Abstract
Thermophilic ammonia-oxidizing bacteria (AOB) have the potential to reduce ammonium emissions from commercial-scale animal waste composting operations. Two new media have been designed to culture and enumerate thermophilic AOB species. C-1 medium was designed to reflect compost composition. C-2 medium and CQ-2 medium were modified synthetic organic wastewater for the cultivation and enumeration of AOB, respectively. These new media have several advantages over the conventional medium. C-2 medium is effective for nitrification at 50°C, and C-1 and C-2 media support reasonable growth of thermophilic AOB. Furthermore, C-1 and CQ-2 media allow enumeration of thermophilic AOB during the composting process. This is the first report of the successful cultivation and enumeration of thermophilic AOB in compost. These results suggest the possibility of isolation and manipulation of novel thermophilic AOB species for environmental bioremediation.
INTRODUCTION
Composting animal waste emits ammonia and sulfur compounds. Ammonia emission in particular produces an unpleasant odor and is a source of irritation for neighbors of large-scale compost operations. Ammonia-oxidizing bacteria (AOB) oxidize ammonia and reduce the smell of ammonia (CitationYomura et al. 2002), making them ideal potential organisms for animal waste bioremediation. Thus, recent research has focused on basic AOB biology.
Thermophilic AOB isolated from compost are needed for biological deodorization of ammonia in compost. Nitrosomonas sp. and Nitrosospira sp. have been reported to be deodorizing AOB (CitationAbd El Haleem et al. 2000; CitationSatoh et al. 2004). Ammonia-oxidizing bacteria such as Nitrosomonas communis, Nitrosomonas europaea, Nitrosomonas nitrosa (CitationPurkhold et al. 2000) and Nitrosospira (“Nitrosolobus”) (CitationSatoh et al. 2007) have been isolated from activated sludge and soil. These AOB were autotrophic and mesophilic (CitationProsser 1989; CitationWatson et al. 1989), and were amenable to growth on defined mineral media at 20–37°C (CitationMatulewich et al. 1975; CitationSuwa et al. 1997). Of these autotrophic AOB, N. europaea was found to grow on organic matter (CitationHommes et al. 2003), and some heterotrophic bacteria have been shown to have nitrification activity (CitationPapen et al. 1989; CitationVerstraete and Alexander 1972). Accordingly, media containing organic matter have come to be used in autotrophic and heterotrophic AOB research (CitationJoo et al. 2005; CitationLaanbroek and Bär-Gilissen 2002; CitationTomiyama et al. 2001).
In contrast to mesophilic AOB species, thermophilic AOB were investigated from hot springs and deep-sea hydrothermal vents (CitationLebedeva et al. 2005; CitationMével and Prieur 2000), but thermophilic AOB have not been isolated from compost (CitationPurkhold et al. 2003). New media that contain organic matter and that can be incubated at high temperature are indispensable culture conditions for thermophilic AOB from compost. The new media containing organic matter must be designed to approximate the composition of the complex composting environment. Furthermore, thermophilic AOB in compost must be cultured at temperatures of more than 50°C, which are generated by the composting process (CitationShimaya et al. 2003). In this study we generated improved new media for the culture and enumeration of thermophilic AOB in compost and we investigated the multiplication of AOB during the composting process.
MATERIALS AND METHODS
Analysis of nitrification
Ammonium nitrogen (NH4-N), nitrite nitrogen (NO2-N) and nitrate nitrogen (NO3-N) resulting from nitrification were measured, and the activity of AOB was compared. Dairy cattle manure and sawdust composted for 3 months at the thermophilic stage were used as sources of AOB. Five grams of compost was suspended in 45 mL of sterile distilled water for 30 min using a SHAKER MODEL SA-51 (Yamato Scientific Company, Tokyo, Japan). The compost suspension (10 mL) was added to a flask containing 150 mL of one of the three media () and the flasks were incubated at 50°C with shaking (200 rpm) for 8 weeks. Uninoculated flasks served as negative controls. The fluid volume was adjusted with sterilized water to maintain a constant solute concentration. Cultures were sampled once per week over the course of the experiment. Samples were filtered to remove bacteria (0.45-µm pore size). Concentrations of NH4-N, NO2-N and NO3-N were measured with an AACSII (Bran+Luebbe, Norderstedt, Germany). Experiments were carried out in triplicate and the results are presented as means and standard deviations.
Enumeration of thermophilic AOB
The AOB were enumerated using the most probable number calculation (CitationHalvorson and Ziegler 1932) using all three media for 8 weeks. Aliquots (0.1 mL) of serial 10-fold sterile distilled water dilutions of the culture supernatant were transferred to 96-well microtiter plates containing each medium. Inoculated microtiter plates were then placed on a wet towel, sealed in boxes to avoid evaporation, and incubated for 4 weeks at 50°C. The AOB activity was assayed using NO− 2 production with Griess–Ilosvay reagents. When necessary, zinc powder was added to test for NO− 3 as a precaution against false negatives (CitationKimura 1992).
Composting test
Composting was carried out in a composting tank (1.8 m3; Kumamoto Sanso, Kumamoto, Japan) with dairy cattle manure. Compost was sampled once per week until the fourth week when the compost maintained a thermophilic state (50–70°C). Composting samples (5 g) were suspended in 45 mL of sterile distilled water for 30 min using a SHAKER MODEL SA-51 (Yamato Scientific Company). Thermophilic AOB were assayed using the most probable number (MPN) method for all three media at 50°C.
RESULTS AND DISCUSSION
Design of improved media for thermophilic AOB in compost
The conventional AOB medium contains a concentration of ammonia (680 mg N L−1) and does not include a carbon source because the medium is used only for autotrophs (CitationKimura 1992). However, thermophilic AOB present in compost may include both autotrophic and heterotrophic species, or species that are capable of both metabolic strategies. Thus, a carbon source can be used as a selective agent by simulating the organic matter and high ammonia concentration of compost, and to encourage more rapid growth of the selected organisms. Conventional media are likely to be insufficient for culturing and isolating composting AOB because these organisms have variable ammonia oxidation activities because of their sensitivity to the C/N ratio (CitationSatoh et al. 1999).
Table 1 Composition of the ammonia-oxidizing bacteria media (L−1)
The C-1 medium was designed for cultivation of AOB and was modified to simulate the composition of compost. Compost is 54.8% water and has a C/N ratio of 18.9, pH of 8.4, electrical conductivity of 4.7 mS cm−1, 1.9% total N, 35.3% total C, 2.3% P2O5, 2.4% K2O, 3.0% CaO and 1.0% MgO (CitationHaga 2001). Thus, the C-1 medium was made with the same N:P:K:Mg ratio and pH as compost, and because the C/N ratio of compost is 18.9, the medium was amended with 756.9 mmol L−1 of CH3COONa·3H2O. Sodium acetate was selected for the isolation of thermophilic ammonium-tolerant bacteria from compost (CitationKuroda et al. 2004). In preliminary experiments, ammonia concentrations of 1,000, 2,000 and 3,000 mg N L−1 were tested to determine the effect of ammonia concentration on nitrification activity. The nitrification activity was highest at 1,000 mg N L−1 (CitationShimaya et al. 2003). Therefore, 1,360 mg N L−1 (NH4)2SO4 (twice the concentration of the conventional medium) was added (; C-1). The C-1 medium was also used for enumeration without further modification.
The C-2 medium was designed for the cultivation of AOB and is essentially a modified synthetic organic wastewater (CitationFurukawa et al. 1994), which contains a high concentration of organic matter. The total carbon of wastewater is 38.57%, which is similar to compost. Peptone and meat extract were used as the carbon, nitrogen and phosphorus sources (CitationFurukawa et al. 1994), and for the same reason as for C-1 medium, twice the (NH4)2SO4 of the conventional medium was added (; C-2). The C-2 medium is deeply colored by peptone and meat extract, making it difficult to assay color development using the MPN method. Therefore, for enumeration CQ-2 medium was modified to contain only 0.6 g L−1 peptone and no meat extract. As NaHCO3 foams with the addition of Griess–Ilosvay reagents (CitationKimura 1992), this salt was reduced to 89.3 mmol L−1 from 499.9 mmol L−1 (; CQ-2).
Comparison of nitrification in the C-1 and C-2 media
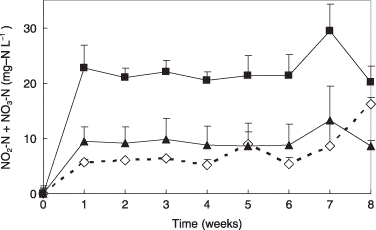
Ammonium nitrogen steadily decreased over 8 weeks in all three media. Ammonium nitrogen was undetectable in uninoculated controls by the end of the incubation period. Therefore, the decrease in NH4-N depends on volatilization and oxidization. In contrast, NO2-N and NO3-N increased 9 ± 3 mg N L−1 within 1 week in the C-1 medium. Nitrite nitrogen and nitrate nitrogen increased 23 ± 4 mg N L−1 within 1 week (P < 0.01) in the C-2 medium. In the C-1 and C-2 media, only NO2-N accumulated within 1 week. More than half of the accumulated NO2-N was converted to NO3-N by 8 weeks. Nitrite nitrogen and nitrate nitrogen reached only 6 ± 0 mg N L−1 within 1 week in the conventional medium (). Significantly less NO2-N accumulated in the conventional medium. No NO2-N or NO3-N was generated in the uninoculated controls. The above results suggest that C-2 medium is superior to the conventional medium for nitrification activity.
Figure 1 Accumulation of NO2-N and NO3-N during incubation at 50°C. (⋄) Conventional medium; (▴) C-1 medium; (▀) C-2 medium. Data are the mean values of three replicates. Error bars indicate standard deviation.
Enumeration of thermophilic AOB using C-1 and CQ-2 media on cultures in C-1 and C-2 media
Thermophilic AOB did not increase in the conventional medium over the course of the 8-week experiment. However, thermophilic AOB entered a logarithmic growth phase after the second week in the C-1 and C-2 media, and had increased by an order of magnitude after 6 weeks (P < 0.01) (). Thermophilic AOB increased after the second week, but growth was not accompanied by an increase in NO2-N and NO3-N (). It was postulated that the multiplied AOB formed granules and the internal AOB cells in the granules had weak oxidization activity. Thus, viable counts were not consistent with the activity (CitationNishio and Furusaka 1971). The C-1 and C-2 media are, thus, superior media for culturing thermophilic AOB from compost (,). Both autotrophic and heterotrophic bacteria may be supported by these media because autotrophic N. europaea and Nitrobacter winogradskyi can survive in the presence of organic matter (CitationFrank et al. 1991). It will be necessary to investigate the growth of autotrophic and heterotrophic AOB in each medium.
Detection of thermophilic AOB for the composting process using C-1 and CQ-2 media
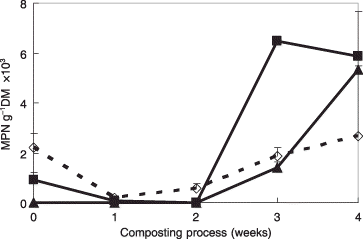
Thermophilic AOB in the composting process were rarely detected within 2 weeks using C-1, CQ-2 or the conventional medium at 50°C. In 3 weeks, however, there were approximately threefold as many thermophilic AOB detected with CQ-2 medium compared with the conventional medium (P < 0.01). Furthermore, after 4 weeks, there were twice as many thermophilic AOB using C-1 and CQ-2 media than using the conventional medium, with 5.4 × 103 MPN g−1 dry matter in C-1 medium, 5.9 × 103 MPN g−1 dry matter in CQ-2 medium and 2.7 × 103 MPN g−1 dry matter in the conventional medium (). The differences in the numbers of thermophilic AOB in each type of medium were difficult to clarify because of the large sample variation in the 4-week-old cultures.
Figure 2 Enumeration of thermophilic ammonia-oxidizing bacteria using C-1 and CQ-2 media on cultures in C-1 and C-2 media. (⋄) Conventional medium; (▴) C-1 medium; (▀) CQ-2 medium. Data are the mean values of three replicates. Error bars indicate standard deviation. MPN, most probable number.
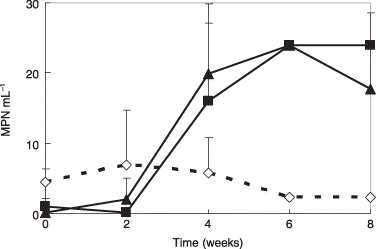
Figure 3 Detection of thermophilic ammonia-oxidizing bacteria using C-1 and CQ-2 media in the composting process. (⋄) Conventional medium; (▴) C-1 medium; (▀) CQ-2 medium. Data are the mean values of three replicates. Error bars indicate standard deviation. dry matter (DM); MPN, most probable number.
Effectiveness of improved media for thermophilic AOB in compost
The C-1 medium was modified to reflect compost composition and the C-2 medium was a modified synthetic organic wastewater. The C-1 and C-2 media were specifically modified for culturing thermophilic AOB from compost by including 1360 mg N L−1 (NH4)2SO4 and organic matter. Amendment of the C-1 medium with sodium acetate, and the C-2 medium with peptone and meat extract were effective for culturing AOB from compost.
The C-1 and C-2 media have several advantages over the conventional medium used for culturing thermophilic AOB from compost. The C-2 medium allows effective nitrification at 50°C (), the C-1 and C-2 media support reasonable growth of thermophilic AOB (), and the C-1 and CQ-2 media allow quantification of thermophilic AOB during the composting process (). Therefore, it is likely that novel thermophilic AOB will be isolated from compost in these media, which may then have commercial application for the bioremediation of ammonia released from animal waste.
Nitrification activity was higher in the C-2 medium than in the C-1 medium (). Therefore, the C-2 medium seems to be superior to the C-1 medium. Further research will be necessary to analyze all cultured bacteria in these media. Isolation and identification of thermophilic AOB would be worthwhile to confirm the superiority of the C-2 medium.
In conclusion, this is the first report of the successful cultivation and enumeration of thermophilic AOB from active, high-temperature compost using media designed specifically for this purpose. These media have the potential to be further modified as solid agar media for single colony isolation, which would provide a means for the isolation and identification of novel AOB species.
ACKNOWLEDGMENTS
The authors would like to express their appreciation to Professor Kyung-Sook Whang, Mokwon University, for valuable suggestions and to Dr Akihiro Tanaka, National Agricultural Research Center for Kyusyu Okinawa Region, for his support in collecting compost samples.
Related Research Data
REFERENCES
- Abd El Haleem , D , von Wintzingerode , F , Moter , A , Moawad , H and Göbel , UB . 2000 . Phylogenetic analysis of rhizosphere-associated β-subclass proteobacterial ammonia oxidizers in a municipal wastewater treatment plant based on rhizoremediation technology . Lett. Appl. Microbiol , 31 : 34 – 38 .
- Frank , J , Verhagen , M and Laanbroek , HJ . 1991 . Competition for ammonium between nitrifying and heterotrophic bacteria in dual energy-limited chemostats . Appl. Environm Microbiol , 57 : 3255 – 3263 .
- Furukawa , K , Tanaka , N and Fujita , M . 1994 . Stoichiometry of activated sludge process for the treatment of synthetic organic wastewater . J. Ferment. Bioeng , 78 : 175 – 178 .
- Haga , K . 2001 . Adviser Stock Raising Environment Training Workshop , Tokyo : Livestock Industry's Environmental Improvement Organization . (in Japanese)
- Halvorson , HO and Ziegler , NR . 1932 . Application of statistics to problems in bacteriology: I. A means of determining bacterial population by the dilution method . J. Bacteriol , 25 : 101 – 121 .
- Hommes , NG , Sayavedra-Soto , LA and Arp , DJ . 2003 . Chemolithoorganotrophic growth of Nitrosomonas europaea on fructose . J. Bacteriol , 185 : 6809 – 6814 .
- Joo , HS , Hirai , M and Shoda , M . 2005 . Characteristics of ammonium removal by heterotrophic nitrification-aerobic denitrification by Alcaligenes faecalis No. 4 . J. Biosci. Bioeng , 100 : 184 – 191 .
- Kimura , M and Study group of soil microbe . 1992 . “ Measurement of nitrification, calculation and separation of nitro bacteria ” . In A New Soil Microbe Experimental Method. , 207 – 214 . Tokyo : Yokendo . (in Japanese)
- Kuroda , K , Hanajima , D Fukumoto , Y . 2004 . Isolation of thermophilic ammonium-tolerant bacterium and its application to reduce ammonia emission during composting of animal wastes . Biosci. Biotechnol. Biochem , 68 : 286 – 292 .
- Laanbroek , HJ and Bär-Gilissen , MJ . 2002 . Weakened activity of starved ammonia-oxidizing bacteria by the presence of pre-activated Nitrobacter winogradskyi. . Microbes Environ , 17 : 122 – 127 .
- Lebedeva , EV , Alawi , M , Fiencke , C , Namsaraev , B , Bock , E and Spieck , E . 2005 . Moderately thermophilic nitrifying bacteria from a hot spring of the Baikal rift zone . FEMS Microbiol. Ecol , 54 : 297 – 306 .
- Matulewich , VA , Strom , PF and Finstein , MS . 1975 . Length of incubation for enumerating nitrifying bacteria present in various environments . Appl. Environ. Microbiol , 29 : 265 – 268 .
- Mével , G and Prieur , D . 2000 . Heterotrophic nitrification by a thermophilic Bacillus species as influenced by different culture conditions . Can. J. Microbiol , 46 : 465 – 473 .
- Nishio , M and Furusaka , C . 1971 . Kinetic study of soil percolated with nitrite . Soil Sci. Plant Nutr , 17 : 61 – 67 .
- Papen , H , Berg , RV , Hinkel , I , Thoene , B and Rennenberg , H . 1989 . Heterotrophic nitrification by Alcaligenes faecalis: NO2–, NO3–, N2O, and NO production in exponentially growing cultures . Appl. Environm. Microbiol , 55 : 2068 – 2072 .
- Prosser , JI . 1989 . Autotrophic nitrification in bacteria . Adv. Microb. Physiol , 30 : 125 – 181 .
- Purkhold , U , Pommerening-Röser , A , Juretschko , S , Schmid , MC , Koops , HP and Wagner , M . 2000 . Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amoA sequence analysis: implications for molecular diversity surveys . Appl. Environm. Microbiol , 66 : 5368 – 5382 .
- Purkhold , U , Wagner , M , Timmermann , G , Pommerening-Röser , A and Koops , HP . 2003 . 16S rRNA and amoA-based phylogeny of 12 novel betaproteobacterial ammonia-oxidizing isolates: extension of the dataset and proposal of a new lineage within the nitrosomonads . Int. J. Syst. Evol. Microbiol , 53 : 1485 – 1494 .
- Satoh , H , Okabe , S and Watanabe , Y . 1999 . Significance of substrate C/N ratio on structure and activity of nitrifying biofilms determined by in situhybridization and the use of microelectrodes . Water Res , 22 : 763 – 769 . (in Japanese)
- Satoh , K , Itoh , C Kang , DJ . 2007 . Characteristics of newly isolated ammonia-oxidizing bacteria from acid sulfate soil and the rhizoplane of leucaena grown in that soil . Soil Sci. Plant Nutr , 53 : 23 – 31 .
- Satoh , K , Takizawa , R , Sarai , M , Sato , N , Takahashi , R and Tokuyama , T . 2004 . Two kinds of ammonia-oxidizing bacteria isolated from biologically deodorizing plants in cold district . J. Biosci. Bioeng , 98 : 207 – 210 .
- Shimaya , C , Tanaka , A , Hashimoto , T and Yakushidou , K . 2003 . Activity of thermophilic bacteria in compost . Kyusyu Agric. Res , 65 : 82 (in Japanese)
- Suwa , Y , Sumino , T and Noto , K . 1997 . Phylogenetic relationships of activated sludge isolates of ammonia oxidizers with different sensitivities to ammonium sulfate . J. Gen. Appl. Microbiol , 43 : 373 – 379 .
- Tomiyama , H , Ohshima , M Ishii , S . 2001 . Characteristics of newly isolated nitrifying bacteria from rhizoplane of paddy rice . Microbes Environ , 16 : 101 – 108 .
- Verstraete , W and Alexander , M . 1972 . Heterotrophic nitrification by Arthrobacter sp . J. Bacteriol , 110 : 955 – 961 .
- Watson , SW , Bock , E , Harms , H , Koops , HP and Hooper , AB . 1989 . Nitrifying Bacteria. Bergey's Manual of Systematic Bacteriology , Edited by: Staley , JT , Bryant , MP , Pfennig , N and Holt , JG . vol. 3 , 1808 – 1834 . Baltimore : The Williams and Wilkins Company .
- Yomura , Y , Oyamada , K , Noda , K , Tsuchiya , H and Miyazawa , K . 2002 . Development of a new biological deodorizing system of ammonium gas . NKK Report , 178 : 81 – 86 . (in Japanese)