
Abstract
Objectives
Comparison of redox conditions in malignant and benign tumours is essential for understanding the role of reactive oxygen species in the pathophysiology of aggressive cancer profiles. Here we compare antioxidative systems in highly malignant brain tumour – glioblastoma multiforme (GBM), and in meningioma, a benign brain tumour.
Methods
Tumour tissues and blood of 67 GBM patients (mean age: 52.9 ± 11.5 years) and 67 meningioma patients (59.2 ± 10.2 years), and blood of 30 control subjects (50.8 ± 12.8 years) were analysed via biochemical assays.
Results
Components of glutathione system, which is responsible for H2O2 removal, showed lower activity/level in GBM: glutathione peroxidase (GBM: 9.90 ± 0.22; meningioma: 11.78 ± 0.23 U/mg of proteins; P < 0.001), glutathione reductase (GBM: 3.83 ± 0.13; meningioma: 4.67 ± 0.11 U/mg of proteins; P < 0.001), and glutathione (GBM: 6.70 ± 0.12; meningioma: 7.58 ± 0.14 μmol/g of tissue; P < 0.001). In contrast, the rank order of glutathione reductase activity and glutathione level in erythrocytes was: GBM > meningioma > control. Superoxide dismutase and catalase activities were lower in the blood of cancer patients compared to controls.
Discussion
Cells of malignant brain tumour show down-regulated antioxidative system which might result in increased levels of H2O2 compared to benign tumour tissue.
Introduction
Altered redox settings have been implicated in cancerogenesis and mutagenesis, tumour cell proliferation, development of metastases, and resistance to chemo- and radiotherapy.Citation1–Citation5 Tumour cells show: (i) accelerated metabolism which might result in increased baseline production of reactive oxygen species (ROS); (ii) altered activity of other sources of ROS; (iii) non-physiological activity of antioxidative system (AOS); (iv) inadequate oxygen supply, which might promote ROS production; and (v) dysfunctional redox signalling (e.g. many critical mutations in oncogene proteins affect redox-sensitive regulatory moieties).Citation3–Citation6 The most important ROS in tumour pathophysiology appears to be hydrogen peroxide (H2O2). High levels of H2O2 have been observed in cancer cells,Citation6,Citation7 which could be related to both promoted production and inefficient removal by AOS. Hydrogen peroxide is produced in mitochondria or cytosol from superoxide via superoxide dismutase (SOD). It is removed by catalase (CAT) or via glutathione system composed of glutathione, glutathione peroxidase (GSH-Px), and glutathione reductase (GR). H2O2 is uncharged and relatively stable ROS that can cross nuclear/cellular membrane.Citation8 Inside nucleus, H2O2 can react with copper to produce notoriously reactive hydroxyl radical that provokes DNA alterations.Citation9 Another pathway for hydroxyl radical production is the reaction between ferrous iron and H2O2 (Fenton reaction). In this respect, a very recent study has shown that the ratio between ferrous and ferric iron increases with brain tumour malignancy grade.Citation10 Furthermore, it has been found that cancer cells can release H2O2 in order to trigger autophagy in nearby cells, which leads to the production of high-energy nutrients that are then used by cancer cells.Citation11 Hydrogen peroxide also acts as a signalling molecule targeting a number of proteins and signalling cascades.Citation8 Altogether, it appears that H2O2 could represent a multipurpose ‘tool’ in cancerogenesis.
Redox conditions in tumours could be assessed by determining the AOS profile of tumour tissue, but also by measuring the activity of AOS enzymes in some normal tissue that is affected by tumour, such as blood.Citation12–Citation15 Large tumour masses are likely to exert oxidative pressure on erythrocytes. For example, drastic changes of AOS in erythrocytes have been reported to occur after surgical removal of brain tumours.Citation16 A number of studies have shown non-physiological activity of specific AOS components in tumour cells and blood of cancer patients.Citation17–Citation19 However, data on brain tumours and comparative analysis of malignant and benign tumours are still scarce and inconclusive. The examination and comparative analysis of redox conditions in malignant and benign tumours could be particularly important for understanding the role of H2O2 and other ROS in the pathophysiology of cancers with aggressive profiles.Citation20–Citation22
Here, we aimed to compare AOS (SOD, CAT, GSH-Px, GR, and glutathione) in tumour tissue and blood of patients with two types of brain tumours that are on the opposite ends of severity scale: glioblastoma multiforme (GBM), which is the most malignant form of primary brain tumours, and benign meningioma.
Materials and methods
Patients and sample collection
We analysed tumour tissues of 67 patients with GBM (WHO grade IV) (mean age: 52.9 ± 11.5 years; range: 22–76 years; sex ratio m/f = 36/31), and tumour tissues of 67 patients with meningioma (mean age: 59.2 ± 10.2 years; range: 30–74 years; m/f = 44/23). In addition, blood samples of all GBM and meningioma patients and of 30 healthy control subjects (mean age: 50.8 ± 12.8 years; range: 23–71 years; sex ratio m/f = 14/16) were analysed. The exclusion criteria were: viral infections, antioxidative therapy and dietary supplements, and smoking. The research was performed in accordance with the Declaration of Helsinki of the World Medical Association and has been approved by the local Ethics Committee. Written informed consents were obtained from all participants.
Tissue samples were collected in the course of total or sub-total tumour resection or tumour biopsy. All specimens were obtained from tumour peripheral parts avoiding necrotic areas. The meninges, blood vessels, and damaged/cauterized tissues were removed in −20°C chamber. The tissue was cut in smaller pieces, samples were briefly washed of blood with ice-cold 0.9% NaCl solution, and then snap-frozen in liquid nitrogen. GBM and meningioma were confirmed by histopathological analysis of surgical resections in accordance with the WHO criteria.Citation23
Fresh venous blood (5 ml) samples were collected from each patient two days before surgery and from control subjects using heparinized vacutainers. Erythrocytes were separated from the plasma, and washed three times with 0.9% NaCl solution by centrifugation at 3000 g/10 minutes/4°C. Following the procedure, erythrocytes and plasma samples were immediately frozen in liquid nitrogen. All samples were stored at −80°C until further analysis.
Biochemical assays
Frozen tumour tissue samples were thawed on ice, and 1 g of tissue was homogenized in 10 volumes (w/v) of ice-cold homogenizing buffer (50 mM Tris-HCl, 250 mM sucrose, 1 mM EDTA, pH 7.4) using homogenizer and sonication (10 kHz; 6 × 10 seconds). The homogenates were centrifuged at 100,000 g/90 minutes/4°C. In tissue samples, total SOD activity (manganese SOD (MnSOD) + copper-zinc SOD (CuZnSOD)) was measured according to the adrenalin method of Misra and Fridovich.Citation24 Previously described methods were used to determine the activities of CAT,Citation25 GSH-Px,Citation26 and GR.Citation27 The activity of enzymes in the tumour tissue was expressed as U/mg of proteins. Protein contents in supernatants were determined using the Lowry method. The total concentration of glutathione (reduced and oxidized) in tissue extracts was determined based on the Griffith method,Citation28 and presented as μmol/g of tissue.
Erythrocytes (0.5 ml) were lysed by adding 3 ml of ice-cold distilled water. SOD (i.e. CuZnSOD; erythrocytes do not contain MnSOD), CAT, GSH-Px, and GR activities were determined using the same methods as for tissue samples. Interference from haemoglobin (Hb) was eliminated by precipitation prior to the assay using ethanol/chloroform (1 : 1, v/v) that was followed by centrifugation at 3000 g/5 minutes/4°C.Citation29 The activity of enzymes in erythrocytes was presented as U/g of Hb. Hb concentration was measured by applying the Drabkin method. Finally, the total concentration of glutathione in plasma was determined based on the Griffith method and presented as μmol/l of plasma. All chemicals were purchased from Sigma-Aldrich (St Louis, MO, USA) or Merck (Darmstadt, Germany).
Statistical analysis
Statistical differences were evaluated by means of Student's t-test using STATISTICA 8.0 (StatSoft Inc., Tulsa, OK, USA). The results are presented as means ± standard error and were taken to be statistically different if P < 0.05.
Results
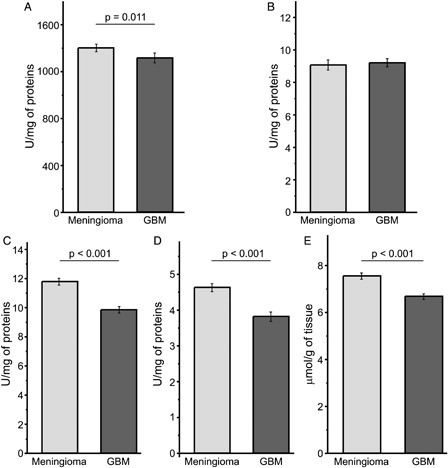
It can be observed that GBM tissue showed a slightly lower total activity of SOD (CuZnSOD + MnSOD) compared to meningioma (A), while CAT activity did not differ between the two tumour types (B). On the other hand, glutathione system was down-regulated in GBM compared to meningioma, i.e. activities of GSH-Px and GR (C and D), and the total glutathione level (E) were significantly lower in GBM tissue.
Figure 1. AOS in tumour tissues of GBM (n = 67) and meningioma patients (n = 67). (A) Total activity of SOD; (B) the activity of CAT; (C) the activity of GSH-Px; (D) the activity of GR; (E) total concentration of glutathione.
SOD activity in erythrocytes of GBM patients was significantly lower compared to control values. This trend was observed in meningioma as well (A). CAT showed a lower activity in both GBM and meningioma compared to controls (B). In contrast, the glutathione system, besides GSH-Px which showed no differences between study groups (C), was up-regulated in the blood of cancer patients. The rank order of GR activity in erythrocytes (D), and of total glutathione level in the plasma (E) was GBM > meningioma > control.
Figure 2. AOS in the blood of control subjects (Ctrl; n = 30), GBM (n = 67), and meningioma (n = 67) patients. (A) CuZnSOD activity in erythrocytes; (B) CAT activity in erythrocytes; (C) GSH-Px activity in erythrocytes; (D) GR activity in erythrocytes; (E) total concentration of glutathione in plasma.

Discussion
We compared AOS in GBM, as a model of highly malignant, aggressive brain tumour, and meningioma, which shows benign phenotype in the prevailing number of cases (∼98%). Hydrogen peroxide-removing glutathione system showed a lower activity in GBM tissue compared to meningioma. This is in accordance with previous findings of one smaller study (conducted on 26 GBM and 22 meningioma patients) that found a lower GSH-Px and GR activity in GBM compared to meningioma.Citation30 In addition, two comparative studies on meningioma and astrocytoma found that malignant brain tumours show a lower activity of GSH-Px (the rank order was: meningioma > low-grade astrocytoma > high-grade astrocytoma),Citation31 as well as lower glutathione concentration and SOD activity.Citation31,Citation32 It is noteworthy that Palani et al.Citation33 have shown that GSH-Px, GR, and SOD activity in gliomas gradually decrease with increasing tumour grade. In discrepancy to our findings, Pu et al.Citation31 have found higher CAT activity in meningioma compared to high-grade astrocytoma. This might be related to the fact that the high-grade astrocytoma group in that particular study included GBM and grade 3 astrocytoma patients, and potentially due to the lower number of patients (10 meningioma and 10 high-grade astrocytoma).
A lower activity of H2O2-removing system implies a down-regulation of enzymes that most likely allows the accumulation of H2O2. In relation to this, the increased level of oxidation in GBM compared to meningioma has been documented in previous studies using different oxidation markers, such as oxidized proteins which were higher in GBM,Citation30 and the level of reduced glutathione which was lower in GBM.Citation34 In addition, the level of lipid peroxidation was significantly higher in astrocytoma compared to meningioma.Citation32 Pro-oxidative settings in GBM, and in meningioma as well (but to a lower level), are further implicated by the results obtained on blood. GR activity in erythrocytes and glutathione level in the plasma showed the rank order: GBM > meningioma > control. These settings in the blood reflect a prolonged release of H2O2 from brain tumour mass into the circulation. This is in accordance with previous findings showing an increased level of markers of lipid peroxidation in the blood of glioblastoma and meningioma patients.Citation19,Citation35 In addition, depletion of antioxidants has been observed in a cohort of patients with different brain tumours.Citation36 Furthermore, a decreased activity of CuZnSOD and CAT was observed in erythrocytes of brain tumour patients compared to healthy subjects. The former might be explained by H2O2-provoked inhibition of CuZnSOD.Citation15 On the other hand, potential mechanisms that led to the suppression of CAT activity in erythrocytes remain blur, but it is important to point up different roles of two H2O2-removing enzymes – GSH-Px and CAT in erythrocytes’ antioxidative defence. GSH-Px is mainly located near the erythrocyte's membrane, and it is responsible for preventing H2O2 influx and lipid peroxidation. CAT is the primary erythrocyte enzyme responsible for coping with endogenous H2O2,Citation37 and the production of H2O2 in erythrocytes is decreased when CuZnSOD is inhibited. Rao et al.Citation38 have found a decreased SOD activity in erythrocytes of glioma and meningioma patients compared to control subjects. CAT was slightly decreased in patients compared to controls, whereas GSH-Px did not show any differences between groups. All this matches well with our findings. Surprisingly, their study has shown a drastically lower GR activity in cancer patients. We do not have an explanation for such discrepancy, but it is worth mentioning that control GR activity was four times lower compared to our results, whereas control values for other enzymes were similar.
A potential mechanism of down-regulation of glutathione system in tumour tissue might involve the shift of cysteine (glutathione precursor) towards a metabolic pathway that appears to be specific for glioblastoma. In brief, glioblastoma cells show an increased expression of cysteine dioxygenase 1, which catalyses the reaction of cysteine with molecular oxygen in order to produce cysteine sulfinic acid. It has been proposed that this metabolite is involved in the development and growth of aggressive tumour phenotypes.Citation39 In addition, specific enzymes that are involved in glutathione biosynthesis and degradation, such as glutamate cysteine ligase and gamma glutamyl transferase, might show a non-physiological activity in tumour cells resulting in dysfunctional glutathione system.Citation40–Citation42
GBM appears to be significantly less susceptible to radiotherapy in comparison to meningioma. Radiotherapy in GBM patients results in a modest improvement of survival period.Citation43 On the other hand, surgical approach combined with post-operative radiotherapy in selected cases is considered to be the most appropriate for meningioma patients,Citation44 and radiation is highly effective in the management of skull-base meningioma, which otherwise require complex combined surgical approaches.Citation45 Radiotherapy is generally mediated via induction of oxidative damage. Therefore, higher radioresistance of tumour cells that show weaker AOS performance might appear paradoxical. In normal cells unaccustomed to pronounced oxidation, glutathione depletion activates cell death pathways and promotes radiosensitivity.Citation46,Citation47 Unfortunately, in spite of down-regulated AOS defence and pro-oxidative conditions, GBM cells grow, proliferate, and show high radioresistance. It is plausible that further increase of oxidation via radiation might have little effect on cells that are already proficient in operating under pro-oxidative conditions and even appear to require supraphysiological ROS production. As a matter of fact, the inhibition of specific enzymes that are involved in ROS production, such as cyclooxygenase-2 (has superoxide as a by-product) or poly (ADP-ribose) polymerase 1 (promotes ROS production in mitochondria), has been reported to result in increased radiosensitivity of glioblastoma.Citation48,Citation49 According to our results, the up-regulation of the glutathione system might also represent a useful strategy in fighting glioblastoma. Some efforts have been already made in this direction. For example, a recent study has shown strong in vitro effects of dimethyl fumarate, which is known to activate the expression of several glutathione-related enzymes, against glioma cells.Citation50
Disclaimer statements
Contributors VB – conceiving and designing the study, obtaining funding and/or ethics approval, collecting the data, analyzing the data, interpreting the data, writing the article in part; MB – conceiving and designing the study, obtaining funding and/or ethics approval, collecting the data, analyzing the data, interpreting the data, writing the article in part; IS – analyzing the data, interpreting the data, writing the article in part, revising the article.
Funding This study was supported by the Ministry of Education and Science of the Republic of Serbia, grant number OI173014.
Conflicts of interest None.
Ethics approval The research was performed in accordance with the Declaration of Helsinki of the World Medical Association and has been approved by the Ethics Committee of the School of Medicine, University of Belgrade.
Acknowledgement
We thank Miss Verica Bogdanovic for excellent technical assistance.
References
- Bakalova R, Zhelev Z, Aoki I, Saga T. Tissue redox activity as a hallmark of carcinogenesis: from early to terminal stages of cancer. Clin Cancer Res 2013;19(9):2503–17.
- Sotgia F, Martinez-Outschoorn UE, Lisanti MP. Mitochondrial oxidative stress drives tumour progression and metastasis: should we use antioxidants as a key component of cancer treatment and prevention? BMC Med 2011;9:62.
- Valko M, Leibfritz D, Moncol J, Cronin MT, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol 2007;39(1):44–84.
- Sosa V, Moliné T, Somoza R, Paciucci R, Kondoh H, LLeonart ME. Oxidative stress and cancer: an overview. Ageing Res Rev 2013;12(1):376–90.
- Goswami PC. Mutant mitochondria and cancer cell metastasis: quest for a mechanism. Cancer Biol Ther 2009;8(14):1386–8.
- Hempel N, Bartling TR, Mian B, Melendez JA. Acquisition of the metastatic phenotype is accompanied by H2O2-dependent activation of the p130Cas signaling complex. Mol Cancer Res 2013;11(3):303–12.
- López-Lázaro M. Dual role of hydrogen peroxide in cancer: possible relevance to cancer chemoprevention and therapy. Cancer Lett 2008;252(1):1–8.
- Spasojević I, Jones DR, Andrades ME. Hydrogen peroxide in adaptation. Oxid Med Cell Longev 2012;2012:596019.
- Lee DH, O'Connor TR, Pfeifer GP. Oxidative DNA damage induced by copper and hydrogen peroxide promotes CG-->TT tandem mutations at methylated CpG dinucleotides in nucleotide excision repair-deficient cells. Nucleic Acids Res 2002;30(16):3566–73.
- Wandzilak A, Czyzycki M, Wrobel P, Szczerbowska-Boruchowska M, Radwanska E, Adamek D, et al. The oxidation states and chemical environments of iron and zinc as potential indicators of brain tumour malignancy grade – preliminary results. Metallomics 2013;5(11):1547–53.
- Pavlides S, Whitaker-Menezes D, Castello-Cros R, Flomenberg N, Witkiewicz AK, Frank PG, et al. The reverse Warburg effect: aerobic glycolysis in cancer associated fibroblasts and the tumour stroma. Cell Cycle 2009;8(23):3984–4001.
- Nikolić-Kokić A, Blagojević D, Spasić M. Complexity of free radical metabolism in human erythrocytes. J Biochem Med 2013;29(3):189–95.
- Demirci S, Ozsaran Z, Celik HA, Aras AB, Aydin HH. The interaction between antioxidant status and cervical cancer: a case control study. Tumori 2011;97(3):290–5.
- Arsova-Sarafinovska Z, Matevska N, Eken A, Petrovski D, Banev S, Dzikova S, et al. Glutathione peroxidase 1 (GPX1) genetic polymorphism, erythrocyte GPX activity, and prostate cancer risk. Int Urol Nephrol 2009;41(1):63–70.
- Ferruzzi E, Franceschini R, Cazzolato G, Geroni C, Fowst C, Pastorino U, et al. Blood glutathione as a surrogate marker of cancer tissue glutathione S-transferase activity in non-small cell lung cancer and squamous cell carcinoma of the head and neck. Eur J Cancer 2003;39(7):1019–29.
- Woźniak B, Woźniak A, Kasprzak HA, Drewa G, Mila-Kierzenkowska C, Drewa T, et al. Lipid peroxidation and activity of some antioxidant enzymes in patients with glioblastoma and astrocytoma. J Neurooncol 2007;81(1):21–6.
- Glorieux C, Dejeans N, Sid B, Beck R, Calderon PB, Verrax J. Catalase overexpression in mammary cancer cells leads to a less aggressive phenotype and an altered response to chemotherapy. Biochem Pharmacol 2011;82(10):1384–90.
- Estrela JM, Ortega A, Obrador E. Glutathione in cancer biology and therapy. Crit Rev Clin Lab Sci 2006;43(2):143–81.
- Martin RC, Li Y, Liu Q, Barker DF, Doll MA, Hein DW. Manganese superoxide dismutase expression as a function of genotype and lung cancer pathology. Cancer Invest 2010;28(8):813–9.
- Young O, Crotty T, O'Connell R, O'Sullivan J, Curran AJ. Levels of oxidative damage and lipid peroxidation in thyroid neoplasia. Head Neck 2010;32(6):750–6.
- Demasi AP, Furuse C, Soares AB, Altemani A, Araújo VC. Peroxiredoxin I, platelet-derived growth factor A, and platelet-derived growth factor receptor alpha are overexpressed in carcinoma ex pleomorphic adenoma: association with malignant transformation. Hum Pathol 2009;40(3):390–7.
- Skrzycki M, Scibior D, Podsiad M, Czeczot H. Activity and protein level of CuZnSOD and MnSOD in benign and malignant liver tumors. Clin Biochem 2008;41(1–2):91–6.
- Kleihues P, Louis DN, Scheithauer BW, Rorke LB, Reifenberger G, Burger PC, et al. The WHO classification of tumours of the nervous system. J Neuropathol Exp Neurol 2002;61(3):215–25.
- Misra HP, Fridovich I. The role of superoxide-anion in the autooxidation of epinephrine and a simple assay for superoxide dismutase. J Biol Chem 1972;247(10):3170–5.
- Beutler E. Red cell metabolism. New York: Grune and Stratton; 1982. p. 102.
- Paglia DE, Valentine WN. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med 1967;70(1):74–7.
- Glatzle D, Vuilleumier JP, Weber F, Decker K. Glutathione reductase test with whole blood a convenient procedure for the assessment of the riboflavin status in humans. Experientia 1974;30(6):665–8.
- Griffith OW. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vynilpiridine. Anal Biochem 1980;106(2):207–12.
- Tsuchihashi M. Zur Kenntnis der Blutkatalase. Biochem Z 1923;140:65–74.
- Tanriverdi T, Hanimoglu H, Kacira T, Sanus GZ, Kemerdere R, Atukeren P, et al. Glutathione peroxidase, glutathione reductase and protein oxidation in patients with glioblastoma multiforme and transitional meningioma. J Cancer Res Clin Oncol 2007;133(9):627–33.
- Pu PY, Lan J, Shan SB, Huang EQ, Bai Y, Guo Y, et al. Study of the antioxidant enzymes in human brain tumors. J Neurooncol 1996;29(2):121–8.
- Zengin E, Atukeren P, Kokoglu E, Gumustas MK, Zengin U. Alterations in lipid peroxidation and antioxidant status in different types of intracranial tumours within their relative peritumoral tissues. Clin Neurol Neurosurg 2009;111(4):345–51.
- Palani M, Arunkumar R, Janardhanam VA. Biochemical and cytogenetic analysis of brain tissues in different grades of glioma patients. Ann Neurosci 2010;17(3):120–5.
- Kudo H, Mio T, Kokunai T, Tamaki N, Sumino K, Matsumoto S. Quantitative analysis of glutathione in human brain tumors. J Neurosurg 1990;72(4):610–5.
- Yilmaz N, Dulger H, Kiymaz N, Yilmaz C, Bayram I, Ragip B, et al. Lipid peroxidation in patients with brain tumor. Int J Neurosci 2006;116(8):937–43.
- Aggarwal S, Subberwal M, Kumar S, Sharma M. Brain tumor and role of beta-carotene, a-tocopherol, superoxide dismutase and glutathione peroxidase. J Cancer Res Ther 2006;2(1):24–7.
- Bajčetić M, Otašević B, Prekajski NB, Spasić S, Spasojević I. Antioxidative system in the erythrocytes of preterm neonates with sepsis: the effects of vitamin E supplementation. Ann Clin Biochem 2014;51(5):550–6.
- Rao GM, Rao AV, Raja A, Rao S, Rao A. Role of antioxidant enzymes in brain tumours. Clin Chim Acta 2000;296(1–2):203–12.
- Prabhu A, Sarcar B, Kahali S, Yuan Z, Johnson JJ, Adam KP, et al. Cysteine catabolism: a novel metabolic pathway contributing to glioblastoma growth. Cancer Res 2013;74(3):787–96.
- Hanigan MH1, Frierson HF Jr, Swanson PE, De Young BR. Altered expression of gamma-glutamyl transpeptidase in human tumors. Hum Pathol 1999;30(3):300–5.
- Corti A, Franzini M, Paolicchi A, Pompella A. Gamma-glutamyltransferase of cancer cells at the crossroads of tumor progression, drug resistance and drug targeting. Anticancer Res 2010;30(4):1169–81.
- Backos DS, Fritz KS, McArthur DG, Kepa JK, Donson AM, Petersen DR, et al. Glycation of glutamate cysteine ligase by 2-deoxy-d-ribose and its potential impact on chemoresistance in glioblastoma. Neurochem Res 2013;38(9):1838–49.
- Keime-Guibert F, Chinot O, Taillandier L, Cartalat-Carel S, Frenay M, Kantor G, et al. Radiotherapy for glioblastoma in the elderly. N Engl J Med 2007;356(15):1527–35.
- Vranic A, Peyre M, Kalamarides M. New insights into meningioma: from genetics to trials. Curr Opin Oncol 2012;24(6):660–5.
- Minniti G, Amichetti M, Enrici RM. Radiotherapy and radiosurgery for benign skull base meningiomas. Radiat Oncol 2009;4:42.
- Franco R, Cidlowski JA. Glutathione efflux and cell death. Antioxid Redox Signal 2012;17(12):1694–713.
- Bump EA, Brown JM. Role of glutathione in the radiation response of mammalian cells in vitro and in vivo. Pharmacol Ther 1990;47(1):117–36.
- Kang KB, Wang TT, Woon CT, Cheah ES, Moore XL, Zhu C, et al. Enhancement of glioblastoma radioresponse by a selective COX-2 inhibitor celecoxib: inhibition of tumour angiogenesis with extensive tumour necrosis. Int J Radiat Oncol Biol Phys 2007;67(3):888–96.
- Venere M, Hamerlik P, Wu Q, Rasmussen RD, Song LA, Vasanji A, et al. Therapeutic targeting of constitutive PARP activation compromises stem cell phenotype and survival of glioblastoma-initiating cells. Cell Death Differ 2014;21(2):258–69.
- Ghods AJ, Glick R, Braun D, Feinstein D. Beneficial actions of the anti-inflammatory dimethyl fumarate in glioblastomas. Surg Neurol Int 2013;4:160.