 to g23. In T4, g18 codes for the tail sheath, g19 for the tail tube, g20 for the head potal proteins, g21 for the prohead core protein, g22 for a scaffolding protein, and g23 for the major capsid protein (CitationHambly et al. 2001). The T4-like phage family was further classified into subgroups of the T-evens, PseudoT-evens, SchizoT-evens and ExoT-evens, with increasing divergence from T4 based on the sequence comparison of g18, g19 and g23 genes (CitationDeplats and Krisch 2003; CitationTétart et al. 2001).
The polymerase chain reaction (PCR) primers CPS1 and CPS2, which specifically amplify a 165-bp region from the majority of cyanomyophages, were constructed first by CitationFuller et al. (1998), although priming efficiency of the primers exhibit phage-to-phage variability. The region has significant similarity to g20 of coliphage T4. Primer pairs of CPS1 and CPS8 constructed by CitationZhong et al. (2002) cover more nucleotide sequences of g20 and elucidated the presence of nine phylogenetic groups among 114 totally different g20 homologs in six natural virus concentrates from estuarine and oligotrophic offshore environments. Only three groups/clusters contained known cyanophage isolates, and the identities of the other six clusters remain unknown.
The analysis of cyanophage communities by the g20 gene is also useful in the natural freshwater environment. CitationDorigo et al. (2004) studied cyanophage communities over time in Lake Bourget, France, using CPS1–CPS8 primers and found 35 distinct cyanomyophage g20 genotypes among 47 sequences analyzed. Phylogenetic analyses showed that these sequences fell into seven genetically distinct operational taxonomic units (OTUs). Some of these freshwater cyanophage sequences were genetically more closely related to marine cyanophage sequences than to other freshwater sequences. Similar findings of a wide distribution of closely related hosts and/or horizontal gene exchange among phage communities from very different environments were also obtained by analyzing g20 genes in various water samples from the Gulf of Mexico, the Arctic, the Southern, Northeast and Southeast Pacific Oceans, an Arctic cyanobacterial mat, a catfish production pond, lakes in Canada and Germany, and a depth of approximately 3246 m in the Chuckchi Sea (CitationShort and Suttle 2005). These researchers also found four novel phylogenetic groups of g20 genes, among which two were only found in freshwater.
In addition, CitationMühling et al. (2005) examined the control of Synechococcus genotypes by phages in the oligotrophic Gulf of Aquba, Red Sea, over an annual cycle from denaturing gradient gel electrophoresis (DGGE) patterns of a 118-bp g20 gene fragment of cyanophages and Restriction Fragment Length Polymorphism (RFLP) patterns of a 403-bp rpoC1 gene fragment of Synechococcus spp., in which they found that both the abundance and genetic diversity of cyanophage communities covaried with those of the Synechococcus communities. CitationWilhelm et al. (2006) observed the pervasive distribution of cyanophages in Lake Erie, USA, which can infect the marine Synechococcus sp. strain WH7803, and analyses of g20 indicate that these phages are related to marine cyanophages, but in some cases form a unique clade, leaving questions with regard to the natural hosts of these phages.
CitationFilée et al. (2005) compared the phylogenies constructed from g23 gene segment sequences with those obtained from the T4-type phage genomes for 16 completely sequenced T4-type phages and found a very good similarity between the phylogenies. Therefore, they designed degenerate primers targeting the g23 gene of phage T4 (MZIA1 bis, MZIA6) and applied them to elucidate T4-like bacteriophage communities in diverse marine environments (fjords and bays of British Columbia, the eastern Gulf of Mexico, and the western Arctic Ocean). Although some of the sequences of the PCR products were closely related to well-studied subgroups of the T4-like phages, such as the T-evens, SchizoT-evens, PseudoT-evens and ExoT-evens, the majority belonged to five previously uncharacterized subgroups. Thus, the g23 gene has more appropriate regions than the g20 gene for the phylogenetic evaluation of T4-type phage communities in the environment.
However, g23 genes may not be specific for T4-type phages. CitationJenkins and Hayes (2006) recently compared amino-acid sequences of g23 fragments among 17 cyanophage isolates of the heterocystous, filamentous cyanobacterium Nodularia spumigena using-specific primer set (CAP 1 and CAP 2). Although they were diverse in terms of their morphology and host range and belonged to two families of Myoviridae and Siphoviridae in Caudovirales, the encoded protein was 99% identical to T4 g23 homologues across all cyanophages compared. This fact indicates that g23 genes may be shared among Myoviridae and Siphoviridae members and that amino-acid sequences of the major capsid protein (g23) are important in phage–host interactions, irrespective of the phylogenetic positions of the phage and the host.
Primer pairs (MZIA1 bis, MZIA6) were also used by CitationJia et al. (2007) to compare DNA extracts of surface soil and rice straw collected from a Japanese paddy field during the flooded rice cultivation period. The g23 genes in these samples were quite distinctive in sequence from those obtained from marine environments (CitationFilée et al. 2005). Phylogenetic analysis showed that most of g23 sequences belonged to two novel subgroups of T4-type phages (Paddy Soil subgroup and Rice Straw subgroup), although some of them were distantly related to well-studied subgroups of T4-type phages, for example, exoT-evens, T-evens and Groups II, III and IV of marine clones (CitationJia et al. 2007). This finding strongly indicates that the virus communities in soil are different from those in marine environments and that soil environments store novel structural and functional genes of viral origins.
pol genes of T7-like phages
Eukaryotes, prokaryotes and some viruses possess B-family (α-like) DNA polymerases of exonuclease and polymerase domains with highly conserved amino acid sequence motifs (CitationBraithwaite and Ito 1993; CitationGrabherr et al. 1992; CitationIto and Braithwaite 1991). The conservation was at the amino-acid level, but not necessarily at the nucleotide level, and degenerate PCR primers (named AVS1 and AVS2) targeting algal-virus-specific pol genes were designed based on the successful amplification of viruses infecting Chlorella-like alga (Chlorophyceae), photosynthetic flagellate Micromonas pusilla (Prasinophyceae) and Chrysochromilina spp. (Prymnesiophyceae), although PCR products were not obtained from viruses infecting marine brown algae Ectocarpus siliculosis and Feldmannia sp. (Phaeophyceae) with the primers (CitationChen and Suttle 1995). The region of PCR amplification is at the catalytic site in the polymerase domain containing the most highly conserved amino acid sequence, YGDTDS. Phylogenetic trees of 13 microalgal viruses based on DNA pol sequences between AVS1 and AVS2 and on hybridization of total genomic DNA showed similar branching patterns, indicating that DNA pol sequences can be used to determine genetic relatedness and to infer phylogenetic relationships among these viruses. In addition, the phylogenetic tree constructed from the deduced amino-acid sequences of DNA pol genes of 24 dsDNA viruses, including phycodnaviruses, herpesviruses, poxviruses, baculoviruses and African swine fever virus, corresponded well with groupings based on the ICTV system (CitationChen and Suttle 1996).
CitationChen et al. (1996) applied these primers to the virus-sized fraction obtained from inshore and offshore water samples of the Gulf of Mexico and obtained five different genotypes or OTUs that were identified on the basis of RFLP-banding patterns, all of which belonged to the family Phycodnaviridae. The primers were applicable to DGGE analysis to rapidly analyze the PCR products of natural marine viral communities with resultant elucidation of spatial and temporal differences in algal-virus community structure (CitationShort and Suttle 1999). Primers of AVS1 and AVS2 were also used for DGGE analysis of geographically isolated natural algal-virus communities from coastal sites in the Pacific Ocean in British Columbia, Canada, and the Southern Ocean near the Antarctic Peninsula. Of the 33 sequences of different DGGE bands, 25 successfully encoded pol gene fragments. Similar virus sequences (> 98% sequence identity) were recovered from British Columbia and Antarctica, demonstrating that closely related algal viruses occur in distant geographical locations (CitationShort and Suttle 2002). Thus, sequence analyses of structural genes and the pol gene in various viral communities suggest that viral diversity could be high on a local scale, but relatively limited globally (CitationBreitbart and Rohwer 2005).
Recently, primers (T7DPol230F, T7DPol510R) targeting pol genes in T7-like podophages were constructed to compare T7-like podophage communities among different environments (CitationBreitbart et al. 2004b). DNA pol sequences of T7-like podophages occurred in all biomes investigated, including marine, estuarine, freshwater, sediment, terrestrial, hypersaline and metazoan-associated ones. The majority of these sequences belonged to a unique Polymerases from Uncultured Podophages (PUP) clade, distantly relating to cultured isolates. Some pol genes from this clade were > 99% conserved at the nucleotide level in multiple different environments, suggesting movement of these phages between biomes in recent evolutionary time (CitationBreitbart et al. 2004b). These findings indicate that pol genes are diverse with at least some pol genes included in the PUP clade and the others remote from the clade, although pol genes specific to respective environments remain unknown.
In addition, picorna-like viruses belonging to positive-sense ssRNA viruses are pathogens of penaeid shrimp, seals and whales. CitationCulley et al. (2003) designed primers (RdRp1, RdRp2) specific to RNA-dependent RNA polymerase (RdRp) sequence in picorna-like viruses, and applied the primers to assays for the presence of those viruses in the coastal waters of British Columbia, Canada. A diverse array of picorna-like viruses was found in the ocean, and all of the sequences were divergent from known picorna-like viruses falling within four monophyletic groups. Thus, polymerase sequences are well conserved among phylogenetic groups and this gene is an excellent molecular marker for examining the diversity of viruses in nature.
Analyses of phage communities in aquatic environments using primers specific to g20, g23 and pol genes elucidated the presence of abundant viruses belonging to clades distantly related to cultured isolates. This is because only a dozen phages are well characterized (CitationFrost et al. 2005), and they are limited to the members infecting specific bacterial hosts. Therefore, more effort should be paid to the isolation of phages infecting common, indigenous bacteria in the environment concurrently with the elucidation of phage genomics with these primers.
psbA genes of cyanophages
Many cyanobacteria are obligate photolithoautotrophs and have two different classes of reaction centers for photosynthesis: Type I and Type II centers (PS I and PS II). The PS II centers consist of two key proteins (D1 and D2 proteins) and are crucial sites of damage in photoinhibition. As a phage known as S-PM2 also encodes the D1 and D2 proteins by psbA and psbD genes, respectively, their expression in infected cells would allow a repair cycle to operate in PS II after the host's protein synthesis had been shut down by photoinhibition (CitationBailey et al. 2004; CitationMann et al. 2003). The transcription and translation of photosynthesis genes of phage origin in infected cells were elucidated by CitationLindell et al. (2005) and CitationClokie et al. (2006b). The transcripts of the psbA gene of phage S-PM2 appeared soon after infection of Synechococcus sp. WH7803 and remained at high levels until lysis, whereas a considerable transient increase in the abundance of the host psbA transcripts occurred shortly after infection with a subsequent decline to a lower level than the level without infection. The photosynthetic capacity of the cells remained constant throughout the course of infection (CitationClokie et al. 2006). In the Prochlorococcus MED-4 and podophage P-SSP7 system, phage psbA and high-light-inducible protein (hli) genes were expressed during infection and were cotranscribed with essential phage capsid genes. The phage D1 protein increased steadily over the infective period, whereas the expression of the host photosynthesis genes declined over the course of infection. In addition, replication of the phage genome was a function of photosynthesis (CitationLindell et al. 2005).
As primary production in the sea is dominated by cyanobacteria, the abundance and community structure of cyanophages may be critical factors in primary production in respective marine ecosystems (CitationLiu et al. 1997; CitationWilhelm and Suttle 1999). Primers (58-VDIDGIREP-66, 331-MHERNAHNFP-340) targeting conserved psbA genes were first constructed to elucidate the diversity of naturally occurring marine oxygenic picoplankton (CitationZeidner et al. 2003). The primers successfully amplified psbA gene fragments of both cyanobacterial groups and eukaryotic algae in the Red Sea and the Mediterranean Sea and also gave good separation of PCR products by DGGE (CitationZeidner and Béjà 2004; CitationZeidner et al. 2003).
Cyanophages belong to three families of tailed phages: Myoviridae, Siphoviridae and Podoviridae (CitationMann 2003; CitationSuttle 2000). Three phages from two families (Myoviridae and Podoviridae) that infect a marine cyanobacterium Prochlorococcus contained genes that encode D1 protein (PsbA) and high-light-inducible protein (Hli). A myophage encoded the second photosystem II core reaction center protein D2 (PsbD) as well, whereas the other myophage encoded the photosynthetic electron transport proteins, plastocyanin (PetE) and ferredoxin (PetF), which suggests that they encode functional proteins that may help maintain photosynthetic activity during infection (CitationLindell et al. 2004). Furthermore, the distributions of phage D1, D2 and Hli proteins to the same clusters with those from Prochlorococcus spp. indicate that those phage proteins are of cyanobacterial origin (CitationLindell et al. 2004). CitationMillard et al. (2004) also found that the phage psbA genes fall into a clade that includes the psbA genes from their potential Synechococcus and Prochlorococcus hosts, suggesting the idea of the acquisition of these genes through horizontal gene transfer from their hosts. In addition, the high degree of sequence identity of the psbAD cassettes of S-PM2 (obtained from the English Channel) and S-RSM88 (from the Gulf of Aqaba) suggests a fairly recent lateral transfer (CitationMillard et al. 2004). Although another conclusion seemed to be drawn from a subsequent study that phage psbA sequences form a separate clade from the clade of their host Synechococcus (CitationZeidner et al. 2005), the separation was indistinguishable when the psbA sequences of Prochlorococcus hosts and their phages were included (CitationHambly and Suttle 2005). Furthermore, possible exchange and reshuffling of psbA genes between Synechococcus and Prochlorococcus via phage intermediates were indicated (CitationZeidner et al. 2005).
Recently, CitationSullivan et al. (2006) isolated 33 Prochlorococcus and Synechococcus phages (six podophages, 25 myophages and two siphophages) and examined the presence of psbA and psbD genes in their genomes, including nine other published cyanophages. Eighty-eight percent of the phage genomes contained psbA, and 50% contained both psbA and psbD. The psbA gene was found in all myophages (n = 32) and Prochlorococcus podophages (n = 5), but not detected in Prochlorococcus siphophages (n = 2) and Synechococcus podophages (n = 3). psbD was found only in phages that contained psbA and only in myophages, but not in all psbA-containing myophages. In general, phages of a broad host range encoded both psbA and psbD. Furthermore, phylogenetic clustering patterns of psbA and psbD genes indicate that transfers of these genes were predominantly from Prochlorococcus to their phages and from Synechococcus to their phages (CitationSullivan et al. 2006).
CitationSullivan et al. (2006) also examined the genetic diversity of psbA and psbD genes retrieved offshore from Hawaii where Prochlorococcus cells commonly outnumber Synechococcus cells by orders of magnitude, by constructing the phylogenetic trees with the Prochlorococcus and Synechococcus isolates mentioned above. More than half of the psbA sequences (42 of 81) formed a large cluster with cultured Prochlorococcus podophages, and other psbA sequences formed subclusters that also contained cultured Prochlorococcus myophages. Thus, cyanophage culture collections represented much of the naturally occurring Prochlorococcus cyanophage psbA gene sequence diversity of that area. In contrast, all psbD sequences from the Hawaiian seawater samples formed a single well-supported cluster that included only a cultured Prochlorococcus cyanophage P-SSM4, and this cluster consisted of several subclusters without ever-known, cultured phages.
Thus, an analysis of phage psbAD sequences enables not only the elucidation of the phylogenetic relationships among cyanophages, but also suggests frequent horizontal gene exchange between cyanophages and their hosts. This progress in phylogenetical phage classification is within the phage members of Synechococcus and Prochlorococcus, mainly because these cyanobacteria are the main primary producers in marine and freshwater environments. Many species of cyanobacteria, different from the species in these aquatic environments, proliferate in paddy fields in the world. For example, Nostoc, Anabaena, Calothrix and Aulosira species are predominant among N2-fixing cyanobacteria in South and Southeast Asia (CitationKimura 2005). CitationHeckman (1979) identified 11 cyanophyta in rice fields in Udon Thani Province, Thailand, and CitationJutono (1973) identified seven genera of Nostocaceae, three genera of Rivulariaceae, three genera of Scytonemataceae, six genera of Chroococcaceae, and eight genera of Oscillatoriaceae in rice fields in the Jogjakarta district, Indonesia. Algal and cyanobacterial successions are a common occurrence in rice fields, for example, Spirogyra sp., Anabaena bharadwajae→A. bharadwajae→A. bharadwajae, Scytonema hofmanni and Nostoc passerianum in well-drained rice fields and A. bharadwajae→Scytonema coactile, Nostoc sp. and Aulosira fritschii in ill-drained lowland rice fields in India (CitationGupta 1966). Viruses infecting these cyanobacteria and algae are different from those infecting aquatic ones and may have very unique, varietal psbA genes. Rice field viruses provide a potentially rich source for the study of phylogenetic diversities of psbAD and other photosynthesis-related genes.
CONCLUSIONS FOR FUTURE STUDIES
Approximately 30% of the surface of the Earth is covered by land and the rest is covered by sea. However, the abundance of prokaryotes in the surface parts of the land/soils and the open ocean are estimated to comprise 2.55 and 1.18 × 1029 cells, respectively. Moreover, more abundant prokaryotes of 0.25–2.5 and 3.5 × 1030 cells may reside in the terrestrial subsurface and in the oceanic subsurface, respectively (CitationWhitman et al. 1998). On a cellular C basis, 26 × 109 ton-C in soil and 2.2 × 109 tC in the open ocean are estimated (CitationWhitman et al. 1998). In addition, as fungal biomass is often greater than that of bacteria in soils (CitationAnderson and Domsch 1980), this circumstantial evidence strongly suggests that soil viruses play a comparable and important role in biogeochemical nutrient cycles and as genomic reservoirs to those in the sea. Bacteriophages and fungal viruses may play decisive roles among soil viruses. The fungal state of the soil and the abundance of fungal viruses are, therefore, important in evaluating the role of viruses in biogeochemical nutrient cycles in soils.
The soil environment is very different to aquatic environments as a habitat for microorganisms and viruses, and the characteristics of soils may modify the roles of viruses in biogeochemical nutrient cycles and as genomic reservoirs. Soil consists of solid, liquid and gas phases, and the solid phase of sands, silts, clays and organic materials makes the pore space into countless, small compartments with sizes of nano- to milli-meter scales (soil structure). Soil structure results in the preparation of aerobic and anaerobic habitats for microorganisms, and these habitats are potentially stable enough for microorganisms to prevail there over several generations. The neighboring pore spaces in soil are generally isolated from each other by solid barriers, forming isolated independent habitats for soil microorganisms. Therefore, heterogeneity is far greater in soils than in aquatic environments as a habitat for microorganisms, indicating that soil may have more potential as a genomic reservoir. Meanwhile, the degree of viral adsorption on soil particles is generally more than 90% for all viruses (CitationBitton et al. 1978; CitationGoyal and Gerba 1979; CitationMoore et al. 1981, Citation1982), which strongly modulates their transmigration from one zone to another. The horizontal gene transfer mediated by viruses may be less common in soils than in aquatic environments.
Bulk soils are poor in substrates for microorganisms, which are generally in a dormant state and spend a major proportion of the energy obtained from substrate utilization not for proliferation, but rather for maintenance and survival. Furthermore estimated yearly C inputs for soil systems do not meet C for maintenance energy requirement by the total metabolizing soil populations by an order or more, indicating their survival in soils as resting/dormant state (CitationAnderson and Domsch 1985). Poor substrates in soils result in long turnover times of microbial communities in soils: more than 1 year in soils in contrast to aquatic environments in which turnover occurs within a few days (CitationJenkinson and Rayner 1977; CitationWeinbauer 2004; CitationWhitman et al. 1998; CitationWommack and Colwell 2000). No evaluation has been done on the effect of viral mortality of soil microorganisms in relation to turnover time. As lysogeny seems to have been chosen in the evolutionary process by viruses as the survival strategy at low host abundance and activity (K-strategic), viruses in bulk soils seem to be in a lysogenic state. And as host cells become more starved, the frequency of the pseudolysogenic life of viruses may increase in soils as well (CitationRipp and Miller 1998). The dominant lifestyle of viruses in soils needs to be elucidated for evaluating viral roles in biogeochemical nutrient cycles. Phages occurring in oligotrophic pelagic waters with low bacterioplankton abundance have a tendency toward polyvalency, whereas phages found in nutrient-rich and host-rich coastal waters are generally host specific (CitationWommack and Colwell 2000). The host range of indigenous soil viruses is an interesting subject for future research.
Changes in environmental parameters, such as moisture content, temperature, pH and aerobicity, are a common occurrence in soils by weather and field management for better crop production at a scale from days, months, seasons to years, which may induce lytic life cycles to lysogens in soils. In paddy field ecosystems, flooding the fields makes the soils anaerobic. Microorganisms in soil adapt to such environmental fluctuations by changing their physiology, for example, aerobic respiration under oxic conditions and denitrification under anoxic conditions for denitrifiers. A change in the physiology of the hosts may result in a change in the surface property of the host cells and the infectivity of viruses may change accordingly (CitationArchibald and Coapes 1976). Ecological studies on viral infection to hosts according to the physiological states of those hosts may be an interesting subject for soil virologists. In addition, phages of anaerobic bacteria, such as methanogens, have not been given much attention in past studies.
In soils, there are “hot spots” for microorganisms where they proliferate very actively. They are the habitats around plant roots (rhizosphere) and plant residues, where lytic life may be favorable for viruses (r-strategic). This is because the turnover time of hosts is short enough for lytic life of viruses, within a few days, and specific hosts grow preferentially. The application of organic fertilizers, such as composts, may afford additional hot spots to soil inhabitants. As it is well known that microorganisms in the rhizosphere and plant residues are different from those in bulk soil, the phylogenetic positions of the viruses infecting them may be different.
Microbial communities in soil are different from those in aquatic environments. For example, Synechococcus and Prochlorococcus occupy a large proportion of the prokaryote communities in the sea, but they are absent in communities in soil environments (CitationKimura 2005). Gram-positive bacteria account for most of the culturable soil bacteria (CitationLochhead 1940), whereas Gram-negative bacteria predominate in the hydrosphere such as groundwater, springs, streams and lakes (CitationRheinheimer 1974). It is well known that less than 1% of the cells observed by direct counting are generally recovered using standard cultivation methods (CitationAmann et al. 1995). The advent of molecular phylogenetic analysis enables a diversity analysis of the unculturable majority in the environment, resulting in elucidation of the high divergence of microbial communities in soil and marine environments and of the marked differences in community structure between them (e.g. CitationLiles et al. 2003; CitationTorsvik et al. 1990; CitationVenter et al. 2004). These findings expect the existence of novel viruses with novel genes characteristic to soil environments. The finding of novel clades of g23 genes from a paddy field may support this inference (CitationJia et al. 2007).
As a first step in the third age of virus ecology in soils, the development of reliable extraction methods of viruses is an immediate necessity. It is a prerequisite for evaluating the contribution of soil viruses to global biogeochemical nutrient cycles. At the same time, soil microbiologists need to redirect their interest to synecology of viral communities from autecology of specific viruses. The methodologies for the community analysis are common and familiar to soil microbiologists, for example, molecular approaches to conserved structure/functional genes of specific viral groups. Phylogenetic study on viral communities in soils is important to assess horizontal gene transfer on a global scale. Detection of comparable genes in soils to those found in aquatic environments expands the gene transfer from the scale within aquatic environments to the whole planet. Thus, the third age of virus ecology in oceanography and limnology invites soil microbiologists to cooperate together to elucidate pertinent global issues in biogeochemistry and evolution biology.
ACKNOWLEDGMENT
The authors sincerely thank Professor Guenther Stotzky, New York University, for his invaluable and inspiring comments and his incredibly patient English correction of our manuscript. Without his encouragement, this manuscript would not have been accomplished.
Notes
Present address: Max-Planck-Institute for Terrestrial Microbiology, Karl-von-Frisch-Straße, 35043 Marburg, Germany
Related Research Data
REFERENCES
- Abebe , HM , Sadowsky , MJ , Kinkle , BK and Schmidt , EL . 1992 . Lysogeny in Bradyrhizobium japonicum and its effect on soybean nodulation . Appl. Environ. Microbiol , 58 : 3360 – 3366 .
- Ackermann , HW . 1998 . Tailed bacteriophages: the order Caudovirales . Adv. Virus Res , 51 : 135 – 201 .
- Ackermann , HW . 2001 . Frequency of morphological phage descriptions in the year 2000 . Arch. Virol , 146 : 843 – 857 .
- Ackermann , HW . 2003 . Bacteriophage observations and evolution . Res. Microbiol , 154 : 245 – 251 .
- Ackermann , HW and DuBow , MS . 1987a . Viruses of Prokaryotes, Vol. 1. General Properties of Bacteriophages , Vol. 1 , Boca Raton : CRC Press .
- Ackermann , HW and DuBow , MS . 1987b . Viruses of Prokaryotes, Vol. 2. Natural Groups of Bacteriophages , Vol. 2 , Boca Raton : CRC Press .
- Adams , MH . 1959 . Bacteriophages , New York : Interscience Publishers .
- Alexander , M . 1981 . Why microbial predators and parasites do not eliminate their prey and hosts . Ann. Rev. Microbiol , 35 : 113 – 133 .
- Alonso , MC , Jimenez-Gomez , F , Rodriguez , J and Borrego , JJ . 2001 . Distribution of virus-like particles in an oligotrophic marine environment (Alboran Sea, western Mediterranean) . Microb. Ecol , 42 : 407 – 415 .
- Amann , RI , Ludwig , W and Schleifer , KH . 1995 . Phylogenetic identification and in situ detection of individual microbial cells without cultivation . Microbiol. Rev , 59 : 143 – 169 .
- Anderson , JPE and Domsch , KH . 1980 . Quantities of plant nutrients in the microbial biomass of selected soils . Soil Sci , 130 : 211 – 216 .
- Anderson , TH and Domsch , KH . 1985 . Maintenance carbon requirements of actively-metabolizing microbial populations under in situ conditions . Soil Biol. Biochem , 17 : 197 – 203 .
- Archibald , AR and Coapes , HE . 1976 . Bacteriophage SP50 as a marker for cell wall growth in Bacillus sublitis . J. Bacteriol , 125 : 1195 – 1206 .
- Ashelford , K , Day , MJ , Bailey , MJ , Lilley , AK and Fry , JC . 1999 . Insitu population dynamics of bacterial viruses in a terrestrial environment . Appl. Environ. Microbiol , 65 : 169 – 174 .
- Ashelford , K , Norris , SJ , Fry , JC , Bailey , MJ and Day , MJ . 2000 . Seasonal population dynamics and identifications of competing bacteriophages and their host in the rhizosphere . Appl. Environ. Microbiol , 66 : 4193 – 4199 .
- Azam , F , Fenchel , T , Field , JG , Gray , JS , Meyer-Reil , LA and Thingstad , F . 1983 . The ecological role of water-column microbes in the sea . Mar. Ecol. Prog. Ser , 10 : 257 – 263 .
- Azuaga , MJ , Muñoz , J , González , F and Arias , JM . 1990 . Isolation and characterization of bacteriophages from Myxococcus virescens . Microbios , 61 : 83 – 88 .
- Babich , H , Schiffenbauer , M and Stotzky , G . 1983 . Sensitivity of coliphage T1 to nickel in fresh and salt waters . Curr. Microbiol , 8 : 101 – 105 .
- Babich , H and Stotzky , G . 1978a . Influence of pH on inhibition of bacteria, fungi, and coliphages by bisulfite and sulfite . Environ. Res , 15 : 405 – 417 .
- Babich , H and Stotzky , G . 1978b . Toxicity of zinc to fungi, bacteria, and coliphages: influence of chloride ions . Appl. Environ. Microbiol , 36 : 906 – 914 .
- Babich , H and Stotzky , G . 1979 . Differential toxicities of mercury to bacteria and bacteriophages in sea and in lake water . Can. J. Microbiol , 25 : 1252 – 1257 .
- Babich , H and Stotzky , G . 1980 . Reductions in inactivation rates of bacteriophage by clay minerals in lake water . Water Res , 14 : 185 – 187 .
- Bailey , S , Clokie , MRJ , Millard , A and Mann , NH . 2004 . Cyanophage infection and photoinhibition in marine cyanobacteria . Res. Microbiol , 155 : 720 – 725 .
- Bales , RC , Gerba , CP , Grondin , GH and Jensen , SL . 1989 . Bacteriophage transport in sandy soil and fractured tuff . Appl. Environ. Microbiol , 55 : 2061 – 2067 .
- Bales , RC , Li , S , Maguire , KM , Yahya , MT and Gerba , CP . 1993 . MS-2 and poliovirus transport in porous media: hydrophobic effects and chemical perturbations . Water Resour. Res , 29 : 957 – 963 .
- Bergh , O , Børsheim , KY , Bratbak , G and Heldal , M . 1989 . High abundance of viruses found in aquatic environments . Nature , 340 : 467 – 468 .
- Bird , DF , Juniper , SK , Ricciardi-Rigault , M , Martineu , P , Prairie , YT and Calvert , SE . 2001 . Subsurface viruses and bacteria in Holocene/Late Pleistocene sediments of Saanich Inlet, BC: ODP holes 1033B and 1034B, leg 169S . Mar. Geol , 174 : 227 – 239 .
- Bishop , PE , Supiano , MA and Brill , WJ . 1977 . Technique for isolating phage for Azotobacter vinelandii . Appl. Environ. Microbiol , 33 : 1007 – 1008 .
- Bitton , G , Pancorbo , OC , Overman , AR and Gifford , GE . 1978 . Retention of viruses during sludge application to soils . Prog. Water Technol , 10 : 597 – 606 .
- Boehme , J , Fischer , ME Jiang , SC . 1993 . Viruses, bacterioplankton, and phytoplankton in the southeastern Gulf of Mexico: distribution and contribution to oceanic DNA pools . Mar. Ecol. Prog. Ser , 97 : 1 – 10 .
- Børsheim , KY . 1993 . Native marine bacteriophages . FEMS Microbiol. Ecol , 102 : 141 – 159 .
- Boyd , EF and Brüssow , H . 2002 . Common themes among bacteriophage-encoded virulence factors and diversity among the bacteriophages involved . Trends Microbiol , 10 : 521 – 529 .
- Bradley , DE . 1967 . Ultrastructure of bacteriophages and bacteriocins . Bacteriol. Rev , 31 : 230 – 314 .
- Braithwaite , D and Ito , J . 1993 . Compilation, alignment, and phylogenetic relationships of DNA polymerases . Nucleic Acids Res , 21 : 787 – 802 .
- Bratbak , G and Heldal , M . 1993 . “ Total count of viruses in aquatic environments ” . In Aquatic Microbial Ecology , Edited by: Kemp , PF , Sherr , BF , Sherr , EB and Cole , JJ . 135 – 138 . Boca Raton : Lewis Publishers .
- Bratbak , G , Heldal , M , Norland , S and Thingstad , TF . 1990 . Viruses as partners in spring bloom microbial trophodynamics . Appl. Environ. Microbiol , 56 : 1400 – 1405 .
- Bratbak , G , Thingstad , TF and Heldal , M . 1994 . Viruses and the microbial loop . Microb. Ecol , 28 : 209 – 221 .
- Breitbart , M , Felts , B Kelley , S . 2004a . Diversity and population structure of a near-shore marine-sediment viral community . P. Roy. Soc. Lond. B Bio , 271 : 565 – 574 .
- Breitbart , M , Miyake , JH and Rohwer , F . 2004b . Global distribution of nearly identical phage-encoded DNA sequences . FEMS Microbiol. Lett , 236 : 249 – 256 .
- Breitbart , M and Rohwer , F . 2005 . Here a virus, there a virus, everywhere the same virus? . Trends Microbiol , 13 : 278 – 284 .
- Breitbart , M , Salamon , P Andresen , B . 2002 . Genomic analysis of uncultured marine viral communities . Proc. Natl Acad. Sci. USA , 99 : 14250 – 14255 .
- Burge , WD and Enkiri , NK . 1978 . Virus adsorption by five soils . J. Environ. Qual , 7 : 73 – 76 .
- Burroughs , NJ , Marsh , P and Wellington , EMH . 2000 . Mathematical analysis of growth and interaction dynamics of streptomycetes and a bacteriophage in soil . Appl. Environ. Microbiol , 66 : 3868 – 3877 .
- Bystricky , V , Stotzky , G and Schiffenbauer , M . 1975 . Electron microscopy of T1-bacteriophage adsorbed to clay minerals: application of the critical point drying method . Can. J. Microbiol , 21 : 1278 – 1282 .
- Campbell , JIA , Albrechtsen , M and Sorensen , J . 1995 . Large Pseudomonas phages isolated from barley rhizosphere . FEMS Microbiol. Ecol , 18 : 63 – 74 .
- Canchaya , C , Fournous , G , Chibani-Chennoufi , S , Dillmann , ML and Brüssow , H . 2003a . Phage as agents of lateral gene transfer . Curr. Opin. Microbiol , 6 : 417 – 424 .
- Canchaya , C , Proux , C , Fournous , G , Bruttin , A and Brüssow , H . 2003b . Prophage genomics . Microbiol. Mol. Boil Rev , 67 : 238 – 276 .
- Carlander , A , Aronsson , P , Allestam , G , Stenström , TA and Perttu , K . 2000 . Transport and retention of bacteriophages in two types of willow-cropped lysimeters . J. Environ. Sci. Health , A35 : 1477 – 1492 .
- Carlson , GF Jr , Woodard , FE , Wentworth , DF and Sproul , OJ . 1968 . Virus inactivation on clay particles in natural waters . J. Water Poll. Control Fed , 40 : R98 – R106 .
- Casida , LE and Liu , KC . 1974 . Arthrobacter globiformis and its bacteriophage in soil . Appl. Microbiol , 28 : 951 – 959 .
- Chattopadhyay , S and Puls , RW . 2000 . Forces dictating colloidal interactions between viruses and soil . Chemosphere , 41 : 1279 – 1286 .
- Chen , F and Lu , J . 2002 . Genomic sequence and evolution of marine cyanophage P60: a new insight on lytic and lysogenic phages . Appl. Environ. Microbiol , 68 : 2589 – 2594 .
- Chen , F and Suttle , CA . 1995 . Amplification of DNA polymerase gene fragments from viruses infecting microalgae . Appl. Environ. Microbiol , 61 : 1274 – 1278 .
- Chen , F and Suttle , CA . 1996 . Evolutionary relationships among large double-stranded DNA viruses that infect microalgae and other organisms as inferred from DNA polymerase genes . Virology , 219 : 170 – 178 .
- Chen , F , Suttle , CA and Short , SM . 1996 . Genetic diversity in marine algal virus communities as revealed by sequence analysis of DNA polymerase genes . Appl. Environ. Microbiol , 62 : 2869 – 2874 .
- Chibani-Chennoufi , S , Bruttin , A , Dillmann , ML and Brüssow , H . 2004 . Phage–host interaction: an ecological perspective . J. Bacteriol , 186 : 3677 – 3686 .
- Clokie , MRJ , Millard , AD , Mehta , JY and Mann , NH . 2006a . Virus isolation studies suggest short-time variations in abundance in natural cyanophages populations of the Indian Ocean . J. Mar. Biol. Ass. UK , 86 : 499 – 505 .
- Clokie , MRJ , Shan , J Bailey , S . 2006b . Transcription of a “photosynthetic” T4-type phage during infection of a marine cyanobacterium . Environ. Microbiol , 8 : 827 – 835 .
- Cochlan , WP , Wikner , J , Steward , GF , Smith , DC and Azam , F . 1993 . Spatial distribution of viruses, bacteria and chlorophyll a in neritic, oceanic and estuarine environments . Mar. Ecol. Prog. Ser , 92 : 77 – 87 .
- Cochran , PK and Paul , JH . 1998 . Seasonal abundance of lysogenic bacteria in a subtropical estuary . Appl. Environ. Microbiol , 64 : 2308 – 2312 .
- Cresswell , N , Herron , PR , Saunders , VA and Wellington , EMH . 1992 . The fate of introduced streptomycetes, plasmid and phage populations in a dynamic soil system . J. Gen. Microbiol , 138 : 659 – 666 .
- Culley , AI , Lang , AS and Suttle , CA . 2003 . High diversity of unknown picorna-like viruses in the sea . Nature , 424 : 1054 – 1057 .
- Danovaro , R , Dell’anno , A , Trucco , A , Serresi , M and Vanucci , S . 2001 . Determination of virus abundance in marine sediments . Appl. Environ. Microbiol , 67 : 1384 – 1387 .
- Danovaro , R , Manini , E and Dell’Anno , A . 2002 . Higher abundance of bacteria than of viruses in deep Mediterranean sediments . Appl. Environ. Microbiol , 68 : 1468 – 1472 .
- Danovaro , R and Serresi , M . 2000 . Viral density and virus-to-bacterium ratio in deep-sea sediments of the eastern Mediterranean . Appl. Environ. Microbiol , 66 : 1857 – 1861 .
- Delisle , AL and Levin , RE . 1972 . Characteristics of three phages infectious for psychrophilic fishery isolates of Pseudomonas putrefaciens . Antonie van Leeuwenhoek , 38 : 1 – 8 .
- Demuth , J , Neve , H and Witzel , KP . 1993 . Direct electron microscopy study on the morphological diversity of bacteriophage populations in Lake Plußsee . Appl. Environ. Microbiol , 59 : 3378 – 3384 .
- Deplats , C and Krisch , HM . 2003 . The diversity and evolution of the T4-type bacteriophages . Res. Microbiol , 154 : 259 – 267 .
- Dorigo , U , Jacquet , S and Humbert , JF . 2004 . Cyanophage diversity, inferred from g20 gene analyses, in the largest natural lake in France, Lake Bourget . Appl. Environ. Microbiol , 70 : 1017 – 1022 .
- Dowd , SE , Pillai , SD , Wang , S and Corapcioglu , Y . 1998 . Delineating the specific influence of virus isoelectric point and size on virus adsorption and transport through sandy soils . Appl. Environ. Microbiol , 64 : 405 – 410 .
- Drake , LA , Choi , K-H , Haskell , AGE and Dobbs , FC . 1998 . Vertical profiles of virus-like particles and bacteria in the water column and sediments of Chesapeake Bay, USA . Aquat Microb. Ecol , 16 : 17 – 25 .
- Duboise , SM , Moore , BE , Sorber , CA and Sagik , BP . 1979 . Viruses in soil systems . Crit. Rev. Microbiol , 7 : 245 – 285 .
- Ducklow , HW , Purdie , DA , Williams , PJL and Davies , JM . 1986 . Bacterioplankton: a sink for carbon in a coastal marine plankton community . Science , 232 : 865 – 867 .
- Edwards , RA and Rohwer , F . 2005 . Viral metagenomics . Nat. rev. Microbiol. , 3 : 504 – 510 .
- El Didamony , G and Abdel-Fattah , GM . 1998 . Influence of VA mycorrhiza on growth and nodulation of Vicia faba in soil containing lytic phages . Egypt J. Microbiol , 33 : 153 – 166 .
- Erskine , JM . 1973 . Characteristics of Erwinia amylovora bacteriophage and its possible role in the epidemiology of fire blight . Can. J. Microbiol , 19 : 837 – 845 .
- Ezuka , A and Kaku , H . 2000 . A historical review of bacterial blight of rice . Bull. Natl Inst. Agrobiol. Resour. (Japan) , 15 : 1 – 207 .
- Fauquet , CM , Mayo , MA Maniloff , J , eds. 2005 . Virus Taxonomy: Eighth Report of the International Committee on Taxonomy of Viruses , 1259 London : Elsevier Academic Press .
- Filée , J , Tétart , F , Suttle , CA and Krisch , HM . 2005 . Marine T4-type bacteriophages, a ubiquitous component of the dark matter of the biosphere . Proc. Natl Acad. Sci. USA , 102 : 12471 – 12476 .
- Fischer , U and Velimirov , B . 2002 . High control of bacterial production by viruses in a eutrophic oxbow lake . Aquat. Microb. Ecol , 27 : 1 – 12 .
- Frost , LS , Leplae , R , Summers , AO and Toussaint , A . 2005 . Mobile genetic elements: the agents of open source evolution . Nat. rev. Microbiol. , 3 : 722 – 732 .
- Fuhrman , JA . 1992 . “ Bacterioplankton roles in cycling of organic matter: the microbial food web ” . In Primary Productivity and Biogeochemical Cycles in the Sea , Edited by: Palkowski , PG and Woodhead , AD . 361 – 383 . New York : Plenum .
- Fuhrman , JA . 1999 . Marine viruses and their biogeochemical and ecological effects . Nature , 399 : 541 – 548 .
- Fuhrman , JA and Suttle , CA . 1993 . Viruses in marine planktonic systems . Oceanography , 6 : 51 – 63 .
- Fuller , NJ , Wilson , WH , Joint , IR and Mann , NH . 1998 . Occurrence of a sequence in marine cyanophages similar to that of T4 g20 and its application to PCR-based detection and quantification techniques . Appl. Environ. Microbiol , 64 : 2051 – 2060 .
- Garza , RD and Suttle , CA . 1998 . The effect of cyanophages on the mortality of Synechococcus spp. and selection for UV resistant viral communities . Microb. Ecol , 36 : 281 – 292 .
- Gerba , CP , Wallis , C and Melnick , JL . 1975 . Fate of wastewater bacteria and viruses in sol . J. Irrig. Drain. E. ASCE , 101 : 157 – 174 .
- Germida , JJ and Casida , LE . 1981 . Isolation of Arthrobacter bacteriophage from soil . Appl. Environ. Microbiol , 41 : 1389 – 1393 .
- Germida , JJ and Khachatourians , G . 1988 . Transduction of Escherichia coli in soil . Can. J. Microbiol , 34 : 190 – 193 .
- González , JM and Suttle , CA . 1993 . Grazing by marine nanoflagellates on viruses and virus-sized particles: ingestion and digestion . Mar. Ecol. Prog. Ser , 94 : 1 – 10 .
- Goyal , SM and Gerba , CP . 1979 . Comparative adsorption of human enteroviruses, simian rotavirus, and selected bacteriophages to soils . Appl. Environ. Microbiol , 38 : 241 – 247 .
- Grabherr , R , Strasser , P and Van Etten , JL . 1992 . The DNA polymerase gene from Chlorella viruses PBCV-1 and NY-2A contains an intron with nuclear splicing sequences . Virology , 188 : 721 – 731 .
- Gross , DC , Powelson , ML , Regner , KM and Radamaker , GK . 1991 . A bacteriophage-typing system for surveying the diversity and distribution of strains of Erwinia carotovora in potato fields . Phytopathology , 81 : 220 – 226 .
- Gupta , P . 1966 . Algal flora and its importance in the economy of rice fields . Hydrobiologia , 28 : 213 – 222 .
- Hambly , E and Suttle , CA . 2005 . The viriosphere, diversity, and genetic exchange within phage communities . Curr. Opin. Microbiol , 8 : 444 – 450 .
- Hambly , E , Tétart , F , Desplats , C , Wilson , WH , Krisch , HM and Mann , NH . 2001 . A conserved genetic module that encodes the major virion components in both the coliphage T4 and the marine cyanophage S-PM2 . Proc. Natl Acad. Sci. USA , 96 : 11411 – 11416 .
- Hara , S , Koike , I , Terauchi , K , Kamiya , H and Tanoue , E . 1996 . Abundance of viruses in deep oceanic waters . Mar. Ecol. Prog. Ser , 145 : 269 – 277 .
- Hara , S , Terauchi , K and Koike , I . 1991 . Abundance of viruses in marine waters: assessment by epifluorescence and transmission electron microscopy . Appl. Environ. Microbiol , 57 : 2731 – 2734 .
- Hashem , FM , Kuykendall , LD , Udell , SE and Thomas , PM . 1996 . Phage susceptibility and plasmid profile analysis of Sinorhizobium fredii . Plant Soil , 186 : 127 – 134 .
- Heckman , CW . 1979 . Rice Field Ecology in Northeastern Thailand , The Hague : W. Junk B.V. Publishers .
- Hedal , M and Bratbak , G . 1991 . Production and decay of viruses in aquatic environments . Mar. Ecol. Prog. Ser , 72 : 205 – 212 .
- Hendrix , RW , Lawrence , JG , Hatfull , GF and Casjens , S . 2000 . The origins and ongoing evolution of viruses . Trends Microbiol , 8 : 504 – 508 .
- Hennes , KP and Simon , H . 1995 . Significance of bacteriophages for controlling bacterioplankton growth in a mesotrophic lake . Appl. Environ. Microbiol , 61 : 333 – 340 .
- Hennes , KP and Suttle , CA . 1995 . Direct counts of viruses in natural waters and laboratory cultures by epifluorescence microscopy . Limnol. Oceanogr , 40 : 1050 – 1055 .
- Hennes , KP , Suttle , CA and Chan , AM . 1995 . Fluorescently labeled virus probes show that natural virus populations can control the structure of marine microbial communities . Appl. Environ. Microbiol , 61 : 3623 – 3627 .
- Herron , PR and Wellington , EMH . 1994 . Population dynamics of phage–host interactions and phage conversion of streptomycetes in soil . FEMS Microbiol. Ecol , 14 : 25 – 32 .
- Hewson , I and Fuhrman , JA . 2003 . Viriobenthos production and virioplankton sorptive scavenging by suspended sediment particles in coastal and pelagic waters . Microb. Ecol , 46 : 337 – 347 .
- Hewson , I , O’Neil , JM , Fuhrman , JA and Dennison , WC . 2001 . Virus-like particle distribution and abundance in sediments and overlaying waters along eutrophication gradients in two subtropical estuaries . Limnol. Oceanogr , 46 : 1734 – 1746 .
- Hu , TL . 1998 . A comparison of two methods to recover phages from soil samples . Bioresource Technol , 65 : 167 – 169 .
- Hurst , CJ , Gerba , CP and Cech , I . 1980 . Effects of environmental variables and soil characteristics on virus survival in soil . Appl. Environ. Microbiol , 40 : 1067 – 1079 .
- Hussein , ME , Abou , El-Hawa , ME and El Dydamony , G . 1994 . Population and persistence of Zag-1 phage and cowpea Rhizobium in two sterile soils . Egypt J. Microbiol , 29 : 270 – 283 .
- Ito , J and Braithwaite , DK . 1991 . Compilation and alignment of DNA polymerase sequences . Nucleic Acids Res , 19 : 4045 – 4057 .
- Japanese Ministry of Agriculture Forestry . 1971 . Fundamental and Practical Standards of Survey for Crop Diseases and Insect Pests Forecasting Service , Tokyo : Ministry of Agriculture and Forestry . (in Japanese)
- Jenkins , CA and Hayes , PK . 2006 . Diversity of cyanophages infecting the heterocystous filamentous cyanobacterium Nodularia isolated from the brackish Baltic Sea . J. Mar. Biol. Assoc. UK , 86 : 529 – 536 .
- Jenkinson , DS and Ladd , JN . 1981 . “ Microbial biomass in soil: Measurement and turnover ” . In Siol Biochemistry , Edited by: Paul , EA and Ladd , JN . Vol. 5 , 415 – 471 . New York : Marcel Dekker .
- Jenkinson , DS and Rayner , JH . 1977 . The turnover of soil organic matter in some of the Rothamsted classical experiments . Soil Sci , 123 : 298 – 305 .
- Jia , Z , Ishihara , R , Nakajima , Y , Asakawa , A and Kimura , M . 2007 . Molecular characterization of T4-type bacteriophages in rice field . Environ. Microbiol , 9 : 1091 – 1096 .
- Jiang , SC and Paul , JH . 1994 . Seasonal and diel abundance of viruses and occurrence of lysogeny/bacteriocinogeny in the marine environment . Mar. Ecol. Prog. Ser , 104 : 163 – 172 .
- Jiang , SC and Paul , JH . 1996 . Occurrence of lysogenic bacteria in marine microbial communities as determined by prophage induction . Mar. Ecol. Prog. Ser , 142 : 27 – 38 .
- Jiang , SC and Paul , JH . 1998 . Significance of lysogeny in the marine-environment. Studies with isolates and a model of lysogenic phage production . Microb. Ecol , 35 : 235 – 243 .
- Jin , Y , Pratt , E and Yates , MV . 2000 . Effect of mineral colloids on virus transport through saturated sand columns . J. Environ. Qual , 29 : 532 – 539 .
- Jin , Y , Yates , MV , Thompson , SS and Jury , WA . 1997 . Sorption of viruses during flow through saturated sand columns . Environ. Sci. Technol , 31 : 548 – 555 .
- Jutono . 1973 . Blue-green algae in rice soils of Jogjakarta, central Java . Soil Biol. Biochem , 5 : 91 – 95 .
- Kamiunten , H and Wakimoto , S . 1979 . Biological properties of the filamentous phages released from Xanthomonas oryzae isolates . Ann. Phytopath. Soc. Jap , 45 : 174 – 181 .
- Kankila , J and Lindström , K . 1994 . Host range, morphology and DNA restriction patterns of bacteriophage isolates infecting Rhizobium leguminosarum bv. trifolii . Soil Biol. Biochem , 26 : 429 – 437 .
- Kapuscinski , RB and Mitchell , R . 1980 . Processes controlling virus inactivation in coastal waters . Water Res , 14 : 363 – 371 .
- Keel , C , Ucurum , Z , Michaux , P , Adrian , M and Haas , D . 2002 . Deleterious impact of a virulent bacteriophage on survival and biocontrol activity of Pseudomonas fluorescens strain CHA0 in natural soil . Mol. Plant Microbe In , 15 : 567 – 576 .
- Keswick , BH and Gerba , CP . 1980 . Viruses in groundwater . Environ. Sci. Technol , 14 : 1290 – 1297 .
- Kimura , M . 2005 . Populations, community composition and biomass of aquatic organisms in the floodwater of rice fields and effects of field management . Soil Sci. Plant Nutr , 51 : 159 – 181 .
- Kleczkowska , J . 1945 . A quantitative study of the interaction of bacteriophage with rhizobium using the technique of poured plates . J. Bacteriol , 50 : 81 – 94 .
- Kuo , TT , Huang , TC , Wu , RY and Yang , CM . 1967 . Characterization of three bacteriophages of Xanthomonas oryzae (Uyeda et Ishiyama) Dowson . Bot. Bull. Acad. Sinica , 8 : 246 – 254 .
- Lance , JC and Gerba , CP . 1984 . Effect of ionic composition of suspending solution on virus adsorption by a soil column . Appl. Environ. Microbiol , 47 : 484 – 488 .
- Lanning , S and Williams , ST . 1982 . Methods for the direct isolation and enumeration of actinophages in soil . J. Gen. Microbiol , 128 : 2063 – 2071 .
- Lechevalier , MP and Lechevalier , H . 1970 . Chemical composition as a criterion in the classification of aerobic actinomycetes . Int. J. Syst. Bacteriol , 20 : 435 – 443 .
- Lenski , RE . 1984 . Coevolution of bacteria and phage: are there endless cycles of bacterial defenses and phage counterdefences? . J. Theor. Biol , 108 : 319 – 325 .
- Lenski , RE . 1988 . Dynamics of interactions between bacteria and virulent bacteriophage . Adv. Microb. Ecol , 10 : 1 – 44 .
- Lenski , RE and Levin , BR . 1985 . Constraints on the coevolution of bacteria and virulent phage: a model, some experiments, and predictions for natural communities . Am. Nat , 125 : 585 – 602 .
- Leonardopoulos , J , Achladi-Aliferi , P and Papaconstantinou , A . 1996a . Survival of S. typhimurium in the soil in the presence of bacteriophages . Acta Microbiol. Hellenica , 41 : 586 – 597 . (in Greek with English summary)
- Leonardopoulos , J , Papaconstantinou , A and Georgakopoulou-Papandreou , E . 1996b . The meaning of soil characteristics and temperature for the survival of bacteriophages in naturally contaminated soil samples . Acta Microbiol. Hellenica , 41 : 309 – 316 . (in Greek with English summary)
- Levin , BR , Stewart , FM and Chao , L . 1977 . Resource-limited growth, competition, and predation: a model and experimental studies with bacteria and bacteriophage . Am. Nat , 111 : 3 – 24 .
- Liles , MR , Manske , BF , Bintrim , SB , Handelsman , J and Goodman , RM . 2003 . A census of rRNA genes and linked genomic sequences within a soil metagenomic library . Appl. Environ. Microbiol , 69 : 2684 – 2691 .
- Lindell , D , Jaffe , JD , Johnson , ZI , Church , GM and Chisholm , SW . 2005 . Photosynthesis genes in marine viruses yield proteins during host infection . Nature , 438 : 86 – 89 .
- Lindell , D , Sullivan , MB , Johnson , ZI , Tolonen , AC , Rohwer , F and Chisholm , SW . 2004 . Transfer of photosynthesis genes to and from Prochlorococcus viruses . Proc. Natl Acad. Sci. USA , 101 : 11013 – 11018 .
- Lipson , SM and Stotzky , G . 1983 . Adsorption of reovirus to clay minerals: Effects of cation-exchange capacity, cation saturation, and surface area . Appl. Environ. Microbiol , 46 : 673 – 682 .
- Lipson , SM and Stotzky , G . 1984a . “ Adsorption of viruses to particulates: possible effects on virus survival ” . In Viral Ecology , Edited by: Misra , AH and Polasa , H . 165 – 178 . New Delhi : South Asian Publishers .
- Lipson , SM and Stotzky , G . 1984b . Effect of proteins on reovirus adsorption to clay minerals . Appl. Environ. Microbiol , 48 : 525 – 530 .
- Lipson , SM and Stotzky , G . 1985a . Infectivity of reovirus adsorbed to homoionic and mixed-cation clays . Water Res , 19 : 227 – 234 .
- Lipson , SM and Stotzky , G . 1985b . Specificity of virus adsorption to clay minerals . Can. J. Microbiol , 31 : 50 – 53 .
- Lipson , SM and Stotzky , G . 1986 . Effect of kaolinite on the specific infectivity of reovirus . FEMS Microbiol. Lett , 37 : 83 – 88 .
- Lipson , SM and Stotzky , G . 1987 . “ Interactions between viruses and clay minerals ” . In Human Viruses in Sediments, Sludges, and Soils , Edited by: Rao , VC and Melnick , JL . 197 – 230 . Boca Raton : CRC Press .
- Liu , HB , Nolla , HA and Campbell , L . 1997 . Prochlorococcus growth rate and contribution to primary production in the equatorial and subtropical North Pacific Ocean . Aquat. Microb. Ecol , 12 : 39 – 47 .
- Lochhead , AG . 1940 . Qualitative studies of soil micro-organisms. III. Influence of plant growth on the character of the bacterial flora . Can. J. Res , 18 : 42 – 53 .
- Long , PF , Parekh , NR , Munro , JC and Williams , ST . 1993 . Isolation of actinophage that attack some maduromycete actinomycetes . FEMS Microbiol. Lett , 108 : 195 – 200 .
- Loveland , JP , Ryan , JN , Amy , GL and Harvey , RW . 1996 . The reversibility of virus attachment to mineral surfaces . Colloid Surfaces A , 107 : 205 – 221 .
- Lu , J , Chen , F and Hodson , RE . 2001 . Distribution, isolation, host specificity, and diversity of cyanophages infecting marine Synechococcus spp. in river estuaries . Appl. Environ. Microbiol , 67 : 3285 – 3290 .
- Mann , NH . 2003 . Phages of the marine cyanobacterial picophytoplankton . FEMS Microbiol. Rev , 27 : 17 – 34 .
- Mann , NH . 2005 . The third age of phage . PLoS Biol , 3 : 753 – 755 .
- Mann , NH , Cook , A , Millard , A , Bailey , S and Clokie , M . 2003 . Bacterial photosynthesis genes in a virus . Nature , 424 : 741
- Maranger , R and Bird , DF . 1995 . Viral abundance in aquatic systems: a comparison between marine and fresh waters . Mar. Ecol. Prog. Ser , 121 : 217 – 226 .
- Maranger , R , Bird , DF and Juniper , SK . 1994 . Viral and bacterial dynamics in arctic sea ice during the spring algal bloom near Resolute, N.W.T., Canada . Mar. Ecol. Prog. Ser , 111 : 121 – 127 .
- Maranger , R and Bird , DG . 1996 . High concentrations of viruses in the sediments of Lake Gilbert, Québec . Microb. Ecol , 31 : 141 – 151 .
- Marie , D , Brussaard , CPD , Thyrhaug , R , Bratbak , G and Vaulot , D . 1999 . Enumeration of marine viruses in culture and natural samples by flow cytometry . Appl. Environ. Microbiol , 65 : 45 – 52 .
- Marsh , P and Wellington , EMH . 1994 . Phage–host interactions in soil . FEMS Microbiol. Ecol , 15 : 99 – 108 .
- Mathias , CB , Kirschner , AKT and Velimirov , B . 1995 . Seasonal variations of virus abundance and viral control of the bacterial production in a backwater system of the Danube River . Appl. Environ. Microbiol , 61 : 3734 – 3740 .
- McKenna , F , El-Tarabily , KA , Hardy , GESTJ and Dell , B . 2001 . Novel in vivo use of a polyvalent Streptomyces phage to disinfest Streptomyces scabies-infected seed potatoes . Plant Pathol , 50 : 666 – 675 .
- Meschke , JS and Sobsey , MD . 2003 . Comparative reduction of Norwalk viruses, poliovirus type 1, F+ RNA coliphae MS2 and in Escherichia coli miniature soil columns . Water Sci. Technol , 47 : 85 – 90 .
- Middelboe , M , Glud , RN and Finster , K . 2003 . Distribution of viruses and bacteria in relation to diagenetic activity in an estuarine sediment . Limnol. Oceanogr , 48 : 1447 – 1456 .
- Millard , A , Clokie , RJ , Shub , DA and Mann , NH . 2004 . Genetic organization of the psbAD region in phages infecting marine Synechococcus strains . Proc. Natl Acad. Sci. USA , 101 : 11007 – 11012 .
- Millard , AD and Mann , NH . 2006 . A temporal and spatial investigation of cyanophages abundance in the Gulf of Aqaba, Red Sea . J. Mar. Biol. Assoc. UK , 86 : 507 – 515 .
- Moce-Llivina , L , Muniesa , M , Pimenta-Vale , H , Lucena , F and Jofre , J . 2003 . Survival of bacterial indicator species and bacteriophages after thermal treatment of sludge and sewage . Appl. Environ. Microbiol , 69 : 1452 – 1456 .
- Moebus , K . 1987 . “ Ecology of marine bacteriophages ” . In Phage Ecology , Edited by: Goyal , SM , Gerba , CP and Bitton , G . 137 – 156 . New York : John Wiley and Sons .
- Moebus , K . 1996 . Marine bacteriophage reproduction under nutrient-limited growth of host bacteria. II. Investigations with phage–host system [H3:H3/1] . Mar. Ecol. Prog. Ser , 144 : 13 – 22 .
- Monpoeho , S , Maul , A , Mignotte-Cadiergues , B , Schwartzbrod , K , Billaudel , S and Ferre , V . 2001 . Best viral elution method available for quantification of enteroviruses in sludge by both cell culture and reverse transcription-PCR . Appl. Environ. Microbiol , 67 : 2484 – 2488 .
- Montanié , H , Hartmann , HJ , Crottereau , C and Trichet , C . 2002 . Virus-like particle analysis in a seston-rich coastal pond using transmission electron microscopy . Aquat. Microb. Ecol , 28 : 105 – 115 .
- Moore , RS , Taylor , DH , Reddy , MM and Sturman , LS . 1982 . Adsorption of reovirus by minerals and soils . Appl. Environ. Microbiol , 44 : 852 – 859 .
- Moore , RS , Taylor , DH , Sturman , LS , Reddy , MM and Fuhs , GW . 1981 . Poliovirus adsorption by 34 minerals and soils . Appl. Environ. Microbiol , 42 : 963 – 975 .
- Mühling , M , Fuller , NJ Millard , A . 2005 . Genetic diversity of marine Synechococcus and co-occurring cyanophage communities: evidence for viral control of phytoplankton . Environ. Microbiol , 7 : 499 – 508 .
- Nakayama , N , Okumura , M , Inoue , K , Asakawa , S and Kimura , M . 2007a . Seasonal variations in the abundance of virus-like particles and bacteria in the floodwater of a Japanese paddy field . Soil Sci. Plant Nutr , 53 : 420 – 429 .
- Nakayama , N , Okumura , M , Inoue , K , Asakawa , S and Kimura , M . 2007b . Morphological analysis of viral communities in the floodwater of a Japanese paddy field . Soil Biol. Biochem , doi:10.1016/j/soilbio.2007.06.025
- Noble , RT and Fuhrman , JA . 1997 . Virus decay and its causes in coastal waters . Appl. Environ. Microbiol , 63 : 77 – 83 .
- Noble , RT and Fuhrman , JA . 1998 . Use of SYBR Green I for rapid epifluorescence counts of marine viruses and bacteria . Aquat. Microb. Ecol , 14 : 113 – 118 .
- Nomizu , T and Mizuike , A . 1986 . Electron microscopy of submicron particles in natural waters – specimen preparation by centrifugation . Mikrochim. Acta , 1 : 65 – 72 .
- Novikova , NI and Limeschenko , EV . 1992 . The susceptibility of nodule bacteria with the unclear taxonomic position to bacteriophages of various Rhizobium genera . Mikrobiologiya , 61 : 484 – 489 . (in Russian with English summary)
- Novikova , NI , Pavlova , EA and Limeshchenko , EV . 1993 . Phage sensitivity and host range of Rhizobium strains isolated from root nodules of temperate legumes . Plant Soil , 151 : 45 – 53 .
- Ogunseitan , OA , Sayler , GS and Miller , RV . 1992 . Application of DNA probes to analysis of bacteriophage distribution patterns in the environment . Appl. Environ. Microbiol , 58 : 2046 – 2052 .
- Pantastico-Caldas , M , Duncan , KE , Istock , CA and Bell , JA . 1992 . Population dynamics of bacteriophage and Bacillus subtilis in soil . Ecology , 73 : 1888 – 1902 .
- Paul , JH , Jiang , SC and Rose , JB . 1991 . Concentration of viruses and dissolved DNA from aquatic environments by Vortex flow filtration . Appl. Environ. Microbiol , 57 : 2197 – 2204 .
- Paul , JH , Rose , JB , Jiang , SC , Kellog , CA and Dickons , L . 1993 . Distribution of viral abundance in the reef environment of Key Largo, Florida . Appl. Environ. Microbiol , 59 : 718 – 724 .
- Paul , JH , Sullivan , MB , Segall , AM and Rohwer , F . 2002 . Marine phage genomics . Comp. Biochem. Phys. B , 133 : 463 – 476 .
- Paul , JH and Sullivan , MS . 2005 . Marine phage genomics: what have we learned? . Curr. Opin. Biotechnol , 16 : 299 – 307 .
- Pina , S , Creus , A , González , N , Gironés , R , Felip , M and Sommaruga , R . 1998 . Abundance, morphology and distribution of planktonic virus-like particles in two high-mountain lakes . J. Plankton Res , 20 : 2413 – 2421 .
- Powelson , DK , Gerba , CP and Yahya , MT . 1993 . Virus transport and removal in wastewater during aquifer recharge . Water Res , 27 : 583 – 590 .
- Powelson , DK , Simpson , JR and Gerba , CP . 1991 . Effects of organic matter on virus transport in unsaturated flow . Appl. Environ. Microbiol , 57 : 2192 – 2196 .
- Proctor , LM . 1997 . Advances in the study of marine viruses . Microsc. Res. Techniq. , 37 : 136 – 161 .
- Reanney , DC and Marsh , SCN . 1973 . The ecology of viruses attacking Bacillus stearothermophilus in soil . Soil Biol. Biochem , 5 : 399 – 408 .
- Reiter , W , Zillig , W and Palm , P . 1988 . Archaebacterial viruses . Adv. Virus Res , 34 : 143 – 188 .
- Rheinheimer , G . 1974 . “ Aquatic bacteria ” . In Aquatic Microbiology , 23 – 42 . London : John Wiley & Sons .
- Ripp , S and Miller , RV . 1995 . Effects of suspended particulates on the frequency of transduction among Pseudomonas aeruginosa in a freshwater environment . Appl. Environ. Microbiol , 61 : 1214 – 1219 .
- Ripp , S and Miller , RV . 1997 . The role of pseudolysogeny in bacteriophage–host interactions in a natural freshwater environment . Microbiology , 143 : 2065 – 2070 .
- Ripp , S and Miller , RV . 1998 . Dynamics of the pseudolysogenic response in slowly growing cells of Pseudomonas aeruginosa . Microbiology , 144 : 2225 – 2232 .
- Rohwer , F and Edwards , R . 2002 . The phage proteomic tree: a genome based taxonomy for phage . J. Bacteriol , 184 : 4529 – 4535 .
- Roper , MM and Marshall , KCJ . 1978 . Effect of clay particle size on clay–Escherichia coli–bacteriophage interactions . J. Gen. Microbiol , 106 : 187 – 189 .
- Schaub , SA and Sorber , CA . 1977 . Virus and bacteria removal from wastewater by rapid infiltration through soil . Appl. Environ. Microbiol , 33 : 609 – 619 .
- Schiffenbauer , M and Stotzky , G . 1982 . Adsorption of coliphages T1 and T7 to clay minerals . Appl. Environ. Microbiol , 43 : 590 – 596 .
- Schiffenbauer , M and Stotzky , G . 1983 . Adsorption of coliphages T1 and T7 to host and non-host microbes and to clay minerals . Curr. Microbiol , 8 : 245 – 249 .
- Schijven , JF , Hassanizadeh , SM and de Bruin , HAM . 2002 . Column experiments to study nonlinear removal of bacteriophages by passage through saturated dune sand . J. Contam. Hydrol , 58 : 243 – 259 .
- Schrader , HS , Schrader , JO , Walker , JJ , Wolf , TA , Nickerson , KW and Kokjohn , TA . 1997 . Bacteriophage infection and multiplication occur in Pseudomonas aeruginosa starved for 5 years . Can. J. Microbol , 43 : 1157 – 1163 .
- Seeley , ND and Primrose , SB . 1982 . The isolation of bacteriophages from the environment . J. Appl. Bacteriol , 53 : 1 – 17 .
- Sharma , RS , Mohmmed , A and Babu , CR . 2002 . Diversity among rhizobiophages from rhizospheres of legumes inhabiting three ecogeographical regions of India . Soil Biol. Biochem , 34 : 965 – 973 .
- Sharp , RJ , Ahmad , SI , Munster , A , Dowsett , B and Atkinson , T . 1986 . The isolation and characterization of bacteriophages infecting obligately thermophilic strains of Bacillus . J. Gen. Microbiol , 132 : 1709 – 1722 .
- Short , CM and Suttle , CA . 2005 . Nearly identical bacteriophage structural gene sequences are widely distributed in both marine and freshwater environments . Appl. Environ. Microbiol , 71 : 480 – 486 .
- Short , SM and Suttle , CA . 1999 . Use of the polymerase chain reaction and denaturing gradient gel electrophoresis to study diversity in natural virus communities . Hydrobiologia , 401 : 19 – 32 .
- Short , SM and Suttle , CA . 2002 . Sequence analysis of marine virus communities reveals that groups of related algal viruses are widely distributed in nature . Appl. Environ. Microbiol , 68 : 1290 – 1296 .
- Sinton , LW , Finlay , RK , Pang , L and Scott , DM . 1997 . Transport of bacteria and bacteriophages in irrigated effluent into and through an alluvial gravel aquifer . Water Air Soil Poll , 98 : 17 – 42 .
- Smit , E , Wolters , AC , Lee , H , Trevors , JT and van Elsas , JD . 1996 . Interactions between a genetically marked Pseudomonas fluorescens strain and bacteriophage (R2f in soil: Effects of nutrients, alginate encapsulation, and the wheat rhizosphere . Microb. Ecol , 31 : 125 – 140 .
- Steward , GF , Smith , DC and Azam , F . 1996 . Abundance and production of bacteria and viruses in the Bering and Chukchi Seas . Mar. Ecol. Prog. Ser , 131 : 287 – 300 .
- Stotzky , G . 1985 . “ Mechanisms of adhesion to clays, with reference to soil systems ” . In Bacterial Adhesion: Mechanisms and Physiological Significance , Edited by: Savage , DC and Fletcher , MM . 195 – 253 . New York : Plenum Publishing Corporation .
- Stotzky , G . 1986 . “ Influence of soil mineral colloids on metabolic processes, growth, adhesion, and ecology of microbes and viruses ” . In Interactions of Soil Minerals with Natural Organics and Microbes , Edited by: Huang , PM and Schnitzer , M . 305 – 428 . Madison : Soil Science Society of America .
- Stotzky , G . 1989 . “ Gene transfer among bacteria in soil ” . In Gene Transfer in the Environment , Edited by: Levy , SB and Miller , RV . 165 – 222 . New York : McGraw-Hill .
- Stotzky , G , Devanas , MA and Zeph , LR . 1990 . Methods for studying bacterial gene transfer in soil by conjugation and transduction . Adv Appl. Microbiol , 35 : 57 – 169 .
- Stotzky , G , Schiffernbauer , M , Lipson , SM and Yu , BH . 1981 . “ Surface interactions between viruses and clay minerals and microbes: mechanisms and implications ” . In Viruses and Wastewater Treatment , Edited by: Goddard , M and Butler , M . 199 – 204 . Oxford : Pergamon Press .
- Stotzky , G , Zeph , LR and Devanas , MA . 1991 . “ Factors affecting the transfer of genetic information among microorganisms in soil ” . In Assessing Ecological Risks of Biotechnology , Edited by: Ginzburg , LR . 95 – 122 . Stoneham : Butterworth-Heinemann .
- Straub , TM , Pepper , IL and Gerba , CP . 1992 . Persistence of viruses in desert soils amended with anaerobically digested sewage sludge . Appl. Environ. Microbiol , 58 : 636 – 641 .
- Sullivan , MB , Coleman , ML , Weigele , P , Rohwer , F and Chisholm , SW . 2005 . Three Prochlorococcus cyanophage genomes: signature features and ecological interpretations . PLoS Biol , 3 : 790 – 806 .
- Sullivan , MB , Lindell , D , Lee , JA , Thompson , LR , Bielawski , JP and Chisholm , SW . 2006 . Prevalence and evolution of core photosystem II genes in marine cyanobacterial viruses and their hosts . PloS Biol , 4 : 1344 – 1357 .
- Sullivan , MB , Waterbury , JB and Chisholm , SW . 2003 . Cyanophages infecting the oceanic cyanobacterium Prochlorococcus . Nature , 424 : 1047 – 1051 .
- Suttle , CA . 1993 . “ Enumeration and isolation of viruses ” . In Aquatic Microbial Ecology , Edited by: Kemp , PF , Sherr , BF , Sherr , EB and Cole , JJ . 121 – 134 . Boca Raton : Lewis Publishers .
- Suttle , CA . 1994 . The significance of viruses to mortality in aquatic microbial communities . Microb. Ecol , 28 : 237 – 243 .
- Suttle , CA . 2000 . “ Cyanophages and their role in the ecology of cyanobacteria ” . In The Ecology of Cyanobacteria , Edited by: Whitton , BA and Potts , M . 563 – 589 . the Netherland : Kluwer Academic Publishers .
- Suttle , CA . 2005 . Viruses in the sea . Nature , 437 : 356 – 361 .
- Suttle , CA and Chan , AM . 1993 . Marine cyanophages infecting oceanic and coastal strains of Synechococcus: abundance, morphology, cross-infectivity and growth characteristics . Mar. Ecol. Prog. Ser , 92 : 99 – 109 .
- Suttle , CA and Chen , F . 1992 . Mechanisms and rates of decay of marine viruses in seawater . Appl. Environ. Microbiol , 58 : 3721 – 3729 .
- Sykes , IK , Lanning , S and Williams , ST . 1981 . The effect of pH on soil actinophage . J. Gen. Microbiol , 122 : 271 – 280 .
- Sykes , IK and Williams , ST . 1978 . Interactions of actinophage and clays . J. Gen. Microbiol , 108 : 97 – 102 .
- Szmelcman , S and Hofnung , M . 1975 . Maltose transport in Escherichia coli K-12: involvement of the bacteriophage Lambda receptor . J. Bacteriol , 124 : 112 – 118 .
- Tagami , Y . 1959 . Relationship between the occurrence of bacterial leaf blight of rice and the population of causal bacterium and its phage in rice-growing seasons . Plant Protection , 13 : 389 – 394 . (in Japanese)
- Tan , JSH and Reanney , DC . 1976 . Interactions between bacteriophages and bacteria in soil . Soil Biol. Biochem , 8 : 145 – 150 .
- Tapper , MA and Hicks , RE . 1998 . Temperate viruses and lysogeny in Lake Superior bacterioplankton . Limnol. Oceanogr , 43 : 95 – 103 .
- Tartera , C , Araujo , R , Michel , T and Jofre , J . 1992 . Culture and decontaminating methods affecting enumeration of phages infecting Bacteroides fragilis in sewage . Appl. Environ. Microbiol , 58 : 2670 – 2673 .
- Taylor , DH , Moore , RS and Sturman , LS . 1981 . Influence of pH and electrolyte composition on adsorption of poliovirus by soils and minerals . Appl. Environ. Microbiol , 42 : 976 – 984 .
- Tétart , F , Desplats , C , Kutateladze , K , Monod , C , Ackermann , HW and Krisch , HM . 2001 . Phylogeny of the major head and tail genes of the wide-ranging T4-type bacteriophages . J. Bacteriol , 183 : 358 – 366 .
- Torrella , F and Morita , RY . 1979 . Evidence by electron micrographs for a high incidence of bacteriophage particles in the waters of Yaquina Bay, Oregon: ecological and taxonomical implications . Appl. Environ. Microbiol , 37 : 774 – 778 .
- Torsvik , V , Goksøyr , J and Daae , FL . 1990 . High diversity in DNA of soil bacteria . Appl. Environ. Microbiol , 56 : 782 – 787 .
- Toyoda , H , Kakutani , K , Ikeda , S , Goto , S , Tanaka , H and Ouchi , S . 1991 . Characterization of deoxyribonucleic acid of virulent bacteriophage and its infectivity to host bacteria, Pseudomonas solanacearum . J. Phytopathol , 131 : 11 – 21 .
- Van Etten , JL , Lane , KC and Meints , RH . 1991 . Viruses and virus like particles of eukaryotic algae . Microbiol. Rev , 55 : 586 – 620 .
- Van Regenmortel , MHV . 2000 . “ Introduction to the species concept in virus taxonomy ” . In Virus Taxonomy: Classification and nomenclature of viruses. Seventh Report of the International Committee on Taxonomy of Viruses , Edited by: Van Regenmortel , MHV , Fauquet , CM Bishop , DHL . 3 – 16 . San Diego : Academic Press .
- Venter , JC , Remington , K Heidelberg , JF . 2004 . Environmental genome shotgun sequencing of the Sargasso Sea . Science , 304 : 66 – 74 .
- Vettori , C , Gallori , E and Stotzky , G . 2000 . Clay minerals protect bacteriophage PBS1 of Bacillus subtilis against inactivation and loss of transducing ability by UV radiation . Can. J. Microbiol , 46 : 770 – 773 .
- Vettori , C , Stotzky , G , Yoder , M and Gallori , E . 1999 . Interaction between bacteriophage PBS1 and clay minerals and transduction of Bacillus subtilis by clay–phage complexes . Environ. Microbiol , 1 : 347 – 355 .
- Wakimoto , S . 1967 . “ Strains of Xanthomonas oryzae in Asia and their virulence against rice varieties ” . In Proceedings of the Symposium on Rice Diseases and Their Control by Growing Resistant Varieties and Other Measures , 19 – 24 . Tokyo : Agriculture, Forestry and Fisheries Research Council, Ministry of Agriculture and Forestry .
- Ward , RL and Ashley , CS . 1976 . Inactivation of poliovirus in digested sludge . Appl. Environ. Microbiol , 31 : 921 – 930 .
- Waterbury , JB and Valois , FW . 1993 . Resistance to co-occurring phages enables marine Synechococcus communities to coexist with cyanophages abundant in seawater . Appl. Environ. Microbiol , 59 : 3393 – 3399 .
- Weinbauer , M , Brettar , I and Hofle , M . 2003 . Lysogeny and virus-induced mortality of bacterioplankton in surface, deep, and anoxic waters . Limnol. Oceanogr , 48 : 1457 – 1465 .
- Weinbauer , MG . 2004 . Ecology of prokaryotic viruses . FEMS Microbiol. Rev , 28 : 127 – 181 .
- Weinbauer , MG , Fuks , D and Peduzzi , P . 1993 . Distribution of viruses and dissolved DNA along a coastal trophic gradient in the northern Adriatic sea . Appl. Environ. Microbiol , 59 : 4074 – 4082 .
- Weinbauer , MG , Fuks , D , Puskaric , S and Peduzzi , P . 1995 . Diel, seasonal, and depth-related variability of viruses and dissolved DNA in the northern Adriatic Sea . Microb. Ecol , 30 : 25 – 41 .
- Weinbauer , MG and Höfle , MG . 1998 . Significance of viral lysis and flagellate grazing as factors controlling bacterioplankton production in a eutrophic lake . Appl. Environ. Microbiol , 64 : 431 – 438 .
- Weinbauer , MG and Peduzzi , P . 1994 . Frequency, size, and distribution of bacteriophages in different marine bacterial morphotypes . Mar. Ecol. Prog. Ser , 108 : 11 – 20 .
- Weinbauer , MG and Rassoulzadegan , F . 2004 . Are viruses driving microbial diversification and diversity? . Environ. Microbiol , 6 : 1 – 11 .
- Weinbauer , MG and Suttle , CA . 1996 . Potential significance of lysogeny to bacteriophage production and bacterial mortality in coastal waters of the Gulf of Mexico . Appl. Environ. Microbiol , 62 : 4374 – 4380 .
- Wellington , EMH and Williams , ST . 1981 . Host ranges of phages isolated to Streptomycetes and other genera . Zbl. Bakt. Mik. Hyg. Suppl , 11 : 7 – 16 .
- Whitman , WB , Coleman , DC and Wiebe , W . 1998 . Prokaryotes: The unseen majority . Proc. Natl Acad. Sci. USA , 95 : 6578 – 6583 .
- Wichels , A , Biel , SS , Gelderblom , HR , Brinkhoff , T , Muyzer , G and Schütt , C . 1998 . Bacteriophage diversity in the North Sea . Appl. Environ. Microbiol , 64 : 4128 – 4133 .
- Wilcox , RM and Fuhrman , JA . 1994 . Bacterial viruses in coastal seawater: lytic rather than lysogenic production . Mar. Ecol. Prog. Ser , 114 : 35 – 45 .
- Wilhelm , SW , Carberry , MJ , Eldridge , ML , Poorvin , L , Saxton , MA and Doblin , MA . 2006 . Marine and freshwater cyanophages in a Laurentian Great Lake: Evidence from infectivity assays and molecular analyses of g20 genes . Appl. Environ. Microbiol , 72 : 4957 – 4963 .
- Wilhelm , SW and Suttle , CA . 1999 . Viruses and nutrient cycles in the sea: viruses play critical roles in the structure and function of aquatic food webs . BioScience , 49 : 781 – 788 .
- Wilhelm , SW , Weinbauer , MG , Suttle , CA and Jeffrey , WH . 1998 . The role of sunlight in the removal and repair of viruses in the sea . Limnol. Oceanogr , 43 : 586 – 592 .
- Williams , ST , Mortimer , AM and Manchester , L . 1987 . “ Ecology of soil bacteriophages ” . In Phage Ecology , Edited by: Goyal , SM , Gerba , CP and Bitton , G . 157 – 179 . New York : John Wiley and Sons .
- Williamson , KE , Radosevich , M and Wommack , KE . 2005 . Abundance and diversity of viruses in six Delaware soils . Appl. Environ. Microbiol , 71 : 3119 – 3125 .
- Williamson , KE , Wommack , KE and Radosevich , M . 2003 . Sampling natural viral communities from soil for culture-independent analyses . Appl. Environ. Microbiol , 69 : 6628 – 6633 .
- Williamson , SJ , Houchin , L , McDaniel , L and Paul , JH . 2002 . Seasonal variation in lysogeny as depicted by prophage induction in Tampa Bay, Florida . Appl. Environ. Microbiol , 68 : 4307 – 4314 .
- Wommack , KE and Colwell , RR . 2000 . Virioplankton: viruses in aquatic ecosystems . Microbiol. Mol. Biol. R , 64 : 69 – 114 .
- Wommack , KE , Hill , RT , Kessel , M , Russek-Cohen , E and Colwell , RR . 1992 . Distribution of viruses in the Chesapeake Bay . Appl. Environ. Microbiol , 58 : 2965 – 2970 .
- Wommack , KE , Hill , RT , Muller , TA and Colwell , RR . 1996 . Effects of sunlight on bacteriophage viability and structure . Appl. Environ. Microbiol , 62 : 1336 – 1341 .
- Wommack , KE , Ravel , J , Hill , RT , Chun , J and Colwell , RR . 1999a . Population dynamics of Chesapeake Bay virioplankton: total-community analysis by pulsed-field gel electrophoresis . Appl. Environ. Microbiol , 65 : 231 – 240 .
- Wommack , KE , Ravel , J , Hill , RT and Colwell , RR . 1999b . Hybridization analysis of Chesapeake Bay virioplankton . Appl. Environ. Microbiol , 65 : 241 – 250 .
- Yates , MV , Gerba , CP and Kelley , LE . 1985 . Virus persistence in groundwater . Appl. Environ. Microbiol , 49 : 778 – 781 .
- Yeager , JG and O’Brien , RT . 1979a . Enterovirus inactivation in soil . Appl. Environ. Microbiol , 38 : 694 – 701 .
- Yeager , JG and O’Brien , RT . 1979b . Structural changes associated with poliovirus inactivation in soil . Appl. Environ. Microbiol , 38 : 702 – 709 .
- Yin , X and Stotzky , G . 1997 . Gene transfer among bacteria in natural environments . Adv. Appl. Microbiol , 45 : 153 – 212 .
- Yoshii , H , Yoshida , T and Matsui , C . 1953 . Isolation of a Xanthomonas oryzae phage . Ann. Phytopath. Soc. Jap , 17 : 177
- Yoshimura , S , Saito , M , Yoshino , R and Morihashi , T . 1960 . Properties of a new Xanthomonas oryzae phage OP2 . Proc. Plant Protect. Hokuriku , 8 : 15 – 20 . (in Japanese)
- Zachary , A . 1976 . Physiology and ecology of bacteriophages of the marine bacterium Beneckea natriegens: salinity . Appl. Environ. Microbiol , 31 : 415 – 422 .
- Zachary , A . 1978 . An ecological study of bacteriophages of Vibrio natriegens . Can. J. Microbiol , 24 : 321 – 324 .
- Zeidner , G and Béjà , O . 2004 . The use of DGGE analyses to explore eastern Mediterranean and Red Sea marine pichphytoplankton assemblages . Environ. Microbiol , 6 : 528 – 534 .
- Zeidner , G , Bielawski , JP , Shmoish , M , Scanlan , DJ , Sabehi , G and Béjà , O . 2005 . Potential photosynthesis gene recombination between Prochlorococcus and Synechococcus via viral intermediates . Environ. Microbiol , 7 : 1505 – 1513 .
- Zeidner , G , Preston , CM Delong , EF . 2003 . Molecular diversity among marine picophytoploankton as revealed by psbA analyses . Environ. Microbiol , 5 : 212 – 216 .
- Zeph , LR , Onaga , MA and Stotzky , G . 1988 . Transduction of Escherichia coli by bacteriophage P1 in soil . Appl. Environ. Microbiol , 54 : 1731 – 1737 .
- Zeph , LR and Stotzky , G . 1989 . Use of a biotinylated DNA probe to detect bacteria transduced by bacteriophage P1 in soil . Appl. Environ. Microbiol , 55 : 661 – 665 .
- Zhong , Y , Chen , F , Wilhelm , SW , Poorvin , L and Hodson , RE . 2002 . Phylogenetic diversity of marine cyanophage isolates and natural virus communities as revealed by sequences of viral capsid assembly protein gene g20 . Appl. Environ. Microbiol , 68 : 1576 – 1584 .
- Zhuang , J and Jin , Y . 2003a . Virus retention and transport as influenced by different forms of soil organic matter . J. Environ. Qual , 32 : 816 – 823 .
- Zhuang , J and Jin , Y . 2003b . Virus retention and transport through Al-oxide coated sand columns: effects of ionic strength and composition . J. Contam. Hydrol , 60 : 193 – 209 .
- Zillig , W , Reiter , WD , Palm , P , Gropp , F , Neumann , H and Rettenberger , M . 1988 . “ Viruses of archaebacteria ” . In The Bacteriophages , Edited by: Calendar , R . Vol. 1. , 517 – 558 . New York : Plenum Press .
- Present address: Max-Planck-Institute for Terrestrial Microbiology, Karl-von-Frisch-Straße, 35043 Marburg, Germany