Abstract
Background
Deep oceans have been suggested as a possible site where the origin of life occurred. Along with this theoretical lineage, experiments using components from deep ocean water to recreate life is underway. Here, we propose that if terrestrial organisms indeed evolved from deep oceans, supply of deep ocean mineral water (DOM) to humans, as a land creature, may replenish loss of molecular complexity associated with evolutionary sea-to-land migration.
Methods
We conducted a randomized, double-blind, placebo-controlled crossover human study to evaluate the effect of DOM, taken from a depth of 662 meters off the coast of Hualien, Taiwan, on time of recovery from a fatiguing exercise conducted at 30°C.
Results
The fatiguing exercise protocol caused a protracted reduction in aerobic power (reduced VO2max) for 48 h. However, DOM supplementation resulted in complete recovery of aerobic power within 4 h (P < 0.05). Muscle power was also elevated above placebo levels within 24 h of recovery (P < 0.05). Increased circulating creatine kinase (CK) and myoglobin, indicatives of exercise-induced muscle damage, were completely eliminated by DOM (P < 0.05) in parallel with attenuated oxidative damage (P < 0.05).
Conclusion
Our results provide compelling evidence that DOM contains soluble elements, which can increase human recovery following an exhaustive physical challenge.
Electronic supplementary material
The online version of this article (doi:10.1186/1550-2783-10-7) contains supplementary material, which is available to authorized users.
Chih-Yang Huang and Chia-Hua Kuo contributed equally to this work.
Copyright comment
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
A living organism can be regarded as a gathering of diverse molecules originating from the earth that works cooperatively to decrease entropy against the catabolic stresses from an ever-changing environment. Deep ocean mineral water (DOM) has been suggested to contain the primordial source of chemical components contributing to the creation of life [Citation1, Citation2]. Besides the major minerals, more than 70 trace elements existing in the ocean water have been documented [Citation3]. The question regarding how many chemical components are necessary or required to support the best complexity of human life is not completely defined.
Presently, there is no information as to the effect of DOM on the physiological function of animals or humans following extreme environmental or physiological challenges. The most consistent observations reside around the anti-atherogenic effects of DOM against dietary challenges [Citation4–Citation7]. Compared to desalinated surface ocean water with a similar profile of major minerals (magnesium, potassium, calcium, sodium, chloride, and sulfate ions), desalinated DOM has been found to have a much superior effect on preventing the development of atherosclerosis in rabbits challenged with a high cholesterol diet [Citation4]. This result suggests that the highly diverse trace elements found in DOM are responsible for its anti-atherogenic capabilities and have significant physiological effects on terrestrial animals. It is possible the surface waters of the oceans where sunlight is permeable are devoid of these important trace elements as a result of the photosynthetic activity of many marine organisms [Citation8].
Due to environmental limitations marine and terrestrial organisms rely on different nutritive sources to maintain life [Citation9]. Paleobiological evidence, however, strongly suggests terrestrial life evolved from marine ancestor [Citation10]. Although sharing common cellular constituents with marine organisms, terrestrial survivors had to acquire alternative nutritive sources from the land to compensate for the loss associated with ancient sea-to-land migration. We proposed that if deep oceans contain the evolutionary preferred constituents for terrestrial descendents, DOM supplementation can be complementary to achieve the best biological complexity for land animals. To test this hypothesis, we conducted a human study in which we determined the time required for physical performance to recover after a dehydrating exercise when desalinated DOM or placebo drink was supplied for rehydration.
Methods
Subjects
Subjects taking alcohol, medication, or nutritional supplements were excluded from the study. Twelve healthy male volunteers (age 24 ± 0.8 y; height 171.8 ±1.5 cm; weight 68.2 ±2.3 kg; VO2max 49.7 ± 2.2 ml · kg−1 · min−1) were enrolled as participants in the study. Baseline VO2max were measured 72 h before the beginning of the study. Written informed consent was obtained after explanation of the purpose and experimental procedures of the study. This study was approved by the appropriate university Institutional Review Boards and performed in accordance with principles of the Declaration of Helsinki.
Drink
The desalinated DOM, taken from the West Pacific Ocean (662 meters in depth), was kindly provided by Taiwan Yes Deep Ocean Water Co., Ltd. (Hualien, Taiwan). DOM was filtered by a micro-filter (removal of microorganism) and an ultra-filter (removal of macromolecule and virus) before use. Molecules sized above 1.5 KD were removed after the two filtration procedures. To mask the taste difference between DOM and placebo, the same amount of sucrose, artificial flavors, citrate, citrus juice, calcium lactate, potassium chloride, vitamin C, and mixed amino acids was added to each. Tap water purified by reverse osmosis process was used for making the placebo drink.
Experimental design
An exercise-challenge protocol used by Nose et al. was modified for this study [Citation11]. Subjects were required to run on a motorized treadmill at 40% VO2max at a room temperature of 30°C until a body mass decline of 3% (maximal running time: 240 min). During recovery, subjects consumed pure water or DOM containing the ingredients listed above at an amount equivalent to 1.5 fold of their body mass loss [Citation12]. Water supplements were evenly divided into 4 sub-supplements and ingested at 30-minute intervals. Measures of physical performance (aerobic power and lower-body muscle power), physiological stress, and muscle damage were determined 4, 24, and 48 h during the recovery period. To control for possible confounding effects of individual variation, a randomized double-blind crossover design was employed with trials spaced 7 d apart.
Physical performance
Aerobic power (maximal oxygen consumption, VO2max) and peak lower-body muscle power were the physical performance measures selected for determining the degree of physical fatigue recovery. VO2max was evaluated by the Bruce graded treadmill running protocol. This protocol consists of a 5-min warm up and incremental increases in speed and grade every 3 min until exhaustion. Verification that VO2max was achieved was a Respiratory Exchange Ratio (RER) greater than 1.1 and a plateau in VO2 with increasing workload. Samples of expired gases were analyzed using a MetaMax3B (Cortex Biophysik, Nonnenstrasse, Leipzing, Germany). Peak lower-body muscle power was assessed using a Bertec force plate (4060-NC2000, Bertec Corporation, Columbus, Ohio, USA) with a sampling rate of 1,000 Hz. Each subject performed 3 repetitions of maximal squat jumps from a 90° knee flexion angle to full extension. Subjects were signaled when to jump by a light placed 2 meters in front of them at eye level. There was a one-minute rest between jumps. Velocity and power of each jump was calculated from vertical ground reaction forces (VGRF) according to the impulse-momentum theorem (VGRF × time = body mass times ΔV, ΔV is the change in vertical velocity) (Innovative Sports Training, Inc, Chicago, IL, USA). Instantaneous velocity was determined by adding ΔV to the previous time interval, starting at zero at the beginning of the jump. Instantaneous power was derived from the product of VGRF measured by the force plate and the calculated instantaneous velocity [Citation13]. The peak value of instantaneous power during the entire period of each jump was selected as peak power. The peak power values of the 3 jumps were averaged for statistical calculation.
Biochemical analysis
Venous blood samples were assayed for plasma myoglobin (Immunology Consultants Laboratory, Inc. OR, USA), thiobarbituric acid reactive substances (TBARS) (Cayman Chemical Company, Ann Arbor, MI, USA), cortisol (IBL-America, Inc. MN, USA), erythropoietin (eBioscience, Vienna, Austria), IL-6 (eBioscience, Vienna, Austria), and testosterone (Nova Tec Immundiagnostica GmbH, Dietzenbach, Germany) with enzyme-linked immunosorbent (ELISA) readers (Tecan Genios, Salzburg, Austria). Plama CK was analyzed enzymatically using a bench top DT-60II analyzer (Johnson and Johnson, NY, USA).
Statistical analyses
All values are expressed as percent of baseline (mean ± standard error). A two-way analysis of variance with repeated measures was used for comparisons between DOM and pure water at specified time points during recovery. A paired t test with Bonferroni’s correction was used to compare treatment differences at each time point. Probability of a type I error less than 5% was considered statistically significant.
Results
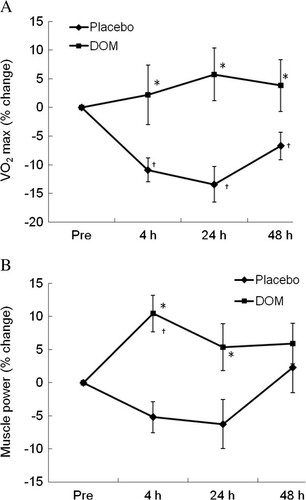
The geographic location of DOM is illustrated in Figure . Concentrations of the minerals and trace elements of DOM are shown in Table . Our physical challenge protocol successfully induced a prolonged physical fatigue in aerobic power of our control trial (RO purified water) for 48 h of recovery (Figure A, P < 0.05). DOM supplementation completely restored the loss of aerobic power to baseline within 4 h. Lower-body muscle power was not affected by our physical challenge protocol, yet DOM supplementation increased the power performance by ~10% above baseline (Figure B) at 4 h and 24 h during the recovery (P < 0.05).
Figure 1 Geographic location of DOM collection. The black square designates the site of seawater collection, providing the shortest piping distance from land down to the deep site of the ocean (a depth of 662 meters off the coast of Hualien, Taiwan) along the circum-Pacific belt (known as Pacific Ring of Fire) in East Asia.
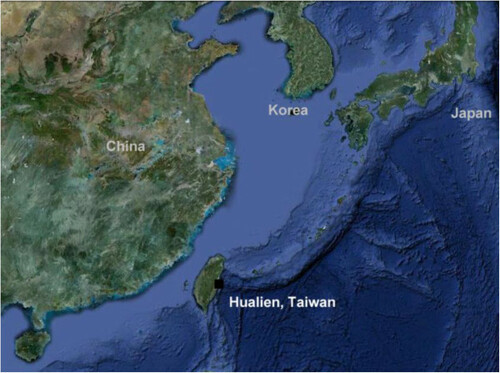
Table 1 Minerals and trace elements in deep ocean mineral water (DOM) drink
Figure 2 Human physical performance. DOM accelerated the recovery of aerobic capacity after a fatiguing exercise (A), and increased lower-body muscle power performance (B) during recovery. *significance against Placebo, P < 0.05; †significance against Pre, P < 0.05. N. D.: non-detectable.
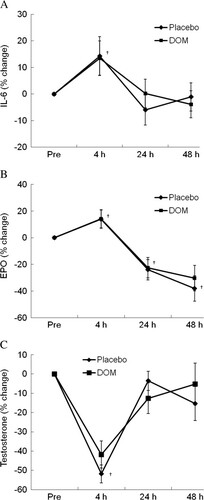
Stress hormone responses are shown in Figure and confirms the same physiological stress produced during each trial. For both control and DOM trials, the exercise challenge temporally elevated plasma IL-6 levels (14%, P < 0.05) at 4 h of recovery to a comparable extent (Figure B). This increase subsided to baseline within 24 h. Similarly, we observed a rise in erythropoietin (EPO) of 14% (P < 0.05) at 4 h of recovery for both treatments. By 24 h of recovery, however, EPO had fallen below baseline and was still below baseline at 48 h of recovery (P < 0.05). Both cortisol and testosterone dropped at 4 h during recovery (by 46% and 52%, P < 0.05), and had returned close to baseline by 24 h and 48 h following exercise. Again, there was no treatment differences associated with these hormones.
Figure 3 Stress hormones. Exercise challenge elevated plasma IL-6 (A) and EPO levels (B, P < 0.05) for both trials to a similar extent. Testosterone dropped on both trials during recovery (C, P < 0.05), and returned to baseline by 24 h during recovery. No group differences in stress hormone responses were found after the physical challenge. †significance against Pre, P < 0.05.
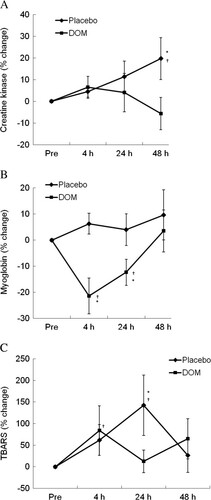
Plasma CK and myoglobin, known as exercise-induced muscle damage markers [Citation14], are shown in Figure . A gradual rise in CK was observed 48 h following exercise in the control trial (Figure A), while DOM eliminated this increase (P < 0.05). A marginal increase in myoglobin was observed at 4 h and 24 h following exercise in the control trial, while following the DOM treatment myoglobin was significantly below the control level at 4 and 24 h of recovery (Figure B). Results for the oxidative marker thiobarbituric acid reactive substances (TBARS) are shown in Figure C. TBARS increased significantly during the control trial at 4 h and 24 h of recovery (P < 0.05), while increasing only at 4 h of recovery during the DOM trial.
Figure 4 Muscle damage markers. Exercise-induced muscle damage was suppressed by DOM, as indicated by attenuated CK (A) and myoglobin (B) responses during recovery. DOM also attenuated oxidative damage (TBARS) increased by exercise (C). *significance against Placebo, P < 0.05; †significance against Pre, P < 0.05.
Discussion
In this study, we propose that if terrestrial organisms evolved from deep ocean [Citation10], supply of deep ocean mineral water (DOM) to humans may replenish loss of molecular complexity associated with evolutionary sea-to-land migration, and optimizes the biological fitness. Here, we provide evidence that desalinated DOM, taken from 662 meters below sea-level, can substantially accelerate recovery from physical fatigue in aerobic power and enhance lower-body muscle power after a prolonged bout of dehydrating exercise. This improvement appears to be associated with a complete elimination of exercise-induced muscle damage, suggesting that DOM contains components, which can complement and enhance the molecular and cellular complexity of humans to minimize entropic stress produced during prolonged physical activity in the heat.
The key components of DOM contributing to the observed ergogenic benefits are not exactly known. In the study, the DOM taken from the west rim of the Pacific Ocean is characterized by enriched contents of boron, magnesium, lithium, and rubidium. In DOM the content of boron (1.59 mg/L), which is now considered an essential nutrient for humans, is 5–10 fold that found in human serum (~0.2-0.3 mg/L) [Citation15]. Boron is known to attenuate exercise-induced rise in plasma lactate in animals [Citation16] and to prevent magnesium loss in humans [Citation17]. Serum magnesium concentration and dietary magnesium intake are known correlates of muscle strength [Citation18, Citation19]. Therefore, the minerals and trace elements in DOM may work cooperatively to sustain normal human performance.
The observed effect of DOM on accelerating fatigue recovery is closely associated with the eradication of exercise-induced muscle damage [Citation20, Citation21]. Elevation of these muscle damage markers normally occurs in parallel with increased oxidative damage [Citation22]. Our results on thiobarbituric acid reactive substances (TBARS) fits well with those on markers of muscle damage (P < 0.05). Higher content of magnesium, lithium, and rubidium in DOM may be associated with strengthened antioxidant capability against oxidative stress during post-exercise recovery [Citation23–Citation25]. In animals, lack of magnesium in their diet leads to increased free radical production [Citation26], while magnesium supplementation eliminates free radical production induced by ischemia reperfusion [Citation23] and alcohol drinking [Citation27]. Lithium can increase the free radical scavenging capability in animals [Citation25] and thus help to increase the resilience of a cell against destructive free radical attack [Citation28].
One significant feature of DOM is the enriched rubidium content compared to fresh water. Rubidium concentration increases considerably in seawater as the depth of the ocean approaches 450 meters. The concentration of this trace element in human plasma ranges from 40–310 μg/L [Citation29], about 2.5-20 fold higher than that found in DOM. However, rubidium has a high retention rate in the human body, taking 39-134 days for 50% of infused rubidium to be excreted into urine and feces [Citation30]. Compared to rats fed rubidium, rats fed a rubidium-free diet exhibit higher urea nitrogen in plasma [Citation31], suggesting that rubidium is essential to preserve biological integrity against daily entropic stress. The rubidium concentration in the human brain decreases with age [Citation32], and supplementation of rubidium chloride has been found to increase spontaneous physical activity in animals [Citation33]. Additions of lithium and rubidium into seawater have been shown to increase frequency of movement in jellyfish [Citation34]. The recommended dietary allowance for rubidium has not yet been defined for humans. Rubidium demonstrates interchangeability with potassium in a variety of biological systems meaning that rubidium deficiency can be compensated by supplementation of potassium in many species [Citation35]. Compared to potassium, rubidium may be an evolutionary preferred nutritive source for animals.
The oceans are the largest water reservoirs on earth, which consists of a great diversity of water-soluble chemical components, feeding a vast quantity of marine organisms [Citation8, Citation36]. However, nutrients in the clear ocean surface water have most likely been exhausted by a high rate of photosynthesis [Citation8, Citation37]. Compared to the surface layer of the oceans, DOM may exert greater metabolic benefit, evidenced by its superior action on eliminating oxidative stress and preventing vascular damage in terrestrial animals challenged with a high cholesterol diet [Citation4]. This observation implies that the water-soluble components unique to (or enriched in) DOM may play an important role in supporting metabolic functions of terrestrial animals when they are faced with a various physiological and metabolic challenges.
The limitation of the study is the loci-specific distribution of minerals and trace elements in the ocean, thus preventing us from being able to generalize that DOM from all sites of the world can confer the same ergogenic benefits as presented. Geographic specificity is suggested by a report documenting relatively lower silver, cobalt and nickel concentrations in the North Atlantic Ocean than the other major oceans [Citation38]. Furthermore, the profile of minerals and trace elements is also varied with the depth of the ocean [Citation37, Citation39], and hydrothermal activity and diffusion from bottom sediments can also influence the composition of minerals and trace elements in the ocean waters [Citation40]. Experiments using Antarctic Ocean waters have also suggested that not all deep ocean water will provide comparable biogenic benefits [Citation41].
On the application side, we confirm the benefit of acute DOM supplementation on decreasing physical fatigue with elimination of post-exercise oxidative damage. However, it has been reported a diminished training effect when antioxidant was supplemented to trained men [Citation42], suggesting that free radicals may play a role for training adaptation. Thus, whether or not decreasing oxidative stress by DOM supplementation may confer negative effects on exercise training adaptation demands more investigation.
Conclusion
Our findings demonstrate that desalinated DOM can increase human robustness against an entropic physical challenge, and this positive outcome appears to be associated with its protection against exercise-induced muscle damage. DOM consists of many minerals and trace elements that could not be de novo synthesized by the human body. Thus the momentary imbalance between loss and gain of essential minerals and trace elements after prolonged exercise may underlie the delayed recovery from physical fatigue in humans. In line with the “deep ocean life of origin hypothesis”, the results of this study imply that DOM can provide required nutrients for humans that will speed recovery from entropic physical stress.
Authors’ original file for figure 1
Download TIFF Image (5 MB)Authors’ original file for figure 2
Download TIFF Image (237 KB)Authors’ original file for figure 3
Download TIFF Image (355.3 KB)Authors’ original file for figure 4
Download TIFF Image (350.9 KB)Acknowledgments
This research was partly supported by grants from the Industrial Development Bureau, Ministry of Economic Affairs (grant number 9831101073–6) and National Science Council, Taiwan (grant number 99-2410-H-154-004-MY3).
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
CWH, WHC, YST, CYC, CYH and CHK designed the experiments. CWH and YST performed the experiments. CWH performed the statistical analyses. CWH, JLI, and CHK wrote the manuscript. All authors read and approved the final manuscript.
Related Research Data
References
- MartinW BarossJ KelleyD et al Hydrothermal vents and the origin of life Nat Rev Micro 2008 6 805 814 1:CAS:528:DC%2BD1cXht1CmtrrE
- WhitfieldJ Nascence man Nature 2009 459 316 319 1:CAS:528:DC%2BD1MXmtFKms70%3D https://doi.org/10.1038/459316a 19458689
- FarringtonJW Ocean Studies Board NRC (Series Editor): 50 years of ocean discovery: National Science Foundation 1950–2000 Achievements in chemical oceanography 2000 Washington, D.C. The National Academics Press
- MiyamuraM YoshiokaS HamadaA et al Difference between deep seawater and surface seawater in the preventive effect of atherosclerosis Biol Pharm Bull 2004 27 1784 1787 1:CAS:528:DC%2BD2cXhtFGmtr3O https://doi.org/10.1248/bpb.27.1784 15516723
- FuZY YangFL HsuHW et al Drinking deep seawater decreases serum total and low-density lipoprotein-cholesterol in hypercholesterolemic subjects J Med Food 2012 15 535 541 3359629 1:CAS:528:DC%2BC38Xot1yis7w%3D https://doi.org/10.1089/jmf.2011.2007 22424458
- LeeCL KungYH WangJJ et al Enhanced hypolipidemic effect and safety of red mold dioscorea cultured in deep ocean water J Agric Food Chem 2013 59 8199 8207 https://doi.org/10.1021/jf201948v
- RadhakrishnanG YamamotoM MaedaH et al Intake of dissolved organic matter from deep seawater inhibits atherosclerosis progression Biochem Biophys Res Commun 2009 387 25 30 1:CAS:528:DC%2BD1MXptValt70%3D https://doi.org/10.1016/j.bbrc.2009.06.073 19540194
- OthmerDF RoelsOA Power, fresh water, and food from cold, deep sea water Science 1973 182 121 125 1:STN:280:DC%2BC3cvgs12rsw%3D%3D https://doi.org/10.1126/science.182.4108.121 17777883
- VenturiS Evolutionary significance of iodine Curr Chem Biol 2011 5 155 162 1:CAS:528:DC%2BC3MXht1Cgt73O
- GingerichPD HaqM ZalmoutIS et al Origin of whales from early artiodactyls: hands and feet of Eocene protocetidae from Pakistan Science 2001 293 2239 2242 1:CAS:528:DC%2BD3MXntFCrtr4%3D https://doi.org/10.1126/science.1063902 11567134
- NoseH MackGW ShiXR et al Role of osmolality and plasma volume during rehydration in humans J Appl Physiol 1988 65 325 331 1:STN:280:DyaL1c3pvVGlug%3D%3D 3403476
- ShirreffsSM TaylorAJ LeiperJB et al Post-exercise rehydration in man: effects of volume consumed and drink sodium content Med Sci Sports Exerc 1996 28 1260 1271 1:STN:280:DyaK2s%2Flt1Onuw%3D%3D https://doi.org/10.1097/00005768-199610000-00009 8897383
- WrightGA PustinaAA MikatRP et al Predicting lower body power from vertical jump prediction equations for loaded jump squats at different intensities in men and women J Strength Cond Res 2012 26 648 655 22173625
- SiegelmAJ SilvermanmLM EvansmWJ Elevated skeletal muscle creatine kinase mb isoenzyme levels in marathon runners JAMA 1983 250 2835 2837 https://doi.org/10.1001/jama.1983.03340200069032
- Friis-HansenB AggerbeckB JansenJA Unaffected blood boron levels in newborn infants treated with a boric acid ointment Food Chem Toxicol 1982 20 451 454 1:CAS:528:DyaL3sXisl2jsQ%3D%3D https://doi.org/10.1016/S0278-6915(82)80111-7 6890023
- YaziciZ KayaY BaltaciAK et al The effects of boron administration on plasma leptin and lactate levels in ovariectomized rats which had acute swimming exercise Neuro Endocrinol Lett 2008 29 173 177 1:CAS:528:DC%2BD1cXktFersrw%3D 18283241
- NielsenFH Biochemical and physiologic consequences of boron deprivation in humans Environ Health Perspect 1994 102 59 63 1566640 1:CAS:528:DyaK2MXjsVahtLw%3D https://doi.org/10.1289/ehp.94102s759 7889883
- DominguezLJ BarbagalloM LauretaniF et al Magnesium and muscle performance in older persons: the inchianti study Am J Clin Nutr 2006 84 419 426 2669297 1:CAS:528:DC%2BD28XosVehsLc%3D 16895893
- SantosDA MatiasCN MonteiroCP et al Magnesium intake is associated with strength performance in elite basketball, handball and volleyball players Magnes Res 2011 24 215 219 1:CAS:528:DC%2BC38XhtVOqs7vJ 21983266
- FridenJ LieberRL Structural and mechanical basis of exercise-induced muscle injury Med Sci Sports Exerc 1992 24 521 530 1:STN:280:DyaK383js1emsg%3D%3D https://doi.org/10.1249/00005768-199205000-00005 1569848
- RowlandsDS RosslerK ThorpRM et al Effect of dietary protein content during recovery from high-intensity cycling on subsequent performance and markers of stress, inflammation, and muscle damage in well-trained men Can J Appl Physiol 2008 33 39 51 1:CAS:528:DC%2BD1cXjtlCmt7c%3D
- WangJS HuangYH Effects of exercise intensity on lymphocyte apoptosis induced by oxidative stress in men Eur J Appl Physiol 2005 95 290 297 1:CAS:528:DC%2BD2MXhtFGlt7bL https://doi.org/10.1007/s00421-005-0005-8 16096840
- GarciaLA DeJongSC MartinSM et al Magnesium reduces free radicals in an in vivo coronary occlusion-reperfusion model J Am Coll Cardiol 1998 32 536 539 1:CAS:528:DyaK1cXls1Wisr0%3D https://doi.org/10.1016/S0735-1097(98)00231-9 9708488
- Manuel y KeenoyB MoorkensG VertommenJ et al Magnesium status and parameters of the oxidant-antioxidant balance in patients with chronic fatigue: effects of supplementation with magnesium J Am Coll Nutr 2000 19 374 382 1:CAS:528:DC%2BD3cXksVGgt7o%3D https://doi.org/10.1080/07315724.2000.10718934 10872900
- ShuklaGS Mechanism of lithium action: In vivo and in vitro effects of alkali metals on brain superoxide dismutase Pharmacol Biochem Behav 1987 26 235 240 1:CAS:528:DyaL2sXhs1yqtLw%3D https://doi.org/10.1016/0091-3057(87)90111-0 3033694
- RockE AstierC LabC et al Dietary magnesium deficiency in rats enhances free radical production in skeletal muscle J Nutr 1995 125 1205 1210 1:CAS:528:DyaK2MXlsVejuro%3D 7738680
- Markiewicz-GorkaI ZawadzkiM JanuszewskaL et al Influence of selenium and/or magnesium on alleviation alcohol induced oxidative stress in rats, normalization function of liver and changes in serum lipid parameters Hum Exp Toxicol 2011 30 1811 1827 1:CAS:528:DC%2BC38XosFGmsw%3D%3D https://doi.org/10.1177/0960327111401049 21474619
- AdipudiV ReddyVK Effect of chronic lithium chloride on membrane adenosine triphosphatases in certain postural muscles of rats Eur J Pharmacol 1994 259 7 13 1:CAS:528:DyaK2cXksFSjs7Y%3D https://doi.org/10.1016/0014-2999(94)90150-3 7957596
- SheldonJH RamageH A spectrographic analysis of human tissues Biochem J 1931 25 1608 1627 1260788 1:CAS:528:DyaA38XhtFyrtA%3D%3D https://doi.org/10.1042/bj0251608 16744726
- BurchGE ThreefootSA RayCT The rate of disappearance of rb86 from the plasma, the biologic decay rates of rb86, and the applicability of rb86 as a tracer of potassium in man with and without chronic congestive heart failure J Lab Clin Med 1955 45 371 394 1:CAS:528:DyaG2MXlsVGiug%3D%3D 14354325
- YokoiK KimuraM ItokawaY Effect of low dietary rubidium on plasma biochemical parameters and mineral levels in rats Biol Trace Elem Res 1996 51 199 208 1:CAS:528:DyaK28XhsVKlu70%3D https://doi.org/10.1007/BF02785438 8907022
- HockA DemmelU SchichaH et al Trace element concentration in human brain Brain 1975 98 49 64 1:STN:280:DyaE2M7ivVOlsg%3D%3D https://doi.org/10.1093/brain/98.1.49 1122375
- JohnsonFN Effects of alkali metal chlorides on activity in rats Nature 1972 238 333 334 1:CAS:528:DyaE38XltlGktLs%3D https://doi.org/10.1038/238333b0 4561839
- HoffmannC SmithDF Lithium and rubidium: effects on the rhythmic swimming movement of jellyfish Cell Mol Life Sci 1979 35 1177 1178 1:CAS:528:DyaE1MXlvVOhur0%3D https://doi.org/10.1007/BF01963271
- RelmanAS The physiological behavior of rubidium and cesium in relation to that of potassium Yale J Biol Med 1956 29 248 262 2603856 1:CAS:528:DyaG2sXptlajtA%3D%3D 13409924
- AlversonDL LonghurstAR GullandJA et al How much food from the sea? Science 1970 168 503 505 1:STN:280:DC%2BC3cvlvVKqsw%3D%3D https://doi.org/10.1126/science.168.3930.503 17838131
- ButlerA Acquisition and utilization of transition metal ions by marine organisms Science 1998 281 207 210 1:CAS:528:DyaK1cXksFKjtbk%3D https://doi.org/10.1126/science.281.5374.207 9660742
- SchutzDF TurekianKK The investigation of the geographical and vertical distribution of several trace elements in sea water using neutron activation analysis Geochim Cosmochim Acta 1965 29 259 313 1:CAS:528:DyaF2MXovFCgtw%3D%3D https://doi.org/10.1016/0016-7037(65)90022-0
- JamesRH PalmerMR Marine geochemical cycles of the alkali elements and boron: The role of sediments Geochim Cosmochim Acta 2000 64 3111 3122 1:CAS:528:DC%2BD3cXnt1yisr8%3D https://doi.org/10.1016/S0016-7037(00)00418-X
- Von DammKL EdmondJM GrantB et al Chemistry of submarine hydrothermal solutions at 21n, east pacific rise Geochim Cosmochim Acta 1985 49 2197 2220 1:CAS:528:DyaL28Xos1ei https://doi.org/10.1016/0016-7037(85)90222-4
- MartinJH FitzwaterSE GordonRM Iron deficiency limits phytoplankton growth in antarctic waters Global Biogeochemical Cycles 1990 4 5 12 1:CAS:528:DyaK3MXitFersLk%3D https://doi.org/10.1029/GB004i001p00005
- Gomez-CabreraMC DomenechE RomagnoliM et al Oral administration of vitamin c decreases muscle mitochondrial biogenesis and hampers training-induced adaptations in endurance performance Am J Clin Nutri 2008 87 142 149 1:CAS:528:DC%2BD1cXmt1amsw%3D%3D
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.