Abstract
The aim of this study was to evaluate the effect of oat bran supplementation on time to exhaustion, glycogen stores and cytokines in rats submitted to training. The animals were divided into 3 groups: sedentary control group (C), an exercise group that received a control chow (EX) and an exercise group that received a chow supplemented with oat bran (EX-O). Exercised groups were submitted to an eight weeks swimming training protocol. In the last training session, the animals performed exercise to exhaustion, (e.g. incapable to continue the exercise). After the euthanasia of the animals, blood, muscle and hepatic tissue were collected. Plasma cytokines and corticosterone were evaluated. Glycogen concentrations was measured in the soleus and gastrocnemius muscles, and liver. Glycogen synthetase-α gene expression was evaluated in the soleus muscle. Statistical analysis was performed using a factorial ANOVA. Time to exhaustion of the EX-O group was 20% higher (515 ± 3 minutes) when compared with EX group (425 ± 3 minutes) (p = 0.034). For hepatic glycogen, the EX-O group had a 67% higher concentrations when compared with EX (p = 0.022). In the soleus muscle, EX-O group presented a 59.4% higher glycogen concentrations when compared with EX group (p = 0.021). TNF-α was decreased, IL-6, IL-10 and corticosterone increased after exercise, and EX-O presented lower levels of IL-6, IL-10 and corticosterone levels in comparison with EX group. It was concluded that the chow rich in oat bran increase muscle and hepatic glycogen concentrations. The higher glycogen storage may improve endurance performance during training and competitions, and a lower post-exercise inflammatory response can accelerate recovery.
Electronic supplementary material
The online version of this article (doi:10.1186/1550-2783-7-32) contains supplementary material, which is available to authorized users.
Background
The importance of dietary carbohydrates (CHO) in sporting performance was shown in the classical gaseous exchange experiments and biopsy studies, in which increasing exercise intensity utilises a greater proportion of CHO [Citation1, Citation2]. These data provided a major breakthrough for the science of sports nutrition, as it enabled the exact amount of CHO for athletes to be quantified.
The recommendations concerning carbohydrates (CHO) for athletes are around 6 g-10 g/Kg/day [Citation3–Citation5] and these quantities vary in accordance with the quantity of body mass, gender, volume and intensity of the training. According to Tarnopolsky [Citation3] elite athletes train around 8 to 40 hours per week, exponentially increasing their nutritional needs. The International Olympic Committee (IOC) recommends that: "following a diet rich in carbohydrates days before a competition can help to increase sporting performance, particularly when the exercise is kept up for longer than 60 minutes" [Citation6].
Exhaustive endurance exercise can induce immune disturbances and consequently increase susceptibility to upper respiratory tract infections [Citation7]. Several mechanisms have been proposed in an attempt to explain the susceptibility of athletes to respiratory infections. Cortisol contributes only minimally to the exercise induced rise in liver glucose output [Citation8], while it plays a role in immune disturbances [Citation9, Citation10]. Several components of the innate immune system are compromised during single or repeated sessions of exercise stress.
Physical exercise can affect the levels of systemic cytokines, such as TNF-α [Citation11–Citation13], interleukin 1 beta (IL-1β) [Citation12], IL-6 [Citation12–Citation16], interferon and others [Citation11]. Recently, it has been suggested that the disruptions in the balance between pro- and antiinflammatory cytokines may lead to a loss of inflammatory control, with possible implications for overall immune system function [Citation17, Citation18]. The effect of ingesting carbohydrates during long duration exercises, with the purpose of attenuating immune suppression is well established [Citation6, Citation12–Citation14].
Cereals oat bran has a high nutritional quality, an naturally source of CHO [Citation19], rich in proteins, unsaturated fatty acids, vitamins, and complex starches that comprise the part with the largest quantity of soluble fiber. Another important nutrient in oat bran is β-Glucan, and has well-documented stimulation effects on the immune system. Also may help enhance immune resistance to various viral, bacterial, protozoan, and fungal diseases [Citation20]. Animal studies show that oat β-glucan can offset exercise-induced immune suppression and decrease susceptibility to infection during heavy training [Citation21]. Therefore, the aim of this study was to evaluate the effect of oat bran supplementation on time to exhaustion, glycogen stores and cytokines profile in rats submitted to training.
Materials and methods
Experimental groups
All experiments were conducted according to the policy of the American College of Sports Medicine on Research with Experimental Animals. Two-month-old male Wistar rats (Rattus novergicus var. albinus, Rodentia, Mammalia) with a mean ± SEM weight of 200 ± 5 g were used. The animals had free access to water and were fed a commercial chow for rodents (NUVILAB, Purina®) ad libitum. The animals were kept in collective cages (3 rats per cage) at a constant temperature of 23 ± 2°C, and a cycle of 12 hours light/12 hours darkness, with light from 06:00 h to 18:00 h (in pathogen-free housing). Before the experimental period began, the animals underwent 48 hours of adaptation to the research laboratory conditions. The 27 animals were divided into 3 groups (n = 9 each group): sedentary control group that underwent no physical training (C), an exhaustion group that received a control chow (EX) and an exhaustion group that received a chow supplemented with 30% of soluble oat bran fibers (EX-O). All experiments conducted on animals were previously approved by the Ethics Committee on Animal Testing, Federal University of San Carlos.
Chow Preparation
For eight weeks, the animals received chow prepared weekly, stored and analyzed. Only carbohydrate, protein, lipid and fibre content in chow were analyzed. Every care was taken to ensure that these diets remained homogeneous during the entire experimental period. The chow was prepared from a commercial chow (NUVILAB, Purina®) which, after milling, had its fibre content adjusted by adding 30% of oat bran (Oat bran Quaker®), or 300 g/Kg of standard commercial chow. The chow was characterized according to the procedures of Cavaglieri [Citation22]. Table demonstrates the chow compositions.
Table 1 Nutritional Composition in grams (g) of the chows used.
Exercise Protocol
The animals were submitted to a 5-day period of adaptation to the liquid environment (5 minutes on the first day, 15 minutes on the second, 30 minutes on the third, 45 minutes on the fourth and 60 minutes on the fifth), in accordance with Sampaio-Barros [Citation23]. Importantly, the control groups were submitted in contact with water, but did not perform the exercise. This was done to equalize the stress compared to the exercised group. A tank was used to perform the swimming sessions, were made of plastic and did not have places where animals could cling to. This was necessary to achieve constant exercise. The water temperature was monitored at approximately 28 ± 2°C. After adaptation, the training consisted of 60 minutes of daily swimming, five days per week, for eight weeks, performed in the afternoon between 14:00-17:00. The moderate intensity they used a load of 5% of their body weight strapped to their backs, which corresponds to intensity below the point of inflection of the lactate threshold curve. At the end of eight weeks training, the animals were submitted to the exhaustion test, characterized by being incapable of keeping themselves on the surface of the water [Citation24, Citation25].
Animal sacrifice and sample collection
Immediately after the exhaustion test, the animals were sacrificed by decapitation. During exsanguination, the mixed arteriovenous blood was collected in heparinized tube and chilled on ice. Blood was then spun at 500g for 15 min to obtain plasma for cytokine and corticosterone analyses. In the following order, the liver, soleus and white and red gastrocnemius muscle were collected and stored at -70°C until the time of measurement of hepatic and muscle glycogen. The white and red portion of the gastrocnemius was divided throughout the major colour of muscle fibres.
Determination of muscle and hepatic glycogen concentrations
The muscle samples were digested in 30% KOH at 100° C and glycogen was precipitated by passage through ethanol. Between each precipitation the sample was centrifuged at 3000 rpm for 15 minutes. The precipitated glycogen was submitted to acid hydrolysis in the presence of phenol. The values were expressed in mg/100 mg of wet weight, using the Siu method [Citation26].
Determination of serum cytokines
After the period of supplementation and training, measurements of IL-6, TNF-α and IL-10 in plasma were made by ELISA using the R & D Systems Quantikine High Sensitivity kit (R&D Systems, Minneapolis, MN, USA) for each cytokine. The intra-assay coefficient of variance (CV) was 4.1 - 10%, the inter-assay CV was 6.6 - 8%, and the sensitivity was 0.0083 pg/ml [Citation13]. The duplicate plasma aliquots for all cytokines analysis were used.
Corticosterone determination
Plasma corticosterone was determined by ELISA, using the Stressgen kit (Corticosterone ELISA KIT Stressgen@), Michigan, USA). The sensitivity range of the assay was 32-20.000 ng/ml. The duplicate plasma aliquots for hormone analysis were used.
Determination of glycogen synthetase-alpha (GS-α) mRNA expression in the soleus muscle
Total RNA extraction
Total RNA was obtained from 100 mg of soleus muscle. The tissue were stored at -70°C until the time of measurement. Cells were lysed using 1 mL of Trizol reagent (Life Technologies, Rockville, MD, USA). After incubation of 5 min at room temperature, 200 μL chloroform was added to the tubes and centrifuged at 12,000 × g. The aqueous phase was transferred to another tube and the RNA was pelleted by centrifugation (12,000 × g) with cold ethanol and air-dried. After this, RNA pellets were diluted in RNase-free water and treated with DNase I. RNAs were stored at -70°C until the time of measurement. RNA was quantified by measuring absorbance at 260 nm. The purity of the RNAs was assessed by the 260/280 nm ratios and on a 1% agarose gel stained with ethidium bromide at 5 μg per mL [Citation27].
RT-PCR
RT-PCR was performed using parameters described by Innis and Gelfand [Citation28]. The number of cycles used was selected to allow quantitative comparison of the samples in a linear manner. For semi-quantitative PCR analysis, the housekeeping β-actin gene was used as reference. The primer sequences and their respective PCR fragment lengths are: GSK3-α sense: AATCTCGGACACCACCTGAGG - 3'; anti-sense: 5'GGAGGGATGAGAATGGCTTG - 3'. Control: β-actina sense: 5'-ATGAAGATCCTGACCGA GCGTG-3'; anti-sense: 5'- TTGCTGATCCACATCTGCTGG-3'. Published guidelines were followed to guard against bacterial and nucleic acid contamination [Citation29].
Analysis of the PCR products
The PCR amplification products were analyzed in 1.5% gels containing 0.5 μg per mL of ethidium bromide and were electrophoresed for 1 h at 100 V. The gels were photographed using a DC120 Zoom Digital Camera System from Kodak (Life Technologies, Inc., Rockville, MD, USA). The images were processed and analyzed in the software Kodak Digital Science 1D Image Analysis (Life Technologies). PCR band intensities were expressed as Optic Density (OD) normalized for β-actin expression. Data are presented as a ratio compared with the respective controls, which received an arbitrary value of 1 in each experiment.
Statistical analysis
Data are presented as mean ± SEM (standard error of the mean). The normality of distribution of all parameters was checked with the Kolmogorov-Smirnov test and by the homocedasticity test (Bartlett criterion). All variables presented normal distribution and homocedasticity, thus the two-way ANOVA test was used, (taking into consideration the variables exercise × oat bran enriched diet) and when the difference presented was significant, Tukey's post hoc test was used. A significance level of p ≤0.05 was used for all comparisons. The software package used was SPSS for Windows version 10.0.
Results
Time to Exhaustion
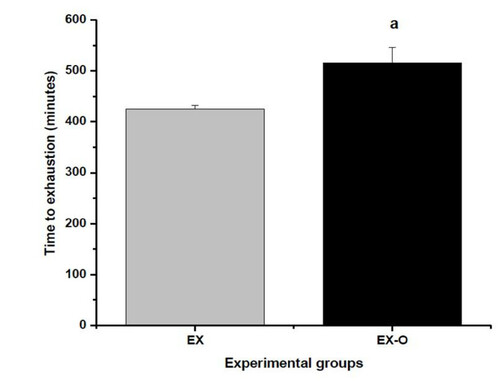
The time to exhaustion of the EX-O group was 515 ± 30 minutes and 425 ± 30 for the EX group (p = 0.034). This represented a 20% higher exhaustion time for the EX-O group when compared with the EX group. Figure
Figure 1 Time to exhaution on experimental groups. a = statistical difference to exhaution group (EX)
Corticosterone and Cytokine Concentrations
Corticosterone levels were significantly elevated after exercise to exhaustion compared with the control group. The EX group presented significantly higher corticosterone levels compared with the EX-O group, (p = 0.039) (figure ). Similarly, after exercise IL-6 was increased in EX and EX-O compared with the control. The EX-O group showed lower levels of IL-6 compared with the EX group, (p = 0.001) (Table ). The serum levels of TNF-α were significantly decreased after exercise in the EX and EX-O groups compared with the control group. However, no statistically significant differences were observed between EX and EX-O for TNF-α serum levels (Table ). IL-10 serum levels were increased after exercise compared with the control group, and EX presented significantly higher levels of IL-10 as compared with EX-O (p = 0.032) (Table ).
Figure 2 Corticosterone levels in experimental groups. a = statistical difference to control group b = statistical difference to EX group
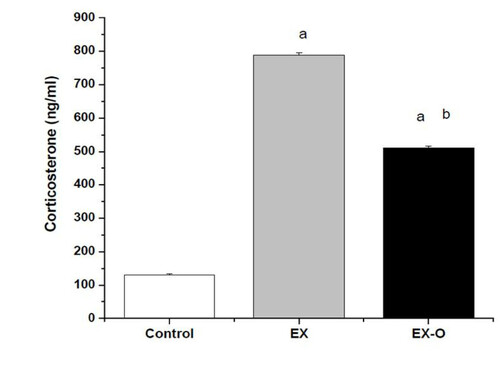
Table 2 Plasma cytokine concentration in experimental groups.
Glycogen Concentrations and expression of Glycogen synthetase mRNA
With regard to hepatic glycogen, the exhaustion test diminished the hepatic glycogen of the EX-O group by 61% and that of the EX group by 87% in comparison with the control group. In the comparison between the exercise groups, EX-O presented a 67% higher hepatic glycogen concentrations when compared with EX (p = 0.022), as shown in Table .
Table 3 Hepatic and muscle glycogen concentration (mg/100 mg)
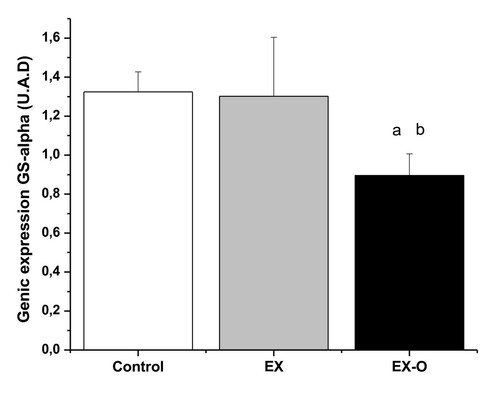
There was a decrease of 47% in soleus muscle glycogen concentrations for the EXO group (p = 0.043), and of 78.5% for the EX group (p = 0.036) when compared with the control group. When comparing the exercise groups, EX-O presented a 59.4% higher soleus glycogen concentrations than the EX group (p = 0.021, see Table ). Gene expression of GS-alpha (U.A.D) in the C group was 1.32 ± 0.1, EX group 1.30 ± 0.3 and EXO group 0.89 ± 0.1 (Figure ). Furthermore, the EX-O group presented lower levels of glycogen synthetase-α enzyme in the soleus muscle when compared with the EX group (p = 0.013).
Figure 3 Glucogen synthetase gene expression. a = statistical difference with control group b = statistical difference with EX group
The quantity of glycogen in the white gastrocnemius muscle decreased by 77% in the EX-O (p = 0.011), and 80% in the EX group (p = 0.037) when compared with the control. There were no significant differences between EX-O and EX in the glycogen concentrations of the white gastrocnemius muscle (Table ).
The exhaustion test diminished the muscle glycogen concentrations of the red gastrocnemius by 69.8% in the EX-O group, and by 73.5% in the EX group (p < 0.05), when compared with the control group. In the comparison between the exercise groups, no significant differences were observed (Table ).
Discussion
The aim of this study was to evaluate the effect of oat bran supplementation on time to exhaustion, glycogen stores and cytokines profile in rats submitted to training. The animals did not receive any type of carbohydrate during the time they were performing the exercise, only ad libitum food during the eight weeks of training. In the present investigation, the exercise protocol used was one hour of daily swimming, 5 days per week during two months At the end of the eight weeks, were performed the test exhaust. For the impact in performance, the carbohydrate content should be equal, there by the experimental chows had the same quantity of carbohydrates, being 45.5 g/100 g for the control and 45.6 g/100 g in the experimental chow.
Similarly, in the chows of the present study, one can note a lower quantity of total fibres in the experimental chow (18.9 g) and a higher quantity in the soluble portion (4.5 g). Although the total fibre content was higher (21.9 g) in the control diet, the quantity in the soluble part was lower (3.9 g).The difference in available carbohydrate (avCHO = total carbohydrate minus fiber) is the better explanation: control chow has 45.5 cho-21.9 fiber = 23.6 g avCHO while the oat bran diet contains 45.6 cho-18.9 fiber = 26.7 g avCHO. It is a 13% increase in the oat bran chow.
Changes in the intestinal microflora that occur with the consumption of prebiotic fibres may potentially mediate immune changes via: the direct contact of lactic acid bacteria or bacterial products (cell wall or cytoplasmic components) with immune cells in the intestine; the production of short-chain fatty acids from fibre fermentation; or by changes in mucin production. The link between oat bran and immune system its regard with the content of β-glucan, especially water-soluble β-glucan. This soluble fiber can enhance the activities of both the innate and specific immune system components via direct activation of specific receptors on macrophage, neutrophils, and NK cells [Citation30, Citation31] or indirectly after activation of pinocytic M-cells located in the Peyer's patches of the small intestine [Citation32, Citation33]. There is increasing evidence that fermentable dietary fibres and the newly described prebiotics can modulate various properties of the immune system, including those of the gut-associated lymphoid tissues (GALT).
In published data on the immune system of the same experimental group, Donatto [Citation34] demonstrated that the EX-O group presented better phagocytic capacity of peritoneal macrophages, increased amount of lymphocytes from lymph nodes and shows less leukocytosis after exhausting exercise. We found no side effects in this study, including no increase in the plasma concentration of pro inflammatory cytokine. β-glucan found in oat bran could not exaggerate the inflammatory response to severe exercise.
Glycogen metabolism is largely controlled by the actions of glycogen synthase and glycogen phosphorylase enzymes [Citation35]. The gene expression of Glycogen synthase increased after both resistance and aerobic training, but not when aerobic exercise was combined with a high CHO diet in comparison with diet without exercise [Citation36]. In the present study, we found a lower expression of the glycogen synthetase enzyme in the soleus muscle in the EXO group. Probably, the higher glycogen levels in the soleus muscle had an important relationship with the impaired glycogen synthetase expression. It may reflect a lower need for re-synthesis [Citation37] since this group presented higher glycogen concentrations in the soleus when compared with exhaustion of the non-oat bran enriched diet group (EX).
The oat bran is a nutritional search of dietary fiber, especially soluble fiber and this nutriente may retard the absorption of nutrients by the intestinal villosities [Citation38]. In this case, the glucose absorption metabolism had a modulation to lower and constant delivery to blood circulation and this could be responsible for a more efficient replacement of muscular glycogen during a longer recovery period [Citation37, Citation39, Citation40]. There was a correlation between the low levels of glycogen and higer corticosterone and IL-6. During prolonged and exhausting physical exercises (duration in excess of 90 minutes), the IL-6 has a close relationship with the amount of muscle glycogen and regulation of the homeostasis of blood glucose during long duration exercises. Muscular glycogen and blood glucose are the major sources of substrates for oxidative metabolism, and the immune depletion and fatigue coincides with their depletion, due to the low availability to the skeletal muscle and the central nervous system [Citation41–Citation45].
In the EX group glycogen levels were low while IL-6 and corticosterone were high. In contrast, the inverse was observed in the EX-O group which had higher levels of muscle glycogen and lower levels of corticosterone and IL-6. These results were shown in EX group, since the animals swam an average of 11 hours, ending in a worst metabolic condition On the other hand, EX-O swam an average of 2 hours longer, totalling 13 hours of physical exercise with lower levels of IL-6 and corticosterone, consequently at the end of exercise protocol shows an better condition.
Plasma concentration shows the total secreted of some products like corticosterone and cytokines by all tissues, but does not know the source of secretion. Unfortunately, some of the shortcomings of this study were not to analyze the cytokines levels in different tissues. One of the hypotheses regarding the mechanism of central fatigue is that IL-6 can exert direct influence on hypothalamus-pituitary-adrenal axis, thereby increasing ACTH-cortisol release [Citation15, Citation46]. Moreover, the different kits used to measure IL-6 plasma levels difficult the comparison between studies.
The exercise protocol used in the present study modulated the serum levels of TNF-α, as a result of the lower levels of TNF-α in the trained groups when compared with the control group. In 1999, Ostrowski and colleagues [Citation47] presented the plasma cytokines profile after a marathon race (mean duration 3: 26 (h: mi.), with increased levels of TNF-a, IL-6 and IL-10. Their study revealed a proinflammatory and anti-inflammatory profile after a marathon race. Pedersen [Citation16] suggested that regular exercise modulates some pro-and anti-inflammatory cytokines, induces suppression of TNF-alpha and thereby offers protection against exacerbated inflammation.
Unfortunately, the levels of cytokines in the adipose tissue and muscle were not measured, so that the source of cytokine production cannot be determined. This is an important issue because there is a different production of cytokines in muscle and adipose tissue, and exercise has an influence in this process. Rosa Neto et al. [Citation48] showed an anti-inflammatory effect of strenuous exercise on muscle and a pro-inflammatory effect on adipose tissue.
In this sense, Pedersen (16) revealed an anti-inflammatory effect of acute physical exercise, characterized by an increased circulating level of IL-10, IL-1 receptor antagonist (IL-1ra) and soluble receptor of TNF (TNFRs). Lira et al. [Citation49] showed an anti-inflammatory profile on adipose tissue in rats submitted to aerobic training (decreased TNF-alpha and increased IL-10 levels). In the present study, the combination of exercise with oat bran induced a decrease on TNF-alpha levels associated with an increase in IL-10 serum levels (anti-inflammatory cytokine).
These results show that oat bran, how another search of carbohydrate can directly influence the metabolic stress induced by exhaustive long duration exercise, saving the energy reserves and promoting better performance during exercise, thus corroborating findings in the literature [Citation7, Citation15, Citation42, Citation44]. If our data can be clinically translated, they may lead to an important new nutritional strategy to boost the immune system and decrease the risk of infection that can be a problem in athletes and military personnel who are often exposed to combinations of severe physical, psychological, and environmental stressors. In practical terms, athletes who practice long duration exercises may maintain the stocks of glycogen at more favourable concentrations to perform daily training sessions, by means of ingesting carbohydrate, vitamins, minerals, and β-glucan in the form of oat bran.
Conclusions
In summary, it could be concluded that soluble fibres (i.e. chow rich in oat bran) increased muscular and hepatic glycogen concentrations, and this resulted in a longer time to exhaustion with an associated reduction in pro-inflammatory cytokines. In practical terms, these results demonstrate the importance, not only of the quantity of carbohydrates, but also the balance of dietary fibre content. Further studies conducted in athletes and animal models, using oat bran supplementation are necessary, with the aim of assessing improved performance, in view of the possible positive effects found in the present research.
Authors’ original file for figure 1
Download PDF (40.8 KB)Authors’ original file for figure 2
Download PDF (46.5 KB)Authors’ original file for figure 3
Download PDF (53.4 KB)Authors’ original file for figure 4
Download JPEG Image (42.7 KB){kind=link}
Authors’ original file for figure 5
Download JPEG Image (42.7 KB){kind=link}
Authors’ original file for figure 6
Download JPEG Image (42.7 KB){kind=link}
Acknowledgements
The authors thank CAPES for the financial support
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
CC: dissertation guidance, interpretation of the data and preparation of experimental chow; JP: randomization of the protocol training of animals, literature review and ELISA assays assistance; FA and DR: animal training assistance; RV and AP: molecular biology assays. All authors read and approved the final manuscript.
Related Research Data
References
- ChristensenEHDer Stoffwechsel und die Respiratorischen Funktionen bei schwerer ko¨rperlicher ArbeitScand Arch Physiol193281160https://doi.org/10.1111/j.1748-1716.1939.tb01320.x
- BergstromJHultmanEA study of glycogen metabolism during exercise in manScand J Clin Invest1967192181:CAS:528:DyaF2sXltFCqu70%3Dhttps://doi.org/10.3109/003655167090906296048626
- TarnopolskyMAGibalaMJeukendrupAPhillipsSMNutritional needs of elite endurance athletes. Part1: Carbohydrate and fluid requirementsEuropean Journal of Sports Sciences200551314https://doi.org/10.1080/17461390500076741
- American College of Sports Medicine and Dietitians Canada Joint Position Statement. Nutrition and Athletic PerformanceMedicine Science and Sports Exercise2000321221302145https://doi.org/10.1097/00005768-200012000-00025
- BurkeLMKiensBIvyJLCarbohydrate and fat for training and recoveryJournal of Sports Sciences2004221530https://doi.org/10.1080/026404103100014052714971430
- JeukendrupACarbohydrate intake during exercise and performanceNutrition2004206696771:CAS:528:DC%2BD2cXltVGitbs%3Dhttps://doi.org/10.1016/j.nut.2004.04.01715212750
- NiemanDCDavisJMHensonDWalberg-RankinAJShuteMCLDumkeACUtterDMVinciJACarsonABrownWJLeeSRMcanultyAMcanultyLSCarbohydrate ingestion influences skeletal muscle cytokine mRNA and plasma cytokine levels after a 3-h runJournal of Applied Physiology200394191719251:CAS:528:DC%2BD3sXjslSmsbc%3Dhttps://doi.org/10.1152/japplphysiol.01130.200212533503
- KjaerMHepatic glucose production during exerciseAdvances in Experimental Medicine and Biology1998441117271:CAS:528:DyaK1cXmvFKmt7w%3Dhttps://doi.org/10.1007/978-1-4899-1928-1_119781319
- MigniniFTrainiETomassoniDVitaliMStreccioniVLeucocyte subset redistribution in a human model of physical stressClinical Experimental Hypertension200830872031https://doi.org/10.1080/0742052080257233319021023
- ZarkovicMIgnjatovicSDajakMCiricJBeleslinBSavicSStojkovicMBulatPTrbojevicBCortisol response to ACTH stimulation correlates with interleukin 6 concentration in healthy humansEuropean Journal of Endocrinology20081595649521:CAS:528:DC%2BD1cXhsVClsLvFhttps://doi.org/10.1530/EJE-08-054418753312
- RivierAet alRelease of cytokines by blood monocytes during strenuous exerciseInternational Journal of Sports Medicine1994151921981:STN:280:DyaK2czktF2ltg%3D%3Dhttps://doi.org/10.1055/s-2007-10210468063468
- MoldoveanuAIet alExercise elevates plasma levels but not gene expression of IL1 beta, IL-6, and TNF-alpha in blood mononuclear cellsJournal Applied Physiology200089414995041:CAS:528:DC%2BD3cXotlylurY%3D
- PrestesJDe FerreiraCKDiasRFrolliniABDonattoFFCury-BoaventuraMFGuereschiMGPithon-CuriTCVerlengiaRPalanchACCuriRCavaglieriCRLymphocyte and Cytokines after Short Periods of ExerciseInternational Journal of Sports Medicine200829101010141:STN:280:DC%2BD1cjgs12rsw%3D%3Dhttps://doi.org/10.1055/s-2008-103873718600609
- OstrowskiKet alPhysical activity and plasma interleukin-6 in humans - effect of intensity of exerciseEuropean Journal of Applied Physiology2000835125151:CAS:528:DC%2BD3cXovVCmtrs%3Dhttps://doi.org/10.1007/s00421000031211192058
- PedersenBKSteenbergAFischerCKellerCKellerPPlomgaardPWolsk-PetersenEFebbraioMThe metabolic role of IL-6 produced during exercise: is IL-6 an exercise factor?Proceeds of Nutrition Society2004632632671:CAS:528:DC%2BD2cXms12msLs%3Dhttps://doi.org/10.1079/PNS2004338
- PedersenBKThe anti-inflammatory effect of exercise: its role in diabetes and cardiovascular disease controlEssays Biochemistry200642105171:CAS:528:DC%2BD2sXhvFSmurY%3Dhttps://doi.org/10.1042/bse0420105
- CoxAJPyneDBSaundersPUCallisterRGleesonMCytokine responses to treadmill running in healthy and illness-prone athletesMedicine and Science in Sports and Exercise20073911191819261:CAS:528:DC%2BD2sXht1KrurvIhttps://doi.org/10.1249/mss.0b013e318149f2aa17986898
- PuglisiMAVaishnavUShresthaSWTorres-GonzalexMWoodRJVolekJSFernandezMLRaisins and additional walking have distinct effects on plasma lipids and inflammatory cytokinesLipids in Health and Disease20087114https://doi.org/10.1186/1476-511X-7-14
- CarusoLMenezesEWGlycemic index of foodsJournal of Brazilian Society of Food and Nutrition2000194964
- NiemanDCPedersenBKNutrition and Exercise Immunology Boca Raton2000FLCRC Press
- DavisJMMurphyEABrownACarmichaelMGhaffarAMayerEPEffects of moderate exercise and oat β-glucan on innate immune function and susceptibility to respiratory infectionAmerican Journal of Physiology Integrative and Comparative Physiology2004286366372https://doi.org/10.1152/ajpregu.00304.2003
- CavaglieriCRMartinsEFColleoneVVRodriguesCVecchiaMGCuriRFibre-rich diets alter rat intestinal leukocytes metabolismJournal of Nutrition and Biochemistry2000115555611:CAS:528:DC%2BD3MXlslKrsQ%3D%3Dhttps://doi.org/10.1016/S0955-2863(00)00118-2
- Sampaio-BarrosMMEffect of swimming session duration and repetition on metabolic markers in ratsStress200362127321:CAS:528:DC%2BD3sXktVGltLk%3Dhttps://doi.org/10.1080/102538903100011016912775332
- VoltarelliFAGobattoCADe MelloMADetermination of anaerobic threshold in rats using the lactate minimum testBraz J Med Biol Res200235111389941:CAS:528:DC%2BD3sXjtVKhsw%3D%3Dhttps://doi.org/10.1590/S0100-879X200200110001812426640
- DawsonCAHarvathSMSwimming in small laboratory animalsMedicine and Science in Sports1970251781:CAS:528:DyaE3MXhtlOnsLc%3D4939286
- SiuLOet alDetermination of glycogen in small tissue samplesJournal of Applied Physiology1970282234236
- SambrookJRussellDWMolecular cloning: A laboratory manual20013 N.YCold Spring Harbor Laboratory Press, Cold Spring Harbor A8.21.
- InnisMAGelfandDHInnisMAGelfandDHSninskyJJWhiteTJOptimization of PCRs. In PCR protocols: a guide to methods and applications19901 San Diego, CAAcademic Press312
- KwokSHiguchRAvoiding false positives with PCRNature19893392372381:STN:280:DyaL1M3jvVGqsw%3D%3Dhttps://doi.org/10.1038/339237a02716852
- CzopJKAustenKFA. B-glucan inhibitable receptor onhuman monocytes: its identity with the phagocytic receptor for particulate activators of the alternative complement pathwayJ Immunol1985134258825931:CAS:528:DyaL2MXhvVemtL4%3D2579146
- VetivickaVThorntonBPRossGDSoluble _-glucan polysaccharide binding to the lectin site of neutrophil or natural killer cell complement receptor type 3 (CD11b/CD18) generates a primed state of the receptor capable of mediating cytotoxicity of iC3b-opsonized target cellsJ Clin Invest1996985061https://doi.org/10.1172/JCI118777
- SakuraiTHasimotoKSuzukiIet alEnhancement of murine alveolar macrophage functions by orally administered B-glucanInt J Immnopharmacol1992148218301:CAS:528:DyaK38XltVertL4%3Dhttps://doi.org/10.1016/0192-0561(92)90080-5
- SuzukiITanakaHKinoshitaAOikawaSOsawaMYadomaeTEffect of orally administered b-glucan on macrophage function in miceInt J Immunopharmacol1990126756841:CAS:528:DyaK3cXlvVCgtbc%3Dhttps://doi.org/10.1016/0192-0561(90)90105-V2272730
- DonattoFPrestesJFerreiraCKDiasRFroliniALeiteGUrtadoCVerlengiaRPalanchAPerezSCavaglieriCEffects of soluble fibers supplementation on immune system cells after exhausting exercise in trained ratsRev Bras Med Esporte200814653337https://doi.org/10.1590/S1517-86922008000600011
- PilegaardHKellerCSteensbergAHelgeJWPedersenBKSaltinBNeuferDInfluence of pre-exercise muscle glycogen concentrations on exercise-induced transcriptional regulation of metabolic genesJournal Physiology2002541261271https://doi.org/10.1113/jphysiol.2002.016832
- HaysNPStarlingRDSullivanDHFluckeyJDCokerRHWilliamsRHEvansWJEffects of an ad libitum, high carbohydrate diet and aerobic exercise training on insulin action and muscle metabolism in older men and womenJournal of Gerontology A Biological Sciences2006613299304https://doi.org/10.1093/gerona/61.3.299
- FebbraioMAKeenanJAngusDJCampbellSEGarnhamAPreexercise carbohydrate ingestion, glucose kinetics, and muscle glycogen use: effect of the glycemic indexJournal of Applied Physiology200089184518511:CAS:528:DC%2BD3cXot1ans78%3D11053335
- SlavinJLDietary fibre and body weightNutrition200521411418https://doi.org/10.1016/j.nut.2004.08.01815797686
- JentjensRJeukendrupADeterminants of post-exercise glycogen synthesis during short-term recoverySports Medicine200333211744https://doi.org/10.2165/00007256-200333020-0000412617691
- WeeSLWilliamsCTsintzasKBoobisLIngestion of a high-glycemic index meal increases muscle glycogen storage at rest but augments its utilization during subsequent exerciseJournal of Applied Physiology2005997077141:CAS:528:DC%2BD2MXpsVaqs7k%3Dhttps://doi.org/10.1152/japplphysiol.01261.200415831796
- GoebelMUMillsPJAcute psychological stress and exercise and changes in peripheral leukocyte adhesion molecule expression and densityPsychmestry Medicine2000626646701:STN:280:DC%2BD3cvmsV2hsQ%3D%3Dhttps://doi.org/10.1097/00006842-200009000-00010
- GleesonMNiemanDPedersenBKExercise, nutrition and immune functionJournal of Sports Sciences200422115125https://doi.org/10.1080/026404103100014059014971437
- KellerCSteensbergAPilegaardHOsadaTSaltinBPedersenBKNeuferPDTranscriptional activation of the IL-6 gene in human contracting skeletal muscle: influence of muscle glycogen concentrationsFASEB Journal200115274827501:CAS:528:DC%2BD3MXptFagtLY%3D11687509
- LancasterGIKhanQDrysdalePTet alEffect of prolonged exercise and carbohydrate ingestion on type 1 and type 2 lymphocyte distribution and intracellular cytokine production in humansJournal of Applied Physiology2005985655711:CAS:528:DC%2BD2MXhsVKmtLg%3Dhttps://doi.org/10.1152/japplphysiol.00754.200415322070
- PedersenBKFebbraioMMuscle-derived inteleukin-6 - A possible link between skeletal muscle, adipose tissue, liver and brainBrain, Behavior, and Immunity2005193713761:CAS:528:DC%2BD2MXntVGjtbc%3Dhttps://doi.org/10.1016/j.bbi.2005.04.00815935612
- SteenbergAFebbraioMvan HallGOsadaTSacchettiMSaltinBPedersenBKInterleukin-6 production in contracting human skeletal muscle is influenced by pre-exercise muscle glycogen contentJournal Physiology2001537633639https://doi.org/10.1111/j.1469-7793.2001.00633.x
- OstrowskiKRohdeTASPSSchjerlingPPedersenBKPro- and anti-inflammatory cytokine balance in strenuous exercise in humansJournal of Physiology199951528729122691321:CAS:528:DyaK1MXhsl2ltb0%3Dhttps://doi.org/10.1111/j.1469-7793.1999.287ad.x9925898
- Rosa NetoJCLiraFSOyamaLZanchiNYamashitaABatistaMJrOllerCSeelaenderMExhaustive exercise causes an anti-inflammatory effect in skeletal muscle and a pro-inflammatory effect in adipose tissue in ratsEur J Appl Physiol20091066977041:CAS:528:DC%2BD1MXotVKgu70%3Dhttps://doi.org/10.1007/s00421-009-1070-119424714
- LiraFRosa NetoJCOyamaLYamashitaABatistaMJrSeelaenderMEndurance training induces depot-specific changes in IL-10/TNF-a ratio in rat adipose tissueCytokine20094580851:CAS:528:DC%2BD1MXhs1ejsrs%3Dhttps://doi.org/10.1016/j.cyto.2008.10.01819097804
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.