
Abstract
This paper investigates changes in subsistence strategies at the upland cave site of Nugljanska (Croatia) at the end of the last ice age, during a time of rapid sea level rise and changing environment. We analysed the faunal assemblage from archaeological levels spanning the Pleistocene–Holocene transition (15,000–8000 years BP) and evaluated changes in relative abundance of species, measured species diversity, and compared the representation of terrestrial mammal and marine resources. We found a significant shift in the most abundant prey species exploited (from red deer, Cervus, to wild boar, Sus). There was some correlation between dietary diversification and periods of increased moisture availability and the spread of deciduous forest in the area. Our results suggest that there was a continuing reliance on terrestrial resources throughout time and that changes in dietary patterns were likely due to local environmental change and potentially, changing seasonal mobility strategies, at the Pleistocene–Holocene transition.
Introduction
The rapid rate of sea level rise is now one of the dire results of modern climate change. In 1989, Tjeerd van Andel suggested that sea level rise at the end of the last ice age was the most important environmental change worldwide in the last 15,000 years. One of the most notable examples of the effects of rapid post-glacial sea level rise is the formation of the Adriatic Sea. The northern third of the sea became inundated in less than 5000 years, and the sea reached its current form around 8000 years ago (Lambeck et al. Citation2004). Prior to this, the Adriatic basin was a large expanse of low-lying, grassy flatland, known as the Great Adriatic Plain; the coast was located just north of Pescara, Italy and slightly south of Zadar, Croatia (Mussi Citation2001; Lambeck et al. Citation2004; Surić Citation2005; Miracle Citation2007).
Human groups living on the Great Adriatic Plain or near the coast would have been especially affected by sea level rise at the end of the last ice age, but low-lying inland areas also changed as groundwater tables ncreased and meltwater rivers contributed to the topography of the Adriatic region (Ferretti et al. Citation1986; Brambati Citation1992; Zavatarelli et al. Citation2000). While the largest-scale impact of sea level rise was the disappearance of the Great Adriatic Plain, smaller-scale ecosystem and microhabitat changes occurred as lowlands transitioned from open grassland ecosystems to estuaries, wetlands and coastal ecozones during the early Holocene. Prehistoric sites located on what was the Great Adriatic Plain are now inundated and inaccessible to archaeologists, but cave sites on the Istrian Peninsula would have been located in the hinterland and offer a glimpse into lifeways of people at the end of the last ice age up until the arrival of farming in the region ().
Figure 1. Location of Nugljanska and other hinterland cave sites in relation to post-glacial sea-level rise.
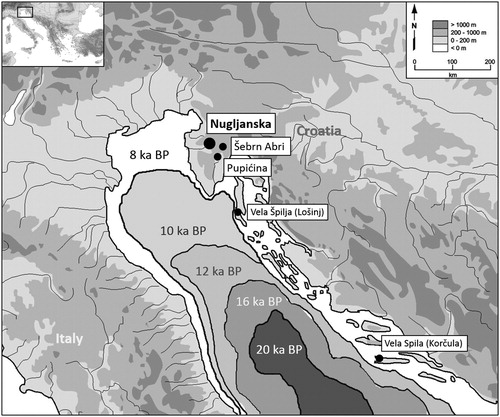
Table 1. Radiocarbon dates for the site of Nugljanska
In this paper, we present the upland cave site of Nugljanska as a case study for identifying and discussing human response to climate change and sea level rise and the associated environmental transformations at the Pleistocene–Holocene transition in the northeastern Adriatic. After introducing the site and its palaeoenvironmental context, we present the results of faunal analysis, assessing changes in species richness, faunal diversity and relative abundances to discuss subsistence strategies in a changing environment.
Nugljanska
Nugljanska is a large, south-west facing cave located 550 m above the sea level with a commanding view over an extensive plain () (Miracle and Forenbaher Citation2000). Its size and shape make it a good shelter (), and its southern exposure may have made it desirable for use even in the winter despite its elevation. It is easy to access in the present day and would have been a noticeable feature on the landscape in the past; elevation and slope are not hindrances to the use of the cave. The site is thought to be the earliest human settlement on the uplands of the Ćićarija Mountains (Komšo and Pellegatti Citation2007). It has been suggested that hunter-gatherers used Nugljanska as an outpost for monitoring resources during the Late Upper Palaeolithic, and as a residential base camp in the Mesolithic (Komšo Citation2006; Miracle Citation2007).
Figure 2. Photo of the view from Nugljanska cave in May 2010 (S.E.P.B.).

Figure 3. Photo of Nugljanska cave in May 2010 (S.E.P.B.).

The most recent excavation at Nugljanska took place in 1998 as part of the Pupićina Cave Project (Miracle and Forenbaher Citation2000). One large trench with four quadrants was excavated against the eastern wall of the cave, covering an area of 4 m2 and reaching a depth of 2·5 m. All sediments were dry-sieved using a 6 mm mesh and flotation samples were taken. The excavated levels contained material from the Late Upper Palaeolithic and Mesolithic (15–8 kya). All post-Mesolithic deposits were likely excavated by Moser at the end of the 19th century, although some deposits may remain in partially brecciated sediment near the cave walls (Barfield Citation1999). There are a large number of stone tool classes in the Late Upper Palaeolithic. This decreases in the uppermost Palaeolithic layers, when there is a large increase in hunting tool types and burins made of regional material (Komšo and Pellegatti Citation2007). The lithic finds from the Mesolithic levels (n = 367) are typical for the time period and region, consisting of endscrapers and linear tools (Komšo Citation2006). There is evidence for hearths and a large number of faunal remains as well as a small amount of worked bone have been recovered (Miracle and Forenbaher Citation2000). There are no known human burials. The stratigraphy at Nugljanska is thought to be in situ (Miracle and Forenbaher Citation2000). Five new radiocarbon dates were obtained on faunal material for this study ().
Environmental Context
Multiple proxies suggest open grassland mixed with herbaceous shrub and pine communities as the dominant groundcover south of the cave on the Great Adriatic Plain during the late glacial, and the expansion of deciduous species in the terminal Pleistocene (Schmidt et al. Citation2000; Boschian and Fusco Citation2007). Nugljanska was probably located within a forested setting, overlooking a plain which may have included some marshy or wet areas with the rise of the local water table at this time. During the Younger Dryas, local woodland was retreated and dry steppe expanded (Willis Citation1994; Miracle Citation2001). In contrast, the Trieste Karst to the north of the cave was a harsh, unwooded landscape experiencing cold and dry environmental conditions during the late glacial that persisted into the terminal Pleistocene and early Holocene (Boschian and Fusco Citation2007). These environmental conditions may have created a barrier to the movement of human populations during the time of increasing sea level further south.
There is evidence of a vegetative change from overall drier conditions to a more mixed deciduous landscape in the initial Holocene in the Adriatic region (Favaretto et al. Citation2008). Pollen evidence from the north and south of Istria attests to an increase in forest and broadleaf taxa including oak (Quercus) and hazel (Corylus) (Gallizia Vuerich and Princivalle Citation1994; Andrič et al. Citation2008; Favaretto et al. Citation2008; Voytek Citation2011). The presence of species such as hazel, hornbeam (Carpinus), linden (Tilia) and elm (Ulmus) indicate cool-temperate summers, with average temperature ranges between a minimum of –15°C in the winter and 10°C in the summer (Favaretto et al. Citation2008). Around 9000 BP the eastern Adriatic climate became mild and wet, with no-frost winters and moist summers (Rossignol-Strick Citation1999).
These climate-driven environmental changes would have strongly influenced human settlement patterns, seasonal site use and subsistence strategies at the Pleistocene–Holocene transition. Forested environments tend to foster larger numbers of small-bodied prey than do open ones. We expected a more diverse diet (higher species richness and evenness) in the levels dated to the late glacial as an indicator of environment rather than dietary stress as might be predicted by optimal foraging theory, where a ‘broad spectrum’ diet with relatively higher proportions of ‘lower ranked’ prey could be interpreted as a result of resource scarcity. The assemblage should include deer species, which exploit woodland edge; boar, which inhabit forested areas; and aurochs, which may have preferred marshy areas, as well as small game. Because of the site's location in the hinterland of the Great Adriatic Plain, taxa such as Equus spp. were not expected in the assemblage. In the Younger Dryas, we expected an increase in prey specialisation, reflecting the fact that larger-bodied animals in open environments are easier to hunt and provide higher returns than smaller-bodied animals in wooded environments (Kelly Citation1983). This would have been suitable habitat for large ungulates like red deer which, if migratory, would be a predictable seasonal resource, and the site may have been an ideal vantage point for their observation. By the early Holocene, the continuation of rapid sea level rise, combined with reforesting of the area, may have led to a broad spectrum diet, or one dominated by wood and wood-edge species such as boar and roe deer. A limited amount of coastal and marine resources were expected in Holocene levels, as the sea was still some distance from the cave and may have been outside the site exploitation area.
Materials and Methods
S.E.P.B. processed and identified the faunal assemblage following a protocol established by Miracle and Pugsley (Citation2006) using comparative skeletal material in the Grahame Clark Laboratory at the University of Cambridge. The faunal remains from Nugljanska had been in storage at the Arheološki Muzej Istre in Pula, Croatia, and were sent to Cambridge on loan. Faunal remains from Levels 3 through 8 + 8A, quadrants K12, K13 and L13 were included in the analysis. Faunal material from quadrant L12 was excluded due to the problematic nature of some of the stratigraphy. The temporal integrity of Levels 1 and 2 is unknown, and as these contexts showed signs of disturbance and mixture with modern material, they were discounted from analysis. In addition, excavated Levels 9–14 yielded very small sample sizes of poorly preserved bone that most likely date outside the time period of interest, and will not be discussed here. Birds and microfauna were excluded from the analysis because they had been previously separated from the assemblage. Small game quantified in this paper was not considered to be part of the microfauna assemblage during the initial sorting (not carried out by the authors). Molluscs were also treated separately from the mammalian assemblage and the data included here originate from the earlier tallies (P. Miracle, personal communication). Each identifiable specimen was coded separately and counted towards the total number of identified specimens (NISP). All studied levels were included in a linear model to see how strongly taxonomic richness might be correlated with sample size in each level. All statistics were computed using the Palaeontological Statistics (PAST) program (Hammer et al. Citation2001).
Results
Taphonomy
In total, 11,682 bones or bone fragments were excavated. Of the 4006 identifiable fragments (34% of the assemblage), 2128 (53%) were identifiable to element and taxon. About a quarter of the identifiable assemblage (26%) had recent breaks which could have occurred during excavation, curation or transportation of the bones. Weathering, calcium carbonate accumulation and gnawing were the most frequent taphonomic variables recorded in the assemblage and are discussed further here ().
Table 2. Frequency of bone modifications by level at Nugljanska
Natural Agents
Natural agents accounted for most of the modification of the faunal assemblage. Between 85 and 97% of the bones exhibited some weathering in all levels. Most weathering consisted of fine-line fracturing and spalling. In addition, most bone was covered with a light coating of calcium carbonate due to the karstic nature of the cave. Both of these influences on the assemblage may have caused some physical attrition, but are not thought to have caused major chemical modifications which may have altered the biomolecular preservation of the majority of the faunal remains, which have been isotopically analysed (Pilaar Birch et al. 2012).
Biological Agents
Gnawing was the most common type of modification and was found in all levels. It generally affected 12–26% of the assemblage. Between 85 and 95% of gnawing or digestion was classified as some combination of carnivore activity (rather than rodent or artiodactyl) in all levels. This suggests that the site was frequented by carnivores with varying degrees of intensity in all time periods. The percentage of burnt bone, indicative of human activity, ranges from as little as 1% in Level 7 to 17% in Level 6. Most bones are classified as ‘burnt’ (reddish) or partially carbonised. In Level 3, there is an increase in intensity of burning, as most burnt bone was fully carbonised or partially calcined. Cut marks, perhaps the most convincing evidence of human modification, are also found with low frequency in all levels. These ranges from marks left from dismembering and skinning to blunt chops and most often occurred on the long bones of the appendicular skeleton in medium- to large-sized game.
Mammalian Taxonomic Composition of the Faunal Assemblages
The distribution of taxa changes through time, and there is a notable shift in the representation of red deer (Cervus elaphus) and boar (Sus scrofa) in the terminal Pleistocene and initial Holocene levels, respectively (). Proportions of Cervus increase up to a maximum of almost 60% in the Pleistocene and fall to approximately 30% throughout the Holocene layers. In contrast, Sus, present in frequencies of about 10% throughout the Pleistocene contexts, constitute over half of the taxa in the Holocene layers. Roe deer (Capreolus capreolus) are always important as the third most numerous taxon, but their frequency is halved – from about 20% in the Pleistocene levels to 10% in the Holocene layers. Aurochs (Bos primigenius) decrease in proportion within the Pleistocene contexts and represent 5% of the assemblage in the terminal Pleistocene and Holocene levels. Frequencies of other game such as chamois (Rupicapra rupicapra) and ibex (Capra ibex) fluctuate through time.
Figure 4. Relative taxonomic abundances of terrestrial mammals at Nugljanska; n is equivalent to NISP. Small carnivore = fox, marten, beaver and Felis spp. Levels 3–5 fall in the Boreal period; Level 6 is early Younger Dryas/late Bølling-Allerød; and Levels 7 + 7A and 8 + 8A are Bølling-Allerød.
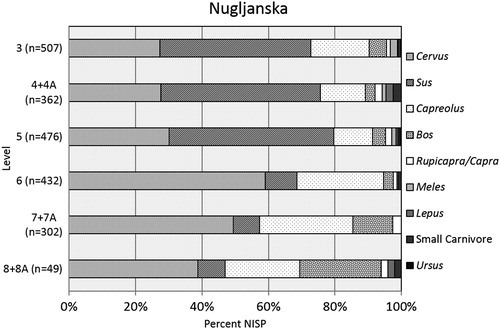
In addition to what were likely the key prey species for the people using Nugljanska, the remains of smaller mammals and carnivores are also present in varying abundances. Hare (Lepus) was very scarce in the assemblage. Fox (Vulpes) are rare throughout the sequence. A single incisor was identified to beaver (Castor) in Level 8 + 8A. Brown bear (Ursus) never contributed more than 1% of the assemblage, with two specimens in Level 5 (both astragali), and one in Level 3 (third metatarsal). Two specimens belonging to the genus Felis were identified in Level 6. A metacarpal was identified to the species Felis lynx, while a hemi-mandible was identified to Felis silvestris. There are no instances of badger (Meles meles) in the Pleistocene layers; the appearance of this small omnivore is a Holocene phenomenon. Due to the difficulty of discerning the pine marten (Martes martes) and the beech marten (Martes foina), these specimens were only assigned to genus level. As with badgers, they appear only in the Holocene and never contribute more than 1% to the assemblage.
Marine and Terrestrial Molluscs
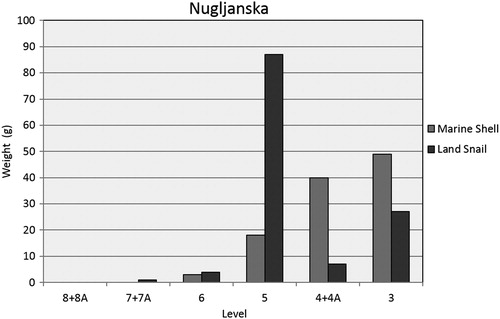
Identification of the mollusc shell from Nugljanska is based on existing tallies from excavation records. Marine shell first appears in Level 6, and is found in increasing abundances in all subsequent levels. Recovered marine taxa include the gastropod Columbella rustica (dove snail), C. edule (common cockle) and M. galloprovincialis (Mediterranean mussel). Edible land snails (Helix) have been recovered from the site in varying abundances, also first appearing in small quantities in Level 6 along with marine shell. They are most abundant in Level 5. summarises recovered shell by weight, which was the only data available at that time.
Figure 5. Marine and terrestrial molluscs by weight at Nugljanska.
Measures of Diversity
The discussion of dietary diversification necessitates a characterisation of the diversity of the faunal assemblages through more than just taxonomic composition. Here, we discuss the taxonomic richness, evenness and heterogeneity of the terrestrial mammal assemblage, as well as the balance between terrestrial mammals and molluscan resources.
Species Richness and Evenness
Species richness increases through time at Nugljanska (). Pleistocene Levels 8 + 8A and 7 + 7A both have six taxa and Level 6 has eight. This is followed by 9 taxa in Holocene Level 5 and 4 + 4A and an increase to 10 taxa in Level 3. When NISP and species richness were plotted in a linear model, the two were not significantly correlated (r2 = 0·52, P = 0·1) (A). This remained the case even if the smallest sample size (8 + 8A, n = 49) was removed (r2 = 0·69, P = 0·08). The ‘core’ prey taxa of Cervus, Capreolus, Sus and Bos are always present. The presence or absence of small game such as Lepus or small carnivores such as Vulpes is what affects the species richness of each level.
Figure 6. (A) A linear model of the relationship between sample size and species richness at Nugljanska. (B) A linear model of the relationship between sample size and evenness at Nugljanska.
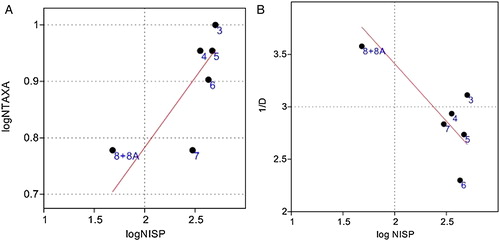
Table 3. NISP and diversity indices at Nugljanska.
Although all core prey species are always present, their relative representation changes, sometimes dramatically, between levels. The reciprocal of Simpson's index (1/D) measures the equity of distribution and at Nugljanska is not significantly correlated with sample size (r2 = 0·56, P = 0·1), suggesting changes in evenness are independent (B). The higher the value, the more relative abundances are evenly distributed. Level 6 has the least evenly distributed faunal assemblage (cf. with ), whereas levels 8 + 8A, 4 + 4A and 3 have the most evenly distributed faunal assemblages. Levels 5 and 7 + 7A also have moderately high measures of evenness. The Shannon–Wiener heterogeneity index measures richness and evenness, and also shows that Levels 8 + 8A, 4 + 4A and 3 were the most heterogeneous, followed by Levels 5 and 7 + 7A, with Level 6 as the most homogeneous.
Marine and Terrestrial Molluscs
As presented above, tallies of marine shell and land snail at Nugljanska are only available by weight. When we compare the relative abundances of terrestrial mammal bone against weight of land snails and marine shells by level, it is clear that these alternative resources were not a large contributor to the faunal assemblage, based on excavated material. Keeping in mind that bone is more dense and therefore heavier than shell, terrestrial mammal bone by weight makes up 98–100% of the total assemblage, and only in one context (the earliest Holocene, Level 5) do land snail and marine shell add up to over 1% of the assemblage when combined (). In order to see past the potential bias of bone weight, it is useful to consider land snail in contrast to marine mollusc abundance. If marine and terrestrial mollusc data are directly compared, both are relatively scarce and may be incidental in the Pleistocene assemblage. Land snails dominate the assemblage in Holocene Level 5 at the site (). Only in Levels 4 + 4A and 3 does marine shell constitute the majority of the shell assemblage ().
Figure 7. Relative abundances of marine and terrestrial mollusc shells at Nugljanska.
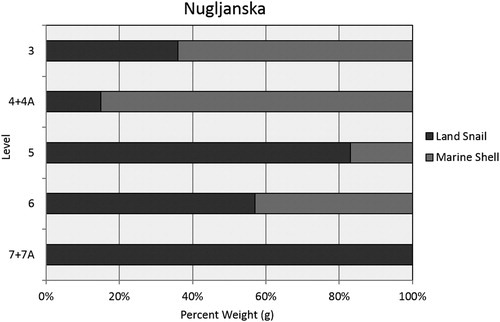
Table 4. Relative abundances of terrestrial mammals and terrestrial and marine molluscs by weight at Nugljanska
Table 5. Relative abundances of marine and terrestrial molluscs by weight within each level at Nugljanska
Discussion
Taphonomy
The characteristics of the assemblage at Nugljanska suggest that it was primarily accumulated by humans, based on the presence of cut marks and burnt bone. The amount and degree of weathered bone suggests that accumulation was attritional, and that most bones probably remained on the surface for a short time before deposition. The percentage of bone that had been gnawed by carnivores was relatively high and could be a function of use of the cave by carnivores during periods when humans were not using the site. Carnivores may have introduced some bone into the assemblages, as well as destroyed or fragmented remains that were left on the surface of the cave floor. It is interesting to note that the extent of carnivore gnawing is lower when indicators of human influence, i.e. burning and cut marks, are more frequent, particularly in Level 4 + 4A. The intensity of burning varies, but the majority appears to be low level, suggesting some degree of cooking, which could include stewing or roasting. The combination of high levels of weathered bone and carnivore activity suggests that site use may not have been that intensive in levels where these occur in high frequencies, but burnt and cut marked bone in all levels testifies to the use of the site and the contribution to the faunal assemblage by humans.
Taxonomic Composition of the Faunal Assemblages
In the late Pleistocene at Nugljanska, there was a focus on large, energy expensive game such as red deer and aurochs. Roe deer and boar supplemented this subsistence economy. In the early Holocene, there was a shift to primary exploitation of boar, supplemented by cervids and bovids. The shift in the representation of red deer (Cervus elaphus) and boar (Sus scrofa) in the late Pleistocene and early Holocene at Nugljanska is immediately noticeable and provides an interesting discussion point. Radiocarbon dates for Levels 5 and 6 suggest a break in the sequence of almost 3500 years. The significant environmental and ecological changes in the area may have resulted in habitat preferred by boar. The representation of boar in the assemblage may also be a result of the nature of seasonal site use at this time. If red deer were seasonally migrating long distances and so were available during only part of the year in the Late Upper Palaeolithic, Nugljanska may have been used during the time period represented by Level 6 for the specific targeting of red deer. With changes in regional ecology occurring during the early Holocene, human groups may have used the site during multiple seasons of the year, relying on locally abundant, non-migratory game such as wild boar and roe deer to supplement the red deer hunting taking place in another season. A detailed discussion of long bone fusion data, tooth eruption and wear patterns, and stable isotope analysis of teeth that contribute to the discussion of sesasonality are outside the scope of this paper (CitationPilaar Birch 2012). It is also noted that large carnivores such as Ursus are very rare and may suggest human curation or incidental deposition rather than occupation of the cave by this species.
Marine resources would have also played a role in dietary practices at Nugljanska, although their overall contribution to the diet is debateable. Nugljanska was approximately 150 km from the coast at the time of lowest sea level during the last glacial maximum around 20,000 years ago, and reached its present day distance of approximately 20 km by about 8000 years ago (Levels 3 and 4). It would have been higher in elevation relative to the sea, and also would have been surrounded by different habitats, hosting different relative abundances of fauna. The availability of various fauna would have been seasonally determined, and depending on when and how often Nugljanska was used, marine or freshwater resources may have been utilised in differing intensities at the site. No fish remains have been documented from any layer of the site. Recovered microfaunal remains have not been systematically analysed, but it is unlikely that marine fish contribute any substantial proportion based on preliminary study carried out by other researchers. When found, specimens of the gastropod Columbella are perforated, suggesting modification for decorative purposes, comparable to those found at Pupićina and other contemporaneous Mesolithic sites. Cerastoderma and Mytilus, both of which are easy-to-gather intertidal species, were most likely collected for consumption. The significance of fluctuating abundances of land snails at any cave site is debatable. Snails are natural cave occupants and so their use and/or consumption by humans, while likely, are difficult to characterise. However, Miracle (Citation2001, Citation2002) has shown that at Mesolithic Pupićina, abundances of snails recovered co-varied with abundances of other edible mammalian taxa, rather than with sample size or excavated area, and they appear to play some role in the diet at Nugljanska throughout the Holocene.
Measures of Diversity
Species richness is not an effect of sample size and there is a real trend of introduction of additional taxa into the faunal assemblage through time. This could be due to human use of these taxa. It is possible these species were hunted for secondary purposes, such as fur, in addition to being consumed, although no cut marks were observed on these specimens. Their appearance could also be unrelated to human activity; they could have accumulated as a result of a change in local environmental conditions which made them more abundant and more likely to end up in the cave as a result of small carnivores or burrowing. For example, the potential extra-dietary species in Level 8 + 8A include small numbers of hare and beaver, while the additional species in Levels 4 + 4A and 3 are badger, hare, fox and marten. When diversity indices were calculated, Level 6 was found to be the most homogeneous, and Levels 8 + 8A, 4 + 4A and 3 were found to be the most heterogeneous. Level 8 + 8A dates to the Bølling-Allerød, a period that is considered to be a climatic optimum but also the time of most rapid change following deglaciation. During this time, the seasonal availability of large ungulates such as red deer may have been unpredictable and smaller bodied forest species may have been more locally abundant. Level 6 falls at the end of the Bølling-Allerød and the beginning of the Younger Dryas (12,845–13,225 cal BP), so the specialised and relatively more homogeneous faunal assemblage are more difficult to correlate with environment. The assemblages in Levels 4 + 4A and 3 are taxonomically rich. Although boar are the most abundant taxa in the Holocene, the remaining ungulate prey species are also more evenly distributed in Levels 4 + 4A and 3 than in previous levels. Level 4 + 4A dates to between 9316 and 8977 cal BP, during the Boreal period, and it is estimated that Level 3 also dates within this range. This is suggestive that dietary diversification may be linked with not only specific habitat factors at this site that influence prey choice (i.e. dominance of boar), but also wider regional environmental changes which may have influenced mobility patterns, leading to a broad spectrum diet. This includes the addition of some terrestrial and rare marine molluscs to the diet.
The Data in Regional Context
It is often the case that the combination of multiple lines of evidence tends to complicate the interpretation, but this may mean that it is closer to the complex reality. Faunal evidence from Nugljanska generally indicates a more heterogeneous use of resources in the early Holocene than in the Pleistocene. This accompanies a switch of dominance from red deer to wild boar. The scale of the increase of boar in the Holocene compared to the Pleistocene at Nugljanska probably reflects a local habitat change which resulted in increased abundances of boar. The additional species which enter the faunal assemblage at this time may have been used for food, but could have also been used for fur or are incidental. The abundance of land snails peaks in the initial Holocene, while marine molluscs are always scarce but appear in increased abundance throughout the Holocene sequence. Overall, the available data support the interpretation of dietary diversification in the Holocene. It is useful to compare these results with some of the sites in the region for which data are available.
At the nearby site of Vela Špilja Lošinj there is a clear difference in pattern between the specialised Late Upper Palaeolithic (Pleistocene) and diverse Mesolithic (Holocene) horizons, though this may be complicated by small sample size (Pilaar Birch and Miracle in press). Abundances of land snails and marine molluscs increase dramatically in the Holocene. In comparison, the Neolithic suggests a complete economic switch to a dependence on domestic sheep and goat, though the incidence of marine molluscs also increased in this horizon. Broad interpretation of seasonal cull patterns may also suggest a shift from autumn use in the Late Upper Paleolithic to spring use, or use in both seasons, in the Mesolithic (Pilaar Birch and Miracle in press).
Miracle (Citation1997) had shown that for much of the Late Upper Palaeolithic and Mesolithic at nearby Pupićina, red deer were the main targeted prey species, but that roe deer and boar were always important. There is a gradual inclusion of additional resources, notably land snails, though marine molluscs also contributed in small amounts. The subtle shifts in relative abundances of taxa in subsequent horizons may have to do with the intensity and continuity in use of this site through time. Use in the Late Upper Palaeolithic tended to be focused on autumn procurement of resources. Long bone fusion data suggest that this pattern continued in the Mesolithic, and seems to have included winter and summer use, and stable isotope analysis of marine shell may add to the evidence of seasonal use (Pilaar Birch 2012). As pertains to mobility in the region, Miracle had considered the fact that sea level rise meant the coast became more accessible to human groups throughout the Pleistocene–Holocene transition. From the Mesolithic the distance to the coast may have been as close as 20 km, which Miracle suggested (based on estimations in Kelly Citation1995) was beyond a logistical foray of 1 day, and that marine resources were brought to Pupićina through residential moves, multi-day logistical forays or trade with other groups. An additional important note about marine molluscs at Pupićina is that Mytilus, which are the most abundant taxa, store particularly well and are able to remain alive for a few days after collection. These may have been targeted in preference to topshells and other species which die quickly. This in turn has implications for the use of mussels as a supplementary resource as groups arrived at Pupićina from the coast (Miracle Citation1997). He used patterns of diversification and the inclusion of mollusc resources as evidence for increasing residential mobility during the early Holocene as opposed to the Late Upper Palaeolithic. This reasoning may support site use in multiple seasons, as people made more frequent residential moves throughout the year within a given resource catchment area, returning to sites in different seasons to target different resources. In contrast, he suggests single-season site use of longer duration may have included more logistical forays rather than residential moves, and acquiring large ungulates which were particularly abundant at a certain time of year within a site exploitation territory may have provided enough meat to last a whole season before moving on.
At the rockshelter of Šebrn Abri there seems to have been a generalised diet in the earliest Mesolithic, with increasing specialisation in red deer predation (up to about 34% of the assemblage) through time (Miracle et al. Citation2000). Roe deer makes up about 13% of the assemblage, and wild boar comprises about 11%. These species, combined with the body-size categories of medium and small ungulate, make up 90% of the total assemblage. Although red deer never comprise the majority of the assemblage, the main pattern which emerges from the data is increasing specialisation at Šebrn, in contrast to diversification at the case-study site. The main cause for this could be the use of Šebrn as a place for obtaining hinterland forest species at only certain parts of the year.
As an outgroup comparison, Vela Spila on Korčula, which is argued to have been a glacial refugium for plant and animal species (Spry-Marqués Citation2012), there is also a trend of resource diversification in the Mesolithic. The Late Upper Palaeolithic assemblage reflects specialised exploitation of red deer and Equus hydruntinus, plentiful on the Great Adriatic Plain. With sea level rise isolating Vela Spila on the island of Korčula, fine bone tools, interpreted as being used for eating shellfish (Čečuk and Radić Citation2005) are found in abundance. Tens of thousands of shells (Osilinus and Patella spp.) were recovered from Mesolithic layers at the site, and were accompanied by a broad mammalian spectrum including boar, hare and small numbers of red deer. The degree of intensity of occupation at this southern site suggests that marine resources were more important than they ever were in Istria, which maintained a focus on terrestrial species within the pattern of diversification. Furthermore, parallels may be drawn between Vela Spila and the sites of Franchthi in the Peloponnese and Grotta dell'Uzzo on Sicily, which also suggest a heavy reliance on marine resources through time as a result of rising sea level.
I would like to further consider the continuing importance of terrestrial resources at Nugljanska and on the Istrian Peninsula as a whole, because the exploitation of shellfish appears to be lower than might expected as a result of sea level rise. Stable isotope analyses of δ13C and δ15N from archaeological human and animal bone collagen by other researchers have contributed further evidence for dietary diversification in the region. Stable isotope results from Pupićina, Šebrn and Klanjčeva caves on economic species such as boar, red deer, roe deer, hare and chamois/ibex suggested these animals were procured from the same geographic area and were part of the same ecosystem (Robinson Citation2006). The results showed little variation between upland and lowland sites and between potentially migratory and non-migratory species in δ13C and δ15N, suggesting upland and lowland subsistence practices were not differentiated. A more detailed consideration by Paine et al. (Citation2009) included human samples (n = 4) from Pupićina and found that Mesolithic people were deriving most of their protein from terrestrial resources. She suggested that people were utilising hinterland terrestrial resources while at Pupićina, and using coastal or aquatic resources only as a supplement. Lightfoot et al. (Citation2011) presented a much larger sample size and provide regional data on dietary change for the Mesolithic–Neolithic transition along the eastern Adriatic coast, including Istria. Their data suggest that coastal Mesolithic populations in Croatia, including people buried at Pupićina, had greater marine resource intake compared to inland sites, though amounts appear to be lower than at other Western European Mesolithic sites. The slightly higher δ15N signal in humans suggests that while Pupićina itself may have been used as a center for autumn hunting of large/medium ungulates supplemented by smaller game (Miracle Citation2001, Citation2002), it was part of a wider, broad spectrum economy in which Mesolithic people were exploiting predictable, available resources specific to local habitats and ecologies, including marine resources at nearby coastal sites which may now be lost.
Conclusion
The continuing reliance on terrestrial resources at Nugljanska through time suggests an extensive use of the hinterland of Istria by human groups. It suggests that further excavation and study of material from Nugljanska may prove fruitful for making interpretations about the seasonal nature of site use and the interplay between subsistence and mobility strategies at the Pleistocene–Holocene transition. The subtle trend of diversification at Nugljanska may have more to do with local environmental changes and changing seasonal mobility patterns than the ‘resource stress’ often suggested as an explanation for such data.
Acknowledgements
We would like to thank Darko Komšo and the Arheološki Muzej Istre for providing access to excavated material. We also thank the Oxford Radiocarbon Accelerator Unit and the Natural Environment Research Council UK (NERC) for providing radiocarbon dates. S.E.P.B. would also like to thank the Gates Cambridge Trust and the Division of Archaeology, Department of Archaeology and Anthropology, University of Cambridge, for providing additional research funding during her PhD, from which these data are derived.
References
- Andrič, M., Kroflič, B., Toman, M. J., Ogrinc, N., Dolenec, T., Dobnikar, M. and Čermelj, B. 2008. Late quaternary vegetation and hydrological change at Ljubljansko barje (Slovenia). Palaeogeography, Palaeoclimatology, Palaeoecology 270, 150–65.
- Barfield, L. H. 1999. The Moser collection in the Naturhistorisches Museum, Vienna. Atti della Società per la Preistoria e Protostoria della Regione Friuli-Venezia Giulia XI, 19–62.
- Boschian, G. and Fusco, F. 2007. Figuring out no-one's land: why was the karst deserted in the Late Glacial?, pp. 15–26 in Whallon, R. (ed.), Late Paleolithic Environments and Cultural Relations Around the Adriatic. British Archaeological Reports International Series 1716. Oxford: Archaeopress.
- Brambati, A. 1992. Origin and evolution of the Adriatic Sea, pp. 327–46 in Colombo, G., Ferrari, I., Ceccherelli, V. U. and Rossi, R. (eds.), Marine Eutrophication and Population Dynamics. Fredensborg: Olsen and Olsen.
- Čečuk, B. and Radić, D. 2005. Vela Spila. Vela Luka: Centar za kulturu ‘Vela Luka’.
- Favaretto, S., Asioli, A., Miola, A. and Piva, A. 2008. Preboreal climatic oscillations recorded by pollen and foraminifera in the southern Adriatic Sea. Quaternary International 190, 89–102.
- Ferretti, M. E., Moretti, D., Savelli, A., Stefanon, M., Tramontana, M. and Wezel, F.-C. 1986. Late Quaternary alluvial sequences in the north-western Adriatic Sea from UNIBOOM profiles. Bollettino di Oceanologia Teorica ed Applicata 4, 63–72.
- Gallizia Vuerich, L. and Princivalle, F. 1994. Studio mineralogico e palinologico su alcuni sediment della grotto dell'Edera (Carso Triestino): un tentative di ricostruzione paleoclimatica. Il Quaternario 7, 569–76.
- Hammer, Ø., Harper, D. A. T. and Ryan, P. D. 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4, 9. Available at: http://palaeo-electronica.org/2001_1/past/issue1_01.htm.
- Kelly, R. L. 1983. Hunter-gatherer mobility strategies. Journal of Anthropological Research 39, 277–306.
- Kelly, R. L. 1995. The Foraging Spectrum: Diversity in Hunter-Gatherer Lifeways. Washington, DC: Smithsonian Institution Press.
- Komšo, D. 2006. The Mesolithic in Croatia. Opuscula Archaeologica 30, 55–92.
- Komšo, D. and Pellegatti, P. 2007. The Late Epigravettian in Istria: Late Paleolithic colonization and lithic technology in the northern Adriatic area, ppp. 27–39 in Whallon, R. (ed.), Late Paleolithic Environments and Cultural Relations Around the Adriatic. British Archaeological Reports International Series 1716. Oxford: Archaeopress.
- Lambeck, K., Antonioli, F., Purcell, A. and Silenzi, S. 2004. Sea-level change along the Italian coast for the past 10,000yr. Quaternary Science Reviews 23, 1567–98.
- Lightfoot, E., Boneva, B., Miracle, P. T., Šlaus, M. and O'Connell, T. C. 2011. Exploring the Mesolithic and Neolithic transition in Croatia through isotopic investigations. Antiquity 85, 73–86.
- Miracle, P. 2001. Feast or famine? Epi-Paleolithic subsistence in the northern Adriatic basin. Documenta Praehistorica 26, 177–97.
- Miracle, P. 2007. The Late Glacial ‘Great Adriatic Plain’: ‘Garden of Eden’ or ‘No Man's Land’ during the Epipaleolithic? A view from Istria (Croatia), pp. 41–51 in Whallon, R. (ed.), Late Paleolithic Environments and Cultural Relations Around the Adriatic. British Archaeological Reports International Series 1716. Oxford: Archaeopress.
- Miracle, P., Galanidou, N. and Forenbaher, S. 2000. Pioneers in the hills: early Mesolithic foragers at Šebrn Abri (Istria, Croatia). European Journal of Archaeology 3, 293–329.
- Miracle, P. T. 1997. Early Holocene foragers in the karst of northern Istria. Poročilo o raziskovanju paleolitika, neolitika in eneolitika v Sloveniji 24, 43–61.
- Miracle, P. T. 2002. Mesolithic meals from Mesolithic middens, pp. 65–88 in Miracle, P. T. and Milner, N. (eds.), Consuming Passions and Patterns of Consumption. Cambridge: McDonald Institute for Archaeological Research.
- Miracle, P. T. and Forenbaher, S. 2000. Pupićina Cave Project: brief summary of the 1998 season. Histria Archaeologica 29, 27–48.
- Miracle, P. T. and Pugsley, L. 2006. Vertebrate faunal remains from Pupićina Cave, pp. 259–399 in Miracle, P. T. and Forenbaher, S. (eds.), Prehistoric Herders of Northern Istria. The Archaeology of Pupićina Cave. Volume 1. Pula: Arheološki Muzej Istre.
- Mussi, M. 2001. Earliest Italy: An Overview of the Italian Paleolithic and Mesolithic. New York: Springer.
- Paine, C., O'Connell, T. and Miracle, P. T. 2009. Stable isotopic reconstruction of diet at Pupićina Cave, pp. 210–6 in McCartan, S., Schulting, R., Warren, G. and Woodman, P. (eds.), Mesolithic Horizons. Papers Presented at the Seventh International Conference on the Mesolithic in Europe, Belfast 2005. Oxford: Oxbow.
- Pilaar Birch, S. E. 2012. Human Adaptations to Climate Change and Sea Level Rise at the Pleistocene-Holocene Transition in the Northeastern Adriatic. Unpublished PhD dissertation, Department of Archaeology, University of Cambridge.
- Pilaar Birch, S. E. and Miracle, P. T. In press. Human response to climate change in the Northern Adriatic during the late Pleistocene and Early Holocene in Monks, G.(ed.), Climate Change and Past Human Responses: An Archaeozoological Perspective. Vertebrate Paleobiology and Paleoanthropology Series. New York: Springer.
- Robinson, S. 2006. Using carbon and nitrogen stable isotopic analysis to reconstruct the food web of the peninsula of Istria, northern Croatia. Unpublished MPhil dissertation, Department of Archaeology, University of Cambridge.
- Rossignol-Strick, M. 1999. The Holocene climatic optimum and pollen records of sapropel 1 in the eastern Mediterranean, 9000–6000 BP. Quaternary Science Reviews 18, 515–30.
- Schmidt, R., Müller, J., Drescher-Schneider, R., Krisai, R., Szeroczynska, K. and Barić, A. 2000. Changes in lake level and trophy at Lake Vrana, a large karstic lake on the island of Cres (Croatia), with respect to palaeoclimate and anthropogenic impacts during the last approx. 16,000 years. Journal of Limnology 59, 113–30.
- Spry-Marqués, V. P. 2012. The Adriatic Plain: a glacial refugium? Epigravettian subsistence strategies at Vela Spila (Croatia). Unpublished PhD dissertation, Department of Archaeology and Anthropology, University of Cambridge.
- Surić, M. 2005. Submerged karst–dead or alive? Example from the eastern Adriatic coast (Croatia). Geoadria 10, 5–19.
- van Andel, T. H. 1989. Late Quaternary sea level change and archaeology. Antiquity 63, 733–45.
- Voytek, B. 2011. In the shadow of the grand narrative: revisiting the early Holocene of the northern Adriatic. Journal of World Prehistory 24, 189–99.
- Willis, K. J. 1994. The vegetational history of the Balkans. Quaternary Science Reviews 13, 769–88.
- Zavatarelli, M., Baretta, J. W., Baretta-Bekka, J. G. and Pinardi, N. 2000. The dynamics of the Adriatic Sea ecosystem: an idealized model study. Deep-Sea Research 47, 937–97.