Abstract
Soil acidity is one of the most serious causes of land degradation and limits crop production in subtropical agriculture. A number of legumes have adapted to P-limiting conditions, and the aim of the present study was to select an appropriate species for use as a winter legume cover crop under subtropical soil conditions in Okinawa, where P deficiency stems from soil acidification. Four vetch and four clover species were examined in pot trials. Of these, hairy vetch (Vicia villosa Roth) was found to be the most tolerant of P-limiting conditions. The shoot biomass of hairy vetch was also generally higher than that in the clover species when the P supply was non-limiting in the spring season. Hairy vetch was then compared in a pot trial with four clover species for an extended period of growth with adequate P supply. The N fixing activity of hairy vetch in the vegetative stage was 3.2-fold greater than that of the clover species. Nitrogen and P uptake in shoots and roots was significantly higher in hairy vetch than in the clover species in the vegetative and flowering stages. The C/N ratio in the shoots and roots of hairy vetch tended to be lower than that in the clover species. Shoot K uptake by hairy vetch was significantly higher in the vegetative stage than that in the clover species. In a field trial, shoot biomass and nutrient uptake of hairy vetch were similar on four different subtropical soils with pH values ranging from 5.7 to 7.5. Hairy vetch is an appropriate winter legume cover crop to improve subtropical soils by increasing organic matter and supplying nutrients to the subsequent crop.
Introduction
Soil acidification is one of the most serious forms of land degradation in subtropical agriculture. Soils that are acidic or are acidifying generally exist where rainfall exceeds 450 mm year−1 (CitationScott et al. 2000), potentially limiting crop production on the most productive agricultural land. The harvesting of high-yielding crops also depletes organic matter from agricultural fields, a process that both contributes to and is accelerated by soil erosion when the soil is left bare after crop harvests (CitationBayer et al. 2001; CitationOyedele and Aina 1998; CitationSanchez and Logan 1992; CitationTiessen et al. 1994).
Phosphorus is often a limiting nutrient in acidic soils owing to the high P-fixing capacity of these soils. Most of the P in mineral fertilizers reacts with Fe and Al compounds and is transformed into relatively insoluble forms. Plants that are tolerant of low P availability are often used to maintain plant productivity on acid soils. Some legumes can exude organic compounds that mobilize P from bound P pools in the soil (CitationLiao et al. 2006).
Winter legume cover crops are an important biological tool for sustainable agriculture because they can increase the inputs of organic matter to the soil, leading to improved soil aggregate stability and therefore reduced run-off and erosion. Maximizing organic residues through the use of winter legume cover crops is an important strategy for improving soil quality in agricultural practices (CitationBayer et al. 2000; CitationZanatta et al. 2007). Moreover, winter legume cover crops fix atmospheric N and suppress weeds. However, considerable variability in growth productivity among winter legume cover crops, which include species of vetch and clover, means that species selection is an important consideration in developing low-input cropping systems (CitationDen Hollander et al. 2007).
The effect of hairy vetch on maize production in a relay cropping system in subtropical regions has been investigated by CitationZougmore et al. (2006). However, the performance of hairy vetch as a cover crop compared with that of other vetch and clover species under subtropical conditions is not well understood.
The objectives of the present study were to: (1) select a species of vetch or clover that is tolerant of P deficiency over a short growing period, (2) confirm the growth of hairy vetch and four clover species over a long growing period when P is not limiting, (3) examine the field performance of hairy vetch in four different soils in subtropical Okinawa by determining biomass and nutrient uptake.
Materials and methods
Pot trials with clover and vetch species
We conducted two sets of experiments under controlled conditions to evaluate the performance of four species of clover and four species of vetch. The species examined were perennial white clover (Trifolium repens L.), biennial red clover (Trifolium pretense L.), annual crimson clover (Trifolium incarnatum L.), annual pink clover (Trifolium gracilentum L.), annual hairy vetch (Vicia villosa Roth), annual tiny vetch (Vicia hirsuta L.), annual narrowleaf vetch (Vicia angustifolia L.) and annual Chinese milk vetch (Astragalus sinicus L.).
The first experiment examined all eight species under two conditions, P limiting and P non-limiting. The second experiment examined the best performing vetch (hairy vetch) from the first experiment against the four clover species with three harvests of above-ground biomass over an extended growing season with P non-limiting. The three harvests represented vegetative, flowering and mature growth stages. The experiments were conducted at the Subtropical Field Science Center, University of the Ryukyus, Okinawa, Japan (26°N, 127°E). The plants were grown in 1/5000-are Wagner pots filled with 2.2 kg Akadama soil. Individual treatments were replicated with three pots in both experiments. The chemical properties of the soil in each experiment are shown in . In both experiments, the N and K2O requirements of the plants were supplied by the addition of 0.5 g pot−1 of each as urea and K2SO4, respectively.
Table 1 Initial chemical properties of the different soils used in the three different experiments
Experiment 1: Clover and vetch species under P-limiting and P-non-limiting conditions
The P-limiting and P-non-limiting conditions were prepared by adding superphosphate to the pots to achieve P2O5 absorption coefficient values of 0.01 and 0.1, respectively (CitationKitou et al. 2009). Ten seeds of each of the four clovers were sown on 19 October 2005. In a separate set of pots, seven seeds of each of the vetch species were sown on 2 November 2005. Clover and vetch species were harvested on 14 December and 28 December 2005, respectively.
Experiment 2: Selection of hairy vetch and clover species under P-non-limiting conditions
In a previous study we compared the growth of narrowleaf vetch, tiny vetch and hairy vetch over a longer growing period (3–5 months after sowing) under P-non-limiting conditions (CitationAnugroho and Kitou 2007). In the present study, we repeated this experiment with hairy vetch and the four clover species. P2O5 was supplied at the same rate as in the P-non-limiting condition of Experiment 1. Seeds were sown at a rate of 50 mg pot−1 on 6 November 2006. All plants were harvested on 6 February, 6 March and 6 April 2007.
Experiment 3: Field performance of hairy vetch on four different soils
Field experiments were carried out at the Tropical Agriculture Research Front of the Japan International Research Center for Agricultural Science, Ishigaki, Okinawa, Japan (24°N, 129°E). Four different soils (yellow, red, dark red and gray soil) in plots of 2 m × 2.4 m at 2 m depth were used. The chemical properties of the four soils are shown in . Hairy vetch was sown on 24 November 2006, with four seeds per hill and 16 hills m−2. No fertilizer was applied to the soils. Hairy vetch was harvested on 24 April 2007 by sampling all above-ground biomass within 0.5 m × 0.5 m randomly placed quadrats per plot. Each treatment had three replicates.
Plant and soil analyses
The N-fixing activity was measured by the acetylene reduction assay method (CitationYoshida 1982) using gas chromatography (GS-2; Sensortec, Tokyo, Japan). Shoots and roots were dried at 70°C for 72 h, weighed and ground to a powder. The total C and N contents in the plant samples were determined using a CHN Corder (JM10; G-Science Laboratory Company, Tokyo, Japan). The total P in the plant material was determined by calorimetry after combustion at 450°C for 1 h, and the resulting ash was dissolved in 1:30 (v/v) diluted nitric acid (CitationHafner et al. 1993). The K, Ca and Mg concentrations were determined by an atomic absorption spectrophotometer (Solaar 969; Japan Thermo Corporation, Tokyo, Japan) after Kjeldahl digestion with H2SO4 and H2O2.
Soils were air-dried and passed through a 2-mm sieve. Soil pH and electrical conductivity (EC) were measured in 1:2.5 and 1:5 (w/v) soil : water slurries, respectively. The total C and N contents in the soils were determined using a CHN Corder (JM10; G-Science Laboratory Company). The ammonium-N and nitrate-N contents were determined by steam distillation after extraction with 2 mol L−1 KCl (CitationMulvaney 1996). The phosphate absorption coefficient was determined calorimetrically after absorption with 25 g L−1 (NH4)2HPO4. Calcium-bound phosphate (as a percentage of the available phosphate) was determined calorimetrically after extraction with 25 mL L−1 acetic acid. Exchangeable cations were determined using a Solaar 969 atomic absorption spectrophotometer after extraction with 1 mol L−1 ammonium acetate.
Statistical analysis
Data are presented as the means of three replicates and the standard errors of the means of the sets of values were statistically compared. Biomass and nutrient uptake data were analyzed using a one-way anova with a complete randomized block design and Fisher’s least significant difference test (P < 0.05) were used for post-hoc comparisons of the means of significant anova results. The statistical procedures were carried out in the software package SSPS 14.0 for Windows (SPSS, Chicago, IL, USA).
Results
Growth of plants under P-limiting and P-non-limiting conditions
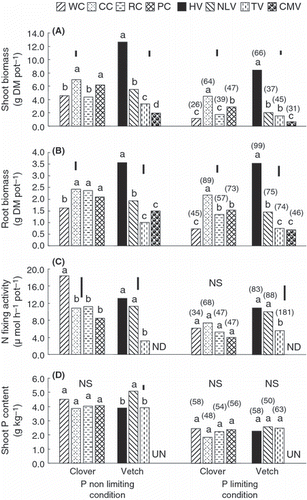
Under P-non-limiting conditions, crimson clover and pink clover produced significantly greater shoot biomass than white clover and red clover (), and crimson clover, red clover and pink clover produced significantly greater root biomass than white clover (). Under P-limiting conditions, the shoot and root biomasses of crimson clover were significantly greater than those of the other clover species. The shoot and root biomasses of hairy vetch were significantly greater than those of the other vetch species under both P-limiting and P-non-limiting conditions. The biomass of the shoots and roots of crimson clover were 64 and 89%, respectively, of those under P-non-limiting condition; these values were higher than the corresponding percentages of the other clover species. Similarly, the biomass of the shoots and roots of hairy vetch under P-limiting conditions were 66 and 99%, respectively, of those under P-non-limiting conditions, and these values were higher than those of the other vetch species. These results show that crimson clover and hairy vetch are tolerant of P-limiting conditions.
Nitrogen-fixing activity under P-non-limiting conditions was significantly higher in white clover than in the other clover species; however, there were no significant differences between clover species under P-limiting conditions (). The N-fixing activities of hairy vetch and narrowleaf vetch were significantly higher than that of tiny vetch under both P-non-limiting and P-limiting conditions. The N-fixing activities of crimson clover and hairy vetch under P-limiting conditions were 68 and 83% of the respective N-fixing activities under P-non-limiting conditions. Narrowleaf vetch and tiny vetch recorded N-fixing activities under P-limiting conditions that were 88 and 181%, respectively, of what they produced under P-non-limiting conditions; however, the growth of both plants was less than that of hairy vetch. Nitrogen-fixing activity was not detected in Chinese milk vetch. Shoot P contents were not significantly different among the clover species under both P-non-limiting and P-limiting conditions (). The shoot P content of narrowleaf vetch was significantly higher than that of hairy vetch and tiny vetch under P-non-limiting conditions, but there were no significant differences between the other vetch species. Clover and vetch species had similar shoot P contents under P-limiting conditions, and these percentages were 48–63 of the comparative P contents under P-non-limiting conditions.
Figure 1 (A) Shoot biomass and (B) root biomass, (C) N-fixing activity and (D) shoot P content of clover and vetch species grown at two different P levels (P-non-limiting and P-limiting) in Experiment 1. WC, white clover; CC, crimson clover; RC, red clover; PC, pink clover; HV, hairy vetch; NLV, narrow leaf vetch; TV, tiny vetch; CMV, Chinese milk vetch; DM, dry matter; ND, not detected; UN, unmeasured. Numbers in parentheses indicate the percentages of the P-limiting to P-non-limiting ratios. Vertical bars represent significant differences according to a least significant difference test (n = 3); columns with the same letter are not significantly different (NS) at P < 0.05.
Growth of plants under P-non-limiting conditions over a longer growing period
The biomass of the shoots and roots of plants increased with each successive harvest, that is, in February, March and April (). Hairy vetch produced greater shoot biomass and root biomass than the clover species at all harvests, except that the shoot biomass of pink clover and the root biomass of red clover and pink clover were greater (but not significantly so) than those of hairy vetch (). Moreover, the shoot biomass of pink clover was significantly greater than that of the other clover species harvested in March and April. The N-fixing activity of hairy vetch was significantly greater than that of the clover species in February and March, except for white clover in March (); in April, however, hairy vetch recorded the lowest N-fixing activity. In February, the N-fixing activity of hairy vetch was 61 μmol h−1 pot−1, double the levels of the clover species.
Figure 2 (A) Shoot biomass, (B) root biomass and (C) N-fixing activity of hairy vetch and clover species harvested from February to April in Experiment 2. HV, hairy vetch; WC, white clover; CC, crimson clover; RC, red clover; PC, pink clover; DM, dry matter. Vertical bars represent significant differences according to a least significant difference test (n = 3); columns with the same letter are not significantly different (NS) at P < 0.05.
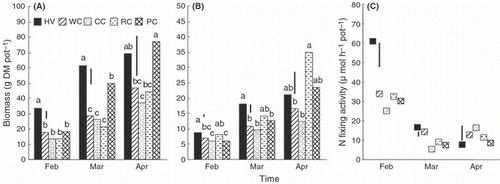
Figure 3 (A) Nitrogen uptake and (B) C/N ratio of shoot and root parts of hairy vetch and clover species harvested from February to April in Experiment 2. HV, hairy vetch; WC, white clover; CC, crimson clover; RC, red clover; PC, pink clover. Vertical bars represent significant differences according to a least significant difference test (n = 3); columns with the same letter are not significantly different (NS) at P < 0.05.
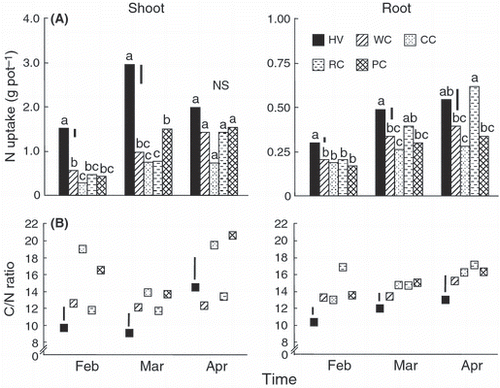
Nitrogen uptake in shoots and roots in all plants followed similar increases to those observed for biomass, except that shoot N uptake did not increase in the April harvest in hairy vetch and crimson clover (). Hairy vetch harvested in February and March had significantly greater shoot N uptake than the clover species. The roots of red clover harvested in April had the highest N uptake of all plants. The C/N ratios varied over ranges of 11.7–20.7 in the shoots and 13.0–17.1 in the roots of clover species; the corresponding ranges for hairy vetch were 9.1–14.5 and 10.3–13.0 (). The C/N ratios of shoots and roots of hairy vetch were generally significantly lower than those of clover species in February, March and April, but the shoot C/N ratio of hairy vetch in April was similar to that of white clover and red clover.
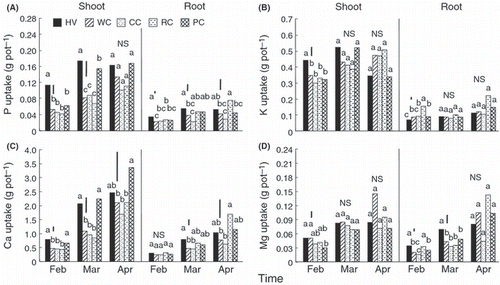
Shoot P uptake also tended to follow shoot biomass, although in hairy vetch P uptake did not increase from the March to April harvests (). Root P uptake increased between February and March, except in crimson clover, and increased again in April, except in hairy vetch and pink clover. Meanwhile, red clover displayed a clear increase in P uptake in the shoots and roots between March and April. In general, P uptake in the shoots and roots of hairy vetch was relatively greater than in clover species in February and March. In addition, shoot mineral uptake of plants increased during February, March and April (), except for shoot K uptake in hairy vetch and pink clover and shoot Mg uptake in crimson clover in April. Red clover displayed significantly greater root uptake of P and minerals in April than the other plants. Shoot K uptake of hairy vetch harvested in February was significantly greater than the uptake in the four clovers. Root K uptake of red clover in February was significantly greater than the uptake of the other four species. Shoot Ca uptake of hairy vetch and pink clover was significantly greater than the uptake of white clover, crimson clover and red clover in February and March, and in the case of pink clover also in April, when hairy vetch was not significantly different from any other species. Root Ca uptake of hairy vetch harvested in March was significantly greater than the uptake of white clover and crimson clover. Shoot Mg uptake of hairy vetch and white clover harvested in February was significantly higher than that of pink clover, whereas root Mg uptake of hairy vetch in February and March was significantly higher than that of all clover species, except red clover, in February. In general, there were very few significant differences between species in shoot uptake of P or minerals at the April harvest.
Figure 4 (A) Phosphorus and (B) K, (C) Ca and (D) Mg uptakes by the shoots and roots of hairy vetch and clover species harvested from February to April in Experiment 2. HV, hairy vetch; WC, white clover; CC, crimson clover; RC, red clover; PC, pink clover. Vertical bars represent significant differences according to a least significant difference test (n = 3); columns with the same letter are not significantly different (NS) at P < 0.05.
Biomass and nutrient uptake of hairy vetch in field trials on four different soils
The shoot biomass and nutrient uptake of hairy vetch were not significantly different between the four different soils (; ). Shoot biomass ranged from 400 to 500 g m−2. Nitrogen uptake by shoots was in the range of 11.5–15.4 g m−2 and P, K, Ca and Mg uptake by shoots displayed ranges of 0.9–1.7, 5.7–8.4, 3.4–6.4 and 0.7–1.2 g m−2, respectively. The P concentrations of hairy vetch grown in the more neutral pH dark red and gray soils were significantly higher than those grown in the more acidic yellow and red soils. The Ca concentration of hairy vetch grown in the neutral gray soil was significantly higher than that in the acid soils. The C/N ratios ranged from 12.8 to 14.4.
Figure 5 Biomass of the shoots of hairy vetch grown in four different soils in Experiment 3. YS, yellow soil; RS, red soil; DS, dark red soil; GS: gray soil; DM, dry matter. Columns with the same letter are not significantly different (NS) at P < 0.05.
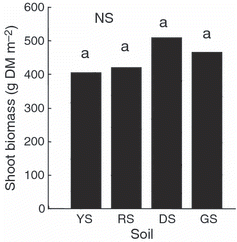
Table 2 Nutrient concentration and uptake of hairy vetch grown under four different soils in Experiment 3
Discussion
With a short growing period under P-limiting conditions, shoot and root biomass () and root P content (data not shown) of hairy vetch and crimson clover were greater than those of other vetch species and other clover species, respectively. The shoot biomass of narrowleaf vetch, tiny vetch and Chinese milk vetch with P non-limiting and a short growing period were 44, 26 and 16%, respectively, of that of hairy vetch. These results are in line with results from an earlier study in which the shoot biomass of narrowleaf vetch and tiny vetch grown under P-non-limiting conditions over a long growing period in a subtropical region ranged from 42–62% and 12–25%, respectively, of the value of hairy vetch (CitationAnugroho and Kitou 2007). Even though Chinese milk vetch is widely used as a cover fertilizer crop in rice fields in temperate regions (CitationSamarajeewa et al. 2005), it did not grow well under the subtropical conditions of the present study.
In the present study, over a longer growing period and with P non-limiting, the total biomass of shoots and roots of hairy vetch in the vegetative (February) and flowering (March) stages was almost double that of the clover species, excluding pink clover in the flowering stage; similar results have been found for hairy vetch and white clover in a northern temperate region (CitationBrandsaeter et al. 2008). The greater biomass of hairy vetch may result from its high N-fixing activity in the late vegetative and early flowering stages (CitationAnugroho et al. 2009a). Unlike hairy vetch and pink clover, the flowering stage of white clover and crimson clover occurred in April, whereas red clover was still in the vegetative stage during the experimental period.
In the field experiments, the pH of the soils ranged from 5.7 to 7.5, which is within the range of 4.9–8.2 recommended for hairy vetch in temperate regions (CitationDuke 1981). The shoot biomass production of hairy vetch in the four different soils was similar to previous results reported from subtropical and northern temperate regions (CitationAnugroho et al. 2009b; CitationBrandsaeter et al. 2008). Incorporation of hairy vetch into soil reduced N fertilizer demand to 90 kg ha−1 for tomato production in a northern temperate region (CitationSainju et al. 2002).
Although the shoot biomass of hairy vetch increased markedly from February to March and increased further in April, N-fixing activity and N uptake decreased. The decreases probably occurred as a result of the transition from vegetative growth to flowering in March. The shoot N concentrations in April (maturity stage) declined to 58% of those measured in March (data not shown). In addition, the shoot C/N ratios of hairy vetch, white clover and red clover were lower than those of crimson clover and pink clover; represented as the potential mineralization of those plant residues. CitationIt was consistent with the previous finding by Brandsaeter et al. (2008) that mineralization rates of N derived from hairy vetch and white clover organic residues were higher than those from crimson clover. CitationSeneviratne (2000) reported that rapid N mineralization generally occurred in plant residues with lower C/N ratios and lower lignin and polyphenol contents.
In the pot experiments, the decrease in shoot P uptake of hairy vetch in April, even though biomass increased, may have occurred because the number of yellow leaves increased, senesced and were shed in April, as also reported by CitationAnugroho et al. (2009a), and hairy vetch displayed significantly higher shoot K uptake than clover species during the vegetative growth stage (February), but no other stage. Shoot K uptake decreased after the flowering stage, as also reported from studies in temperate regions (CitationCaballero et al. 1996; CitationSainju et al. 1998) and subtropical regions (CitationAnugroho et al. 2009a). The decrease in P and K uptakes may have been related to the decrease in P and K concentrations and N-fixing activity as reported by (CitationHogh-Jensen 2003; CitationOlivera et al. 2004).
The present study suggests that hairy vetch is a more appropriate winter legume cover crop than clover species in subtropical regions under either P-limiting or P-non-limiting soil conditions. We confirmed that hairy vetch can be grown as a cover crop in four different soils with no added fertilizer. On acid soils in subtropical regions, low inputs of chemical fertilizer and high inputs of organic matter derived from hairy vetch may be a more sustainable agricultural system than systems based on high chemical fertilizer inputs and clover cover crops. Because of its low C/N ratio, hairy vetch may be a suitable green manure for vegetable crops with high N requirements. Further study should focus on the distribution of inorganic N in subtropical soils produced by incorporating or mulching with hairy vetch. This will better determine the suitability of hairy vetch as a low-cost green manure for meeting the N demand of subsequent high-yielding vegetable crops.
Acknowledgment
This study was supported in part by a Grant-in-Aid No. 17580055 for Scientific Research from the Japan Society for the Promotion of Science.
Related Research Data
References
- Anugroho , F and Kitou , M . 2007 . Growth and nutrient uptake of narrow leaf vetch and tiny vetch in subtropical region (Okinawa). The 46th Meeting of the Weed Science Society of Japan . J. Weed Sci. Tech. , 52 ( Suppl ) : 242 – 243 .
- Anugroho , F , Kitou , M , Nagumo , F , Kinjo , K and Tokashiki , Y . 2009a . Growth, N fixation, and nutrient uptake of hairy vetch as a cover crop in a subtropical region . Weed Biol. Manag. , 9 ( 1 ) : 63 – 71 .
- Anugroho , F , Kitou , M , Nagumo , F , Kinjo , K and Tokashiki , Y . 2009b . Effect of the sowing date on the growth of hairy vetch (Vicia villosa)as a cover crop influenced the weed biomass and soil chemical properties in a subtropical region . Weed Biol. Manag. , 9 ( 2 ) : 129 – 136 .
- Bayer , C , Martin-Neto , L , Mielniczuk , J , Pillon , CN and Sangoi , L . 2001 . Change in soil organic matter fractions under subtropical no-till cropping systems . Soil Sci. Soc. Am. J. , 65 : 1473 – 1478 .
- Bayer , C , Mielniczuk , J , Amado , TJC , Martin-Neto , L and Fernandes , SV . 2000 . Organic matter storage in a sandy clay loam Acrisol affected by tillage and cropping systems in southern Brazil . Soil Till. Res. , 54 : 101 – 109 .
- Brandsaeter , LO , Heggen , H , Riley , H , Stubhaug , E and Henriksen , TM . 2008 . Winter survival, biomass accumulation and N mineralization of winter annual and biennial legumes sown at various times of year in Northern temperate regions . Eur. J. Agron. , 28 : 437 – 448 .
- Caballero , R , Arauzo , M and Hernaiz , PJ . 1996 . Accumulation and redistribution of mineral elements in common vetch during pod filling . Agron. J. , 88 : 801 – 805 .
- Den Hollander , NG , Bastiaans , L and Kropff , MJ . 2007 . Clover as a cover crop for weed suppression in an intercropping design. II. Competitive ability of several clover species . Eur. J. Agron. , 26 : 104 – 112 .
- Duke , JA . 1981 . “ Vicia villosaRoth ” . In Handbook of Legumes of World Economic Importance , Edited by: Duke , JA . 284 – 286 . New York : Plenum Press .
- Hafner , H , George , E , Bationo , A and Marschner , H . 1993 . Effect of crop residues on root growth and phosphorus acquisition of pearl millet in an acid sandy soil in Niger . Plant Soil , 150 : 117 – 127 .
- Hogh-Jensen , H . 2003 . The effect of potassium deficiency on growth and N2-fixation in Trifolium repens . Physiol. Plant. , 119 : 440 – 449 .
- Kitou , M , Matuoka , H , Konndo , Y and Uchida , N . 2009 . Comparison of low P tolerant leguminous plants using akadama soil, and effect of root development on low P tolerance of Sesbania cannabina . Jpn. J. Soil Sci. Plant Nutr. , 80 : 487 – 493 .
- Liao , H , Wan , H , Shaff , J , Wang , X , Yan , X and Kochian , LV . 2006 . Phosphorus and aluminum interactions in soybean in relation to aluminum tolerance: exudation of specific organic acids from different regions of the intact root system . Plant Physiol. , 141 : 674 – 684 .
- Mulvaney , RL . 1996 . “ Nitrogen-Inorganic forms ” . In Methods of Soil Analysis , Edited by: Sparks , DL . 1123 – 1184 . Madison, WI : Soil Science Society of America (SSSA) .
- Olivera , M , Tejera , N , Iribarne , C , Ocana , A and Lluch , C . 2004 . Growth, nitrogen fixation and ammonium assimilation in common bean (Phaseolus vulgaris): effect of phosphorus . Physiol. Plant. , 119 : 440 – 449 .
- Oyedele , DJ and Aina , PO . 1998 . A study of soil factors in relation to erosion and yield of maize on a Nigerian soil . Soil Till. Res. , 48 : 115 – 125 .
- Sainju , UM , Sing , BP and Yaffa , S . 2002 . Soil organic matter and tomato yield following tillage, cover cropping, and nitrogen fertilization . Agron. J. , 94 : 594 – 602 .
- Sainju , UM , Singh , BP and Whitehead , WF . 1998 . Cover crop root distribution and its effects on soil nitrogen cycling . Agron. J. , 90 : 511 – 518 .
- Samarajeewa , KBDP , Horiuchi , T and Oba , S . 2005 . Effect of Chinese milk vetch (Astragalus sinicusL.) as a cover crop on weed control, growth and yield of wheat under different tillage systems . Plant Prod. Sci. , 8 ( 1 ) : 79 – 85 .
- Sanchez , PA and Logan , TJ . 1992 . “ Myths and science about the chemistry and fertility of soils in the tropics ” . In Myths and science of soils of the tropics , Edited by: Lal , R and Sanches , PA . 35 – 46 . Madison, WI : Soil Science Society of America (SSSA) . Spec. Publ. 29.
- Scott , BJ , Ridley , AM and Conyers , MK . 2000 . Management of soil acidity in long-term pastures of south-eastern Australia: a review . Aust. J. Exp. Agric. , 40 : 1173 – 1198 .
- Seneviratne , G . 2000 . Litter quality and nitrogen release in tropical agriculture: a synthesis . Biol. Fertil. Soils , 31 : 60 – 64 .
- Tiessen , H , Cuevas , E and Chacon , P . 1994 . The role of soil organic matter stability in soil fertility and agricultural potential . Nature , 371 : 783 – 785 .
- Yoshida , S . 1982 . Nitrogen fixation of Aechynomene indicagrown under upland and submerged conditions . Jpn J. Trop. Agric. , 26 ( 2 ) : 74 – 79 .
- Zanatta , JA , Bayer , C , Dieckow , J , Vieira , FCB and Mielniczuk , J . 2007 . Soil organic carbon accumulation and carbon costs related to tillage, cropping systems and nitrogen fertilization in a subtropical Acrisol . Soil Till. Res. , 94 : 510 – 519 .
- Zougmore , R , Nagumo , F and Hoshikawa , A . 2006 . Nutrient uptakes and maize productivity as affected by tillage system and cover crops in a subtropical climate at Ishigaki, Okinawa, Japan . Soil Sci. Plant Nutr. , 52 : 509 – 518 .